

Towards Sustainable Medicinal Resources through Marine Soft Coral Aquaculture: Insights into the Chemical Diversity and the Biological Potential

 ,
,  ,
,  , and
, and 
Abstract
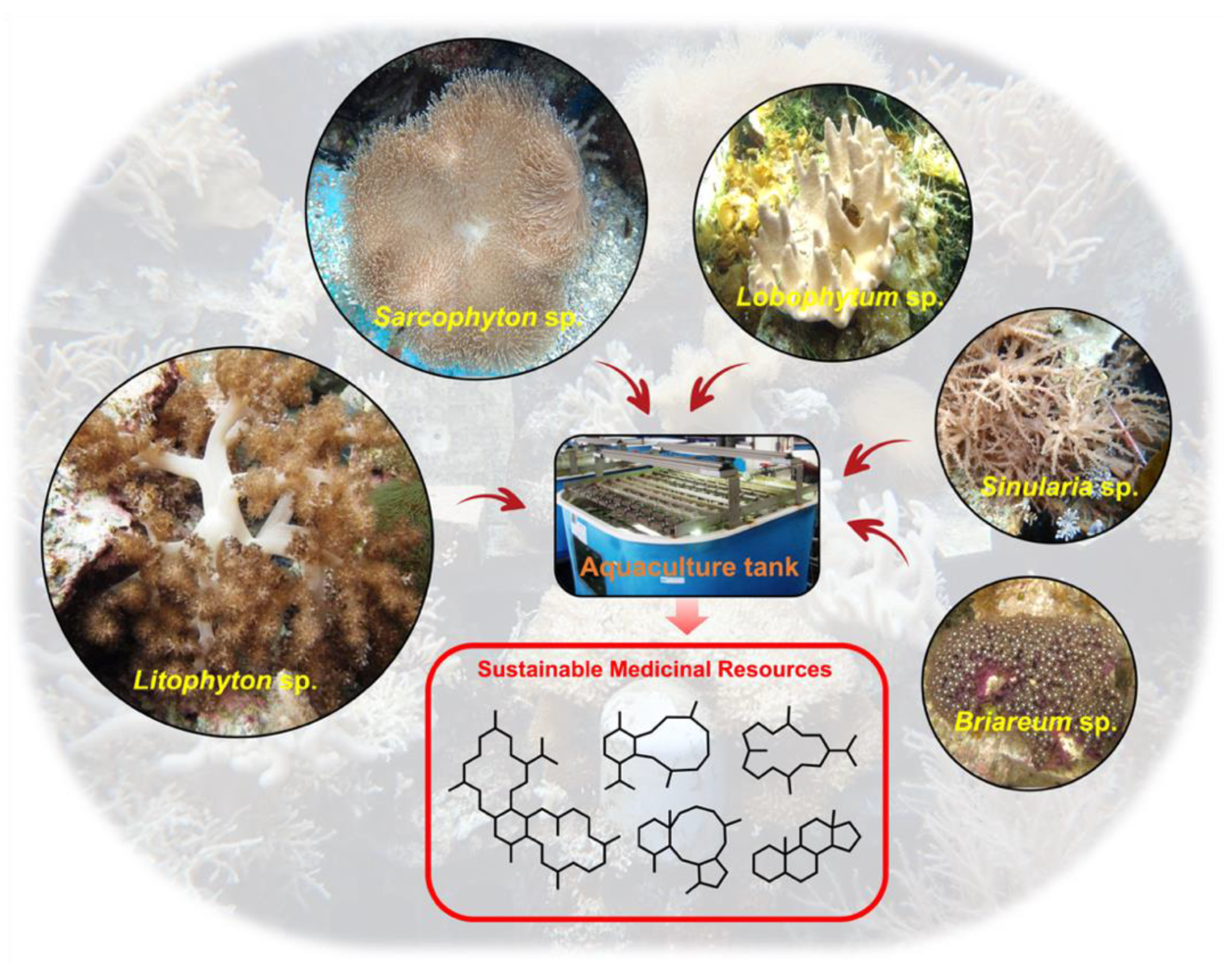
:1. Aquaculture Background
2. Secondary Metabolites Derived from Cultured Soft Corals
2.1. Diterpenes
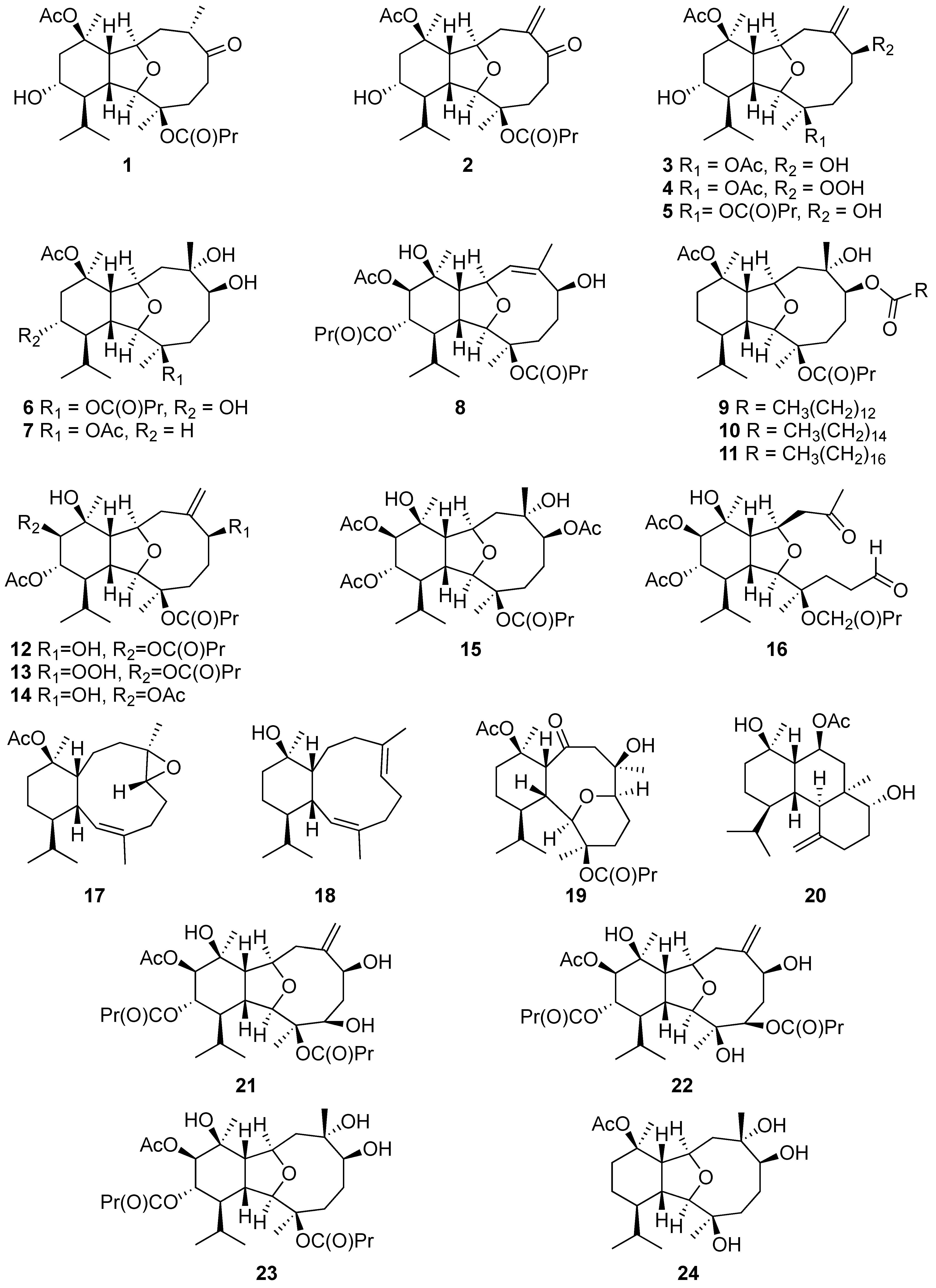
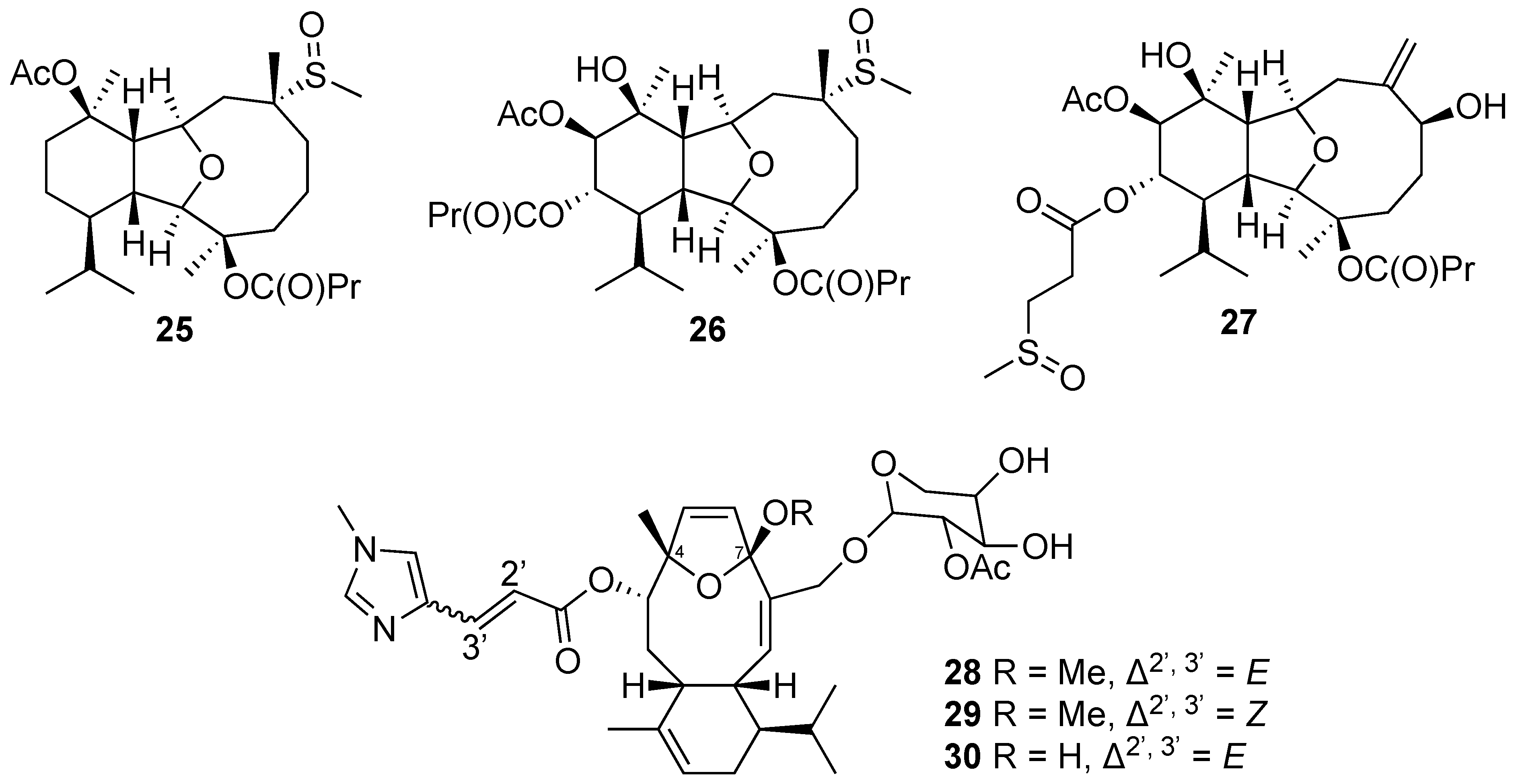
2.1.1. Eunicellin-Based Diterpene
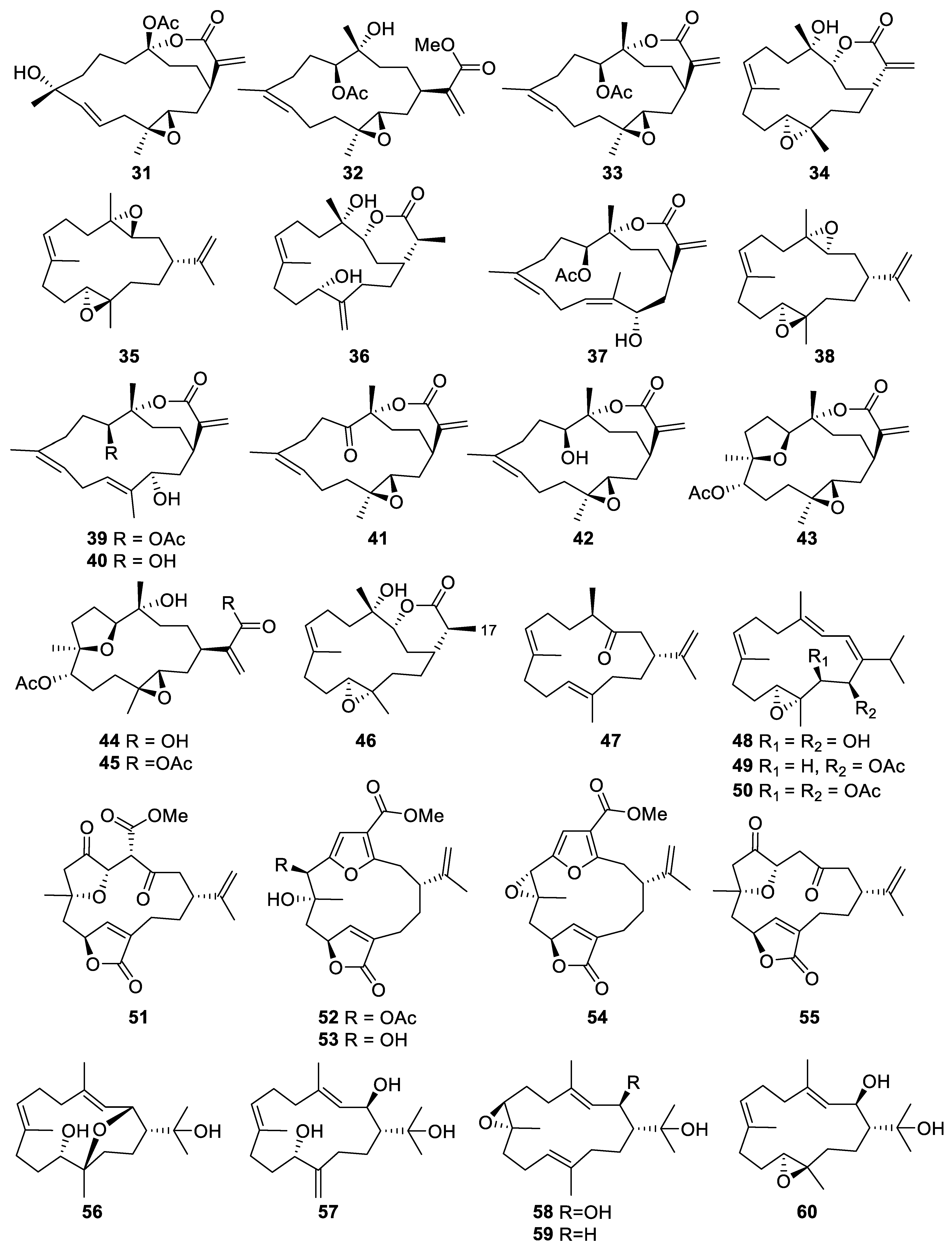
2.1.2. Cembrane-Type Diterpene
2.1.3. Spatane-Type Diterpene
2.1.4. Norcembranoidal Diterpene
2.1.5. Briarane-Type Diterpene
2.1.6. Aquariane-Type Diterpene
2.2. Biscembranoid
2.3. Steroid
2.4. Miscellaneous
3. Active Compounds and Their Diverse Bioactivities
3.1. Anti-Cancer Activity
3.2. Anti-Inflammatory Activity
3.3. Others
4. Conclusion and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Martins, A.; Vieira, H.; Gaspar, H.; Santos, S. Marketed marine natural products in the pharmaceutical and cosmeceutical industries: Tips for success. Mar. Drugs 2014, 12, 1066–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, I.G.; Miguel, M.G.; Mnif, W. A brief review on new naturally occurring cembranoid diterpene derivatives from the soft corals of the genera Sarcophyton, Sinularia, and Lobophytum since 2016. Molecules 2019, 24, 781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, M.E. Structural revisions of natural products by total synthesis. Nat. Prod. Rep. 2009, 26, 1105–1124. [Google Scholar] [CrossRef] [PubMed]
- Delbeek, J. Coral farming: Past, present and future trends. Aquar. Sci. Conserv. 2001, 3, 171–181. [Google Scholar] [CrossRef]
- Bostrom-Einarsson, L.; Babcock, R.C.; Bayraktarov, E.; Ceccarelli, D.; Cook, N.; Ferse, S.C.A.; Hancock, B.; Harrison, P.; Hein, M.; Shaver, E.; et al. Coral restoration-A systematic review of current methods, successes, failures and future directions. PLoS ONE 2020, 15, e0226631. [Google Scholar] [CrossRef]
- Leal, M.C.; Calado, R.; Sheridan, C.; Alimonti, A.; Osinga, R. Coral aquaculture to support drug discovery. Trends Biotechnol. 2013, 31, 555–561. [Google Scholar] [CrossRef]
- Peng, B.R.; Lu, M.C.; El-Shazly, M.; Wu, S.L.; Lai, K.H.; Su, J.H. Aquaculture soft coral Lobophytum crassum as a producer of anti-proliferative cembranoids. Mar. Drugs 2018, 16, 15. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.Y.; Huang, C.Y.; Chao, C.H.; Lin, C.C.; Dai, C.F.; Su, J.H.; Sung, P.J.; Wu, S.H.; Sheu, J.H. New biscembranoids sardigitolides A-D and known cembranoid-related compounds from Sarcophyton digitatum: Isolation, structure elucidation, and bioactivities. Mar. Drugs 2020, 18, 452. [Google Scholar] [CrossRef]
- Huang, T.Y.; Huang, C.Y.; Chen, S.R.; Weng, J.R.; Tu, T.H.; Cheng, Y.B.; Wu, S.H.; Sheu, J.H. New hydroquinone monoterpenoid and cembranoid-related metabolites from the soft coral Sarcophyton tenuispiculatum. Mar. Drugs 2020, 19, 8. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.Y.; Phan, C.S.; Ishii, T.; Kamada, T.; Hamada, T.; Vairappan, C.S. Terpenoids from marine soft coral of the genus Xenia in 1977 to 2019. Molecules 2020, 25, 5386. [Google Scholar] [CrossRef] [PubMed]
- Elshamy, A.I.; Nassar, M.I.; Mohamed, T.A.; Hegazy, M.E. Chemical and biological profile of Cespitularia species: A mini review. J. Adv. Res. 2016, 7, 209–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Lateff, A.; Alarif, W.M.; Alburae, N.A.; Algandaby, M.M. Alcyonium octocorals: Potential source of diverse bioactive terpenoids. Molecules 2019, 24, 1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elkhawas, Y.A.; Elissawy, A.M.; Elnaggar, M.S.; Mostafa, N.M.; Kamal, E.M.; Bishr, M.M.; Singab, A.N.B.; Salama, O.M. Chemical diversity in species belonging to soft coral genus Sacrophyton and its impact on biological activity: A review. Mar. Drugs 2020, 18, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurrachma, M.Y.; Sakaraga, D.; Nugraha, A.Y.; Rahmawati, S.I.; Bayu, A.; Sukmarini, L.; Atikana, A.; Prasetyoputri, A.; Izzati, F.; Warsito, M.F.; et al. Cembranoids of soft corals: Recent updates and their biological activities. Nat. Prod. Bioprospect. 2021, 11, 243–306. [Google Scholar] [CrossRef]
- Yan, X.; Liu, J.; Leng, X.; Ouyang, H. Chemical diversity and biological activity of secondary metabolites from soft coral genus Sinularia since 2013. Mar. Drugs 2021, 19, 335. [Google Scholar] [CrossRef]
- Khalesi, M.; Beeftink, H.; Wijffels, R. The soft coral Sinularia flexibilis: Potential for drug development. Adv. Coral Husb. Public Aquar. 2001, 2, 47–60. [Google Scholar]
- Nagappan, T.; Palaniveloo, K. Biological properties and chemical diversity of Sinularia Flexibilis, an Alcyonacean Soft Coral. J. Sustain. Sci. Manag. 2018, 13, 15–34. [Google Scholar]
- Ermolenko, E.V.; Imbs, A.B.; Gloriozova, T.A.; Poroikov, V.V.; Sikorskaya, T.V.; Dembitsky, V.M. Chemical diversity of soft coral steroids and their pharmacological activities. Mar. Drugs 2020, 18, 613. [Google Scholar] [CrossRef]
- Savic, M.P.; Sakac, M.N.; Kuzminac, I.Z.; Ajdukovic, J.J. Structural diversity of bioactive steroid compounds isolated from soft corals in the period 2015-2020. J. Steroid Biochem. Mol. Biol. 2022, 218, 106061. [Google Scholar] [CrossRef]
- Li, W.; Zou, Y.H.; Ge, M.X.; Lou, L.L.; Xu, Y.S.; Ahmed, A.; Chen, Y.Y.; Zhang, J.S.; Tang, G.H.; Yin, S. Biscembranoids and cembranoids from the soft coral Sarcophyton elegans. Mar. Drugs 2017, 15, 85. [Google Scholar] [CrossRef]
- Wei, W.C.; Sung, P.J.; Duh, C.Y.; Chen, B.W.; Sheu, J.H.; Yang, N.S. Anti-inflammatory activities of natural products isolated from soft corals of Taiwan between 2008 and 2012. Mar. Drugs 2013, 11, 4083–4126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putra, M.Y.; Murniasih, T. Marine soft corals as source of lead compounds for anti-inflammatories. J. Coast. Life Med. 2016, 4, 73–77. [Google Scholar] [CrossRef]
- Frichert, A.; Jones, P.G.; Lindel, T. Synthesis of eunicellane-type bicycles embedding a 1,3-cyclohexadiene moiety. Beilstein J. Org. Chem. 2018, 14, 2461–2467. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-W.; Wu, Y.-C.; Chiang, M.Y.; Su, J.-H.; Wang, W.-H.; Fan, T.-Y.; Sheu, J.-H. Eunicellin-based diterpenoids from the cultured soft coral Klyxum simplex. Tetrahedron 2009, 65, 7016–7022. [Google Scholar] [CrossRef]
- Chen, B.W.; Chao, C.H.; Su, J.H.; Tsai, C.W.; Wang, W.H.; Wen, Z.H.; Huang, C.Y.; Sung, P.J.; Wu, Y.C.; Sheu, J.H. Klysimplexins I-T, eunicellin-based diterpenoids from the cultured soft coral Klyxum simplex. Org. Biomol. Chem. 2011, 9, 834–844. [Google Scholar] [CrossRef]
- Chen, B.-W.; Huang, C.-Y.; Wen, Z.-H.; Su, J.-H.; Wang, W.-H.; Sung, P.-J.; Wu, Y.-C. Klysimplexins U–X, eunicellin-based diterpenoids from the cultured soft coral Klyxum simplex. Bull. Chem. Soc. Jpn. 2011, 84, 1237–1242. [Google Scholar] [CrossRef]
- Chen, B.W.; Chao, C.H.; Su, J.H.; Wen, Z.H.; Sung, P.J.; Sheu, J.H. Anti-inflammatory eunicellin-based diterpenoids from the cultured soft coral Klyxum simplex. Org. Biomol. Chem. 2010, 8, 2363–2366. [Google Scholar] [CrossRef]
- Taglialatela-Scafati, O.; Deo-Jangra, U.; Campbell, M.; Roberge, M.; Andersen, R.J. Diterpenoids from cultured Erythropodium caribaeorum. Org. Lett. 2002, 4, 4085–4088. [Google Scholar] [CrossRef]
- Su, J.-H.; Lin, Y.-F.; Lu, Y.; Yeh, H.-C.; Wang, W.-H.; Fan, T.-Y.; Sheu, J.-H. Oxygenated cembranoids from the cultured and wild-type soft corals Sinularia flexibilis. Chem. Pharm. Bull. 2009, 57, 1189–1192. [Google Scholar] [CrossRef] [Green Version]
- Kashman, Y.; Bodner, M.; Loya, Y.; Benayahu, Y. Cembranolids from marine origin (Red Sea), survey, and isolation of a new sinulariolide derivative. Isr. J. Chem. 1977, 16, 1–3. [Google Scholar] [CrossRef]
- Chen, N.F.; Huang, S.Y.; Lu, C.H.; Chen, C.L.; Feng, C.W.; Chen, C.H.; Hung, H.C.; Lin, Y.Y.; Sung, P.J.; Sung, C.S.; et al. Flexibilide obtained from cultured soft coral has anti-neuroinflammatory and analgesic effects through the upregulation of spinal transforming growth factor-beta1 in neuropathic rats. Mar. Drugs 2014, 12, 3792–3817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, T.C.; Chen, H.Y.; Sheu, J.H.; Chiang, M.Y.; Wen, Z.H.; Dai, C.F.; Su, J.H. Structural elucidation and structure-anti-inflammatory activity relationships of cembranoids from cultured soft corals Sinularia sandensis and Sinularia flexibilis. J. Agric. Food Chem. 2015, 63, 7211–7218. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.W.; Chung, H.L.; Wang, C.C.; Su, J.H.; Chen, Y.J.; Lee, C.J. Anti-acne effects of cembrene diterpenoids from the cultured soft coral Sinularia flexibilis. Mar. Drugs 2020, 18, 487. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-F.; Su, H.-J.; Lee, N.-L.; Su, J.-H. Cembranoids from the cultured soft coral Sinularia gibberosa. Nat. Prod. Commun. 2013, 10, 1363–1364. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, T.H.; Sung, C.S.; Lan, Y.H.; Wang, Y.C.; Lu, M.C.; Wen, Z.H.; Wu, Y.C.; Sung, P.J. New anti-inflammatory cembranes from the cultured soft coral Nephthea columnaris. Mar. Drugs 2015, 13, 3443–3453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, T.H.; Cheng, C.H.; Wu, T.Y.; Lu, M.C.; Chen, W.F.; Wen, Z.H.; Dai, C.F.; Wu, Y.C.; Sung, P.J. New cembranoid diterpenes from the cultured octocoral Nephthea columnaris. Molecules 2015, 20, 13205–13215. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.L.; Su, J.H. Tetrahydrofuran cembranoids from the cultured soft coral Lobophytum crassum. Mar. Drugs 2011, 9, 2526–2536. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.Y.; Sung, P.J.; Uvarani, C.; Su, J.H.; Lu, M.C.; Hwang, T.L.; Dai, C.F.; Wu, S.L.; Sheu, J.H. Glaucumolides A and B, biscembranoids with new structural type from a cultured soft coral Sarcophyton glaucum. Sci. Rep. 2015, 5, 15624. [Google Scholar] [CrossRef] [Green Version]
- Su, J.-H.; Lu, Y.; Lin, W.-Y.; Wang, W.-H.; Sung, P.-J.; Sheu, J.-H. A cembranoid, trocheliophorol, from the cultured soft coral Sarcophyton trocheliophorum. Chem. Lett. 2010, 39, 172–173. [Google Scholar] [CrossRef]
- Huang, P.C.; Lin, W.S.; Peng, B.R.; Chang, Y.C.; Fang, L.S.; Li, G.Q.; Hwang, T.L.; Wen, Z.H.; Sung, P.J. New furanocembranoids from Briareum violaceum. Mar. Drugs 2019, 17, 214. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.-C.; Tseng, C.-C.; Peng, B.-R.; Hu, C.-C.; Lin, N.-C.; Chen, N.-F.; Chen, J.-J.; Wen, Z.-H.; Wu, Y.-C.; Sung, P.-J. Briaviodiols B–E, new anti-inflammatory hydroperoxyfurancembranoids from Briareum violaceum. Tetrahedron 2019, 75, 921–927. [Google Scholar] [CrossRef]
- Liu, L.; Zheng, Y.-Y.; Shao, C.-L.; Wang, C.-Y. Metabolites from marine invertebrates and their symbiotic microorganisms: Molecular diversity discovery, mining, and application. Mar. Life Sci. Technol. 2019, 1, 60–94. [Google Scholar] [CrossRef]
- Tsai, T.C.; Wu, Y.J.; Su, J.H.; Lin, W.T.; Lin, Y.S. A new spatane diterpenoid from the cultured soft coral Sinularia leptoclados. Mar. Drugs 2013, 11, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.-T.; Li, Y.; Guo, Y.-W. Terpenoids of Sinularia soft corals: Chemistry and bioactivity. Acta Pharm. Sin. B 2012, 2, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.F.; Yin, C.T.; Cheng, C.H.; Lu, M.C.; Fang, L.S.; Wang, W.H.; Wen, Z.H.; Chen, J.J.; Wu, Y.C.; Sung, P.J. Norcembranoidal diterpenes from the cultured-type octocoral Sinularia numerosa. Int. J. Mol. Sci. 2015, 16, 3298–3306. [Google Scholar] [CrossRef] [Green Version]
- Chi, W.-C.; Kuo, L.-M.; Yang, S.-N.; Lee, Y.-T.; Wen, Z.-H.; Tsui, K.-H.; Hwang, T.-L.; Zhang, Y.-L.; Sung, P.-J. Briarenols O and P: Novel briaranes from a cultured octocoral Briareum excavatum (Briareidae). Phytochem. Lett. 2021, 41, 134–138. [Google Scholar] [CrossRef]
- Sung, P.-J.; Lin, M.-R.; Chiang, M.Y.; Huang, I.C.; Syu, S.-M.; Fang, L.-S.; Wang, W.-H.; Sheu, J.-H. Briarenolide D, a new hydroperoxybriarane diterpenoid from a cultured octocoral Briareum sp. Chem. Lett. 2010, 39, 1030–1032. [Google Scholar] [CrossRef]
- Sung, P.-J.; Lin, M.-R.; Su, Y.-D.; Chiang, M.Y.; Hu, W.-P.; Su, J.-H.; Cheng, M.-C.; Hwang, T.-L.; Sheu, J.-H. New briaranes from the octocorals Briareum excavatum (Briareidae) and Junceella fragilis (Ellisellidae). Tetrahedron 2008, 64, 2596–2604. [Google Scholar] [CrossRef]
- Sung, P.J.; Lin, M.R.; Hwang, T.L.; Fan, T.Y.; Su, W.C.; Ho, C.C.; Fang, L.S.; Wang, W.H. Briaexcavatins M-P, four new briarane-related diterpenoids from cultured octocoral Briareum excavatum (Briareidae). Chem. Pharm. Bull. 2008, 56, 930–935. [Google Scholar] [CrossRef] [Green Version]
- Hwang, T.-L.; Lin, M.-R.; Tsai, W.-T.; Yeh, H.-C.; Hu, W.-P.; Sheu, J.-H.; Sung, P.-J. New polyoxygenated briaranes from octocorals Briareum excavatum and Ellisella robusta. Bull. Chem. Soc. Jpn. 2008, 81, 1638–1646. [Google Scholar] [CrossRef]
- Sung, P.-J.; Lin, M.-R.; Chiang, M.Y. The structure and absolute stereochemistry of briaexcavatin U, a new chlorinated briarane from a cultured octocoral Briareum excavatum. Chem. Lett. 2009, 38, 154–155. [Google Scholar] [CrossRef]
- Sung, P.-J.; Lin, M.-R.; Chiang, M.Y.; Hwang, T.-L. Briaexcavatins V–Z, discovery of new briaranes from a cultured octocoral Briareum excavatum. Bull. Chem. Soc. Jpn. 2009, 82, 987–996. [Google Scholar] [CrossRef]
- Sung, P.-J.; Su, Y.-D.; Li, G.-Y.; Chiang, M.Y.; Lin, M.-R.; Huang, I.C.; Li, J.-J.; Fang, L.-S.; Wang, W.-H. Excavatoids A–D, new polyoxygenated briaranes from the octocoral Briareum excavatum. Tetrahedron 2009, 65, 6918–6924. [Google Scholar] [CrossRef]
- Sung, P.J.; Chen, B.Y.; Lin, M.R.; Hwang, T.L.; Wang, W.H.; Sheu, J.H.; Wu, Y.C. Excavatoids E and F: Discovery of two new briaranes from the cultured octocoral Briareum excavatum. Mar. Drugs 2009, 7, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Sung, P.-J.; Chen, B.-Y.; Chiang, M.Y.; Hou, C.-H.; Su, Y.-D.; Hwang, T.-L.; Chen, Y.-H.; Chen, J.-J. Excavatoids G–K, new 8,17-epoxybriaranes from the cultured octocoral Briareum excavatum (Briareidae). Bull. Chem. Soc. Jpn. 2010, 83, 539–545. [Google Scholar] [CrossRef]
- Su, J.H.; Chen, B.Y.; Hwang, T.-L.; Chen, Y.-H.; Huang, I.C.; Lin, M.-R.; Chen, J.-J.; Fang, L.-S.; Wang, W.-H.; Li, J.-J.; et al. Excavatoids L-N, new 12-hydroxybriaranes from the cultured octocoral Briareum excavatum (Briareidae). Chem. Pharm. Bull. 2010, 5, 662–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, P.J.; Li, G.Y.; Su, Y.D.; Lin, M.R.; Chang, Y.C.; Kung, T.H.; Lin, C.S.; Chen, Y.H.; Su, J.H.; Lu, M.C.; et al. Excavatoids O and P, new 12-hydroxybriaranes from the octocoral Briareum excavatum. Mar. Drugs 2010, 8, 2639–2646. [Google Scholar] [CrossRef]
- Xu, J.H.; Lai, K.H.; Su, Y.D.; Chang, Y.C.; Peng, B.R.; Backlund, A.; Wen, Z.H.; Sung, P.J. Briaviolides K-N, new briarane-type diterpenoids from cultured octocoral Briareum violaceum. Mar. Drugs 2018, 16, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.-H.; Chang, Y.-C.; Li, G.-Q.; Wen, Z.-H.; Wu, Y.-C.; Sung, P.-J. Briaviolides O and P, new briaranes from a cultured octocoral Briareum violaceum. Phytochem. Lett. 2018, 27, 129–133. [Google Scholar] [CrossRef]
- Tsai, T.-C.; Xu, J.-H.; Li, M.-J.; Chen, J.-J.; Su, J.-H.; Wu, Y.-C.; Wen, Z.-H.; Sung, P.-J. Briaviolide Q, a new briarane from the cultured Briareum violaceum. Nat. Prod. Comm. 2018, 13, 1235–1237. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.Y.; Lin, S.C.; Feng, C.W.; Chen, P.C.; Su, Y.D.; Li, C.M.; Yang, S.N.; Jean, Y.H.; Sung, P.J.; Duh, C.Y.; et al. Anti-inflammatory and analgesic effects of the marine-derived compound excavatolide b isolated from the culture-type formosan gorgonian Briareum excavatum. Mar. Drugs 2015, 13, 2559–2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.Y.; Jean, Y.H.; Lee, H.P.; Lin, S.C.; Pan, C.Y.; Chen, W.F.; Wu, S.F.; Su, J.H.; Tsui, K.H.; Sheu, J.H.; et al. Excavatolide B attenuates rheumatoid arthritis through the inhibition of osteoclastogenesis. Mar. Drugs 2017, 15, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L.; Chiang, C.C.; Lee, Y.T.; Wen, Z.H.; Wu, Y.C.; Wu, Y.J.; Hwang, T.L.; Wu, T.Y.; Chang, C.Y.; Sung, P.J. Briarenols Q-T: Briaranes from a cultured octocoral Briareum stechei (Kukenthal, 1908). Mar. Drugs 2020, 18, 383. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Zhang, Y.L.; Lee, G.H.; Tsou, L.K.; Zhang, M.M.; Hsieh, H.P.; Chen, J.J.; Ko, C.Y.; Wen, Z.H.; Sung, P.J. Briarenols W-Z: Chlorine-containing polyoxygenated briaranes from octocoral Briareum stechei (Kukenthal, 1908). Mar. Drugs 2021, 19, 77. [Google Scholar] [CrossRef] [PubMed]
- Huynh, T.H.; Wen, Z.-H.; Chien, S.-Y.; Chung, H.-M.; Su, J.-H.; Fang, L.-S.; Wu, Y.-J.; Lin, S.-H.; Sung, P.-J. Briavioids A–C, discovery of new polyacetoxybriaranes from octocoral Briareum violaceum (Quoy & Gaimard, 1833). Tetrahedron 2022, in press. [Google Scholar]
- Thornton, P.D.; Burnell, D.J. A Pauson−Khand and ring-expansion approach to the aquariane ring system. Org. Lett. 2006, 8, 3195–3198. [Google Scholar] [CrossRef] [PubMed]
- Phan, G.H.; Tsai, Y.-C.; Liu, Y.-H.; Fang, L.-S.; Wen, Z.-H.; Hwang, T.-L.; Chang, Y.-C.; Sung, P.-J. Sterol constituents from a cultured octocoral Sinularia sandensis (Verseveldt 1977). J. Mol. Struct. 2021, 1246, 131175. [Google Scholar] [CrossRef]
- Huang, C.Y.; Liaw, C.C.; Chen, B.W.; Chen, P.C.; Su, J.H.; Sung, P.J.; Dai, C.F.; Chiang, M.Y.; Sheu, J.H. Withanolide-based steroids from the cultured soft coral Sinularia brassica. J. Nat. Prod. 2013, 76, 1902–1908. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Ahmed, A.F.; Su, J.H.; Sung, P.J.; Hwang, T.L.; Chiang, P.L.; Dai, C.F.; Liaw, C.C.; Sheu, J.H. Bioactive new withanolides from the cultured soft coral Sinularia brassica. Bioorg. Med. Chem. Lett. 2017, 27, 3267–3271. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Su, J.H.; Liaw, C.C.; Sung, P.J.; Chiang, P.L.; Hwang, T.L.; Dai, C.F.; Sheu, J.H. Bioactive steroids with methyl ester group in the side chain from a reef soft coral Sinularia brassica cultured in a tank. Mar. Drugs 2017, 15, 280. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.F.; Kuo, C.Y.; Wen, Z.H.; Lin, Y.Y.; Wang, W.H.; Su, J.H.; Sheu, J.H.; Sung, P.J. Flexibilisquinone, a new anti-inflammatory quinone from the cultured soft coral Sinularia flexibilis. Molecules 2013, 18, 8160–8167. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.J.; Su, J.H.; Tsai, C.C.; Chen, Y.J.; Liao, M.H.; Wu, Y.J. 11-epi-Sinulariolide acetate reduces cell migration and invasion of human hepatocellular carcinoma by reducing the activation of ERK1/2, p38MAPK and FAK/PI3K/AKT/mTOR signaling pathways. Mar. Drugs 2014, 12, 4783–4798. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.J.; Wang, R.Y.; Chen, J.C.; Chiu, C.C.; Liao, M.H.; Wu, Y.J. Cytotoxicity of 11-epi-sinulariolide acetate isolated from cultured soft corals on ha22t cells through the endoplasmic reticulum stress pathway and mitochondrial dysfunction. Int. J. Mol. Sci. 2016, 17, 1787. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.I.; Chen, C.C.; Chen, J.C.; Su, J.H.; Huang, H.H.; Chen, J.Y.; Wu, Y.J. Proteomic analysis of anti-tumor effects of 11-dehydrosinulariolide on CAL-27 cells. Mar. Drugs 2011, 9, 1254–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.I.; Wang, R.Y.; Lin, J.J.; Su, J.H.; Chiu, C.C.; Chen, J.C.; Chen, J.Y.; Wu, Y.J. Proteomic profiling of the 11-dehydrosinulariolide-treated oral carcinoma cells Ca9-22: Effects on the cell apoptosis through mitochondrial-related and ER stress pathway. J. Proteomics 2012, 75, 5578–5589. [Google Scholar] [CrossRef] [PubMed]
- Su, T.R.; Tsai, F.J.; Lin, J.J.; Huang, H.H.; Chiu, C.C.; Su, J.H.; Yang, Y.T.; Chen, J.Y.; Wong, B.S.; Wu, Y.J. Induction of apoptosis by 11-dehydrosinulariolide via mitochondrial dysregulation and ER stress pathways in human melanoma cells. Mar. Drugs 2012, 10, 1883–1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.C.; Su, J.H.; Lin, S.C.; Chang, C.C.; Hsia, T.C.; Tung, Y.T.; Lin, C.C. A soft coral-derived compound, 11-dehydrosinulariolide, induces G2/M cell cycle arrest and apoptosis in small cell lung cancer. Mar. Drugs 2018, 16, 479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheu, J.-H.; Sung, P.-J.; Huang, L.-H.; Lee, S.-F.; Wu, T.; Chang, B.-Y.; Duh, C.-Y.; Fang, L.-S.; Soong, K.; Lee, T.-J. New cytotoxic briaran diterpenes from the formosan gorgonian Briareum sp. J. Nat. Prod. 1996, 59, 935–938. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.Y.; Jean, Y.H.; Lee, H.P.; Chen, W.F.; Sun, Y.M.; Su, J.H.; Lu, Y.; Huang, S.Y.; Hung, H.C.; Sung, P.J.; et al. A soft coral-derived compound, 11-epi-sinulariolide acetate suppresses inflammatory response and bone destruction in adjuvant-induced arthritis. PLoS ONE 2013, 8, e62926. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.Y.; Chen, N.F.; Chen, W.F.; Hung, H.C.; Lee, H.P.; Lin, Y.Y.; Wang, H.M.; Sung, P.J.; Sheu, J.H.; Wen, Z.H. Sinularin from indigenous soft coral attenuates nociceptive responses and spinal neuroinflammation in carrageenan-induced inflammatory rat model. Mar. Drugs 2012, 10, 1899–1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.F.; Chakraborty, C.; Sung, C.S.; Feng, C.W.; Jean, Y.H.; Lin, Y.Y.; Hung, H.C.; Huang, T.Y.; Huang, S.Y.; Su, T.M.; et al. Neuroprotection by marine-derived compound, 11-dehydrosinulariolide, in an in vitro Parkinson’s model: A promising candidate for the treatment of Parkinson’s disease. Naunyn Schmiedebergs Arch. Pharmacol. 2012, 385, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Weinheimer, A.J.; Matson, J.A.; Hossain, M.B.; van der Helm, D. Marine anticancer agents: Sinularin and dihydrosinularin, new cembranolides from the soft coral, Sinularia flexibilis. Tetrahedron Lett. 1977, 18, 2923–2926. [Google Scholar] [CrossRef]
- Li, H.H.; Su, J.H.; Chiu, C.C.; Lin, J.J.; Yang, Z.Y.; Hwang, W.I.; Chen, Y.K.; Lo, Y.H.; Wu, Y.J. Proteomic investigation of the sinulariolide-treated melanoma cells A375: Effects on the cell apoptosis through mitochondrial-related pathway and activation of caspase cascade. Mar. Drugs 2013, 11, 2625–2642. [Google Scholar] [CrossRef] [PubMed]
- Goffredo, S.; Lasker, H.R. An adaptive management approach to an octocoral fishery based on the Beverton-Holt model. Coral Reefs 2008, 27, 751–761. [Google Scholar] [CrossRef]
- Thatcher, C.; Hoj, L.; Bourne, D.G. Probiotics for coral aquaculture: Challenges and considerations. Curr. Opin. Biotechnol. 2022, 73, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Montaser, R.; Luesch, H. Marine natural products: A new wave of drugs? Future Med. Chem. 2011, 3, 1475–1489. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Chemical Classification | Novelty | Source | Bioactivities |
|---|---|---|---|---|
| Klysimplexin B (2) | Eunicellin-based diterpene | New | Klyxum simplex | Exhibited moderate cytotoxicity against Hep G2, Hep 3B, MDA-MB-231, MCF-7, A549, and Ca9-22 cell lines (IC50 = 3.0, 3.6, 6.9, 3.0, 2.0, and 1.8 μg/mL, respectively) [24] |
| Klysimplexin H (8) | Eunicellin-based diterpene | New | Klyxum simplex | Showed cytotoxicity against Hep G2, Hep 3B, MDA-MB-231, MCF-7, A549, and Ca9-22 cell lines (IC50 = 5.6, 6.9, 4.4, 5.6, 2.8 and 6.1 μg/mL, respectively) [24] |
| Klysimplexin J (10) | Eunicellin-based diterpene | New | Klyxum simplex | Significantly reduced the expression of iNOS protein at 10 μM [25] |
| Klysimplexin K (11) | Eunicellin-based diterpene | New | Klyxum simplex | Significantly reduced the expression of iNOS protein at 10 μM [25] |
| Klysimplexin L (12) | Eunicellin-based diterpene | New | Klyxum simplex | Significantly reduced the expression of iNOS protein at 10 μM [25] |
| Klysimplexin M (13) | Eunicellin-based diterpene | New | Klyxum simplex | Significantly reduced the expression of iNOS protein at 10 μM [25] |
| Klysimplexin N (14) | Eunicellin-based diterpene | New | Klyxum simplex | Significantly reduced the expression of iNOS protein at 10 μM [25] |
| Klysimplexin Q (17) | Eunicellin-based diterpene | New | Klyxum simplex | Exhibited cytotoxicity against Hep G2, Hep 3B, MDA-MB-231, MCF-7, A549, and Ca9-22 cell lines (IC50 = 53.2, 35.1, 44.0, 36.5, 40.5, and 40.5 μM, respectively) [25] |
| Klysimplexin R (18) | Eunicellin-based diterpene | New | Klyxum simplex | Significantly reduced the expression of iNOS and COX-2 proteins at 10 μM [25] |
| Klysimplexin S (19) | Eunicellin-based diterpene | New | Klyxum simplex | Significantly reduced the expression of iNOS and COX-2 proteins at 10 μM [25] |
| Klysimplexin T (20) | Eunicellin-based diterpene | New | Klyxum simplex | Showed cytotoxicity against the growth of Hep G2, Hep 3B, MDA-MB-231, MCF-7, A549, and Ca9-22 cells (IC50 = 34.3, 26.4, 44.0, 27.2, 42.0 and 37.4 μM, respectively) [25] |
| Klysimplexin sulfoxide A (25) | Eunicellin-based diterpene | New | Klyxum simplex | Significantly reduced the expression of iNOS protein at 10 μM [27] |
| Klysimplexin sulfoxide B (26) | Eunicellin-based diterpene | New | Klyxum simplex | Significantly reduced the expression of iNOS protein at 10 μM [27] |
| Klysimplexin sulfoxide C (27) | Eunicellin-based diterpene | New | Klyxum simplex | Significantly reduced the expression of iNOS and COX-2 proteins at 10 μM [27] |
| 11-epi-Sinulariolide acetate (33) | Cembrane-type diterpene | Known | Sinularia flexibilis | Exhibited weak cytotoxicity against the proliferation of MCF-7 cells (ED50 = 11.5 μg/mL) [29] Showed a significant antiproliferative effect and inhibitory activity on cell migration and invasion in HA22T cells in a concentration-dependent manner [72] Induced apoptosis in HA22T cells [73] Significantly reduced iNOS levels to 84.89 ± 8.23%, 39.89 ± 5.64%, 11.8 ± 1.03%, and 1.4 ± 1.74% at concentrations of 1, 10, 25, and 50 μM, respectively [79] Significantly reduced COX-2 levels to 82.89 ± 1.63%, 65.93 ± 4.22%, 52.63 ± 4.76%, and 42.13 ± 3.25% at 10, 25, and 50 μM, respectively [79] Anti-rheumatic effect: Significantly reduced the clinical characteristics and the expressions of osteoclast-related proteins and improved the histopathologic features in the AIA rat model [79] |
| Flexibilide (sinularin) (34) | Cembrane-type diterpene | Known | Sinularia flexibilis | Exhibited significant cytotoxicity against KB and PS cell lines (ED50 = 0.3 μg/mL) [82] Significantly reduced iNOS levels to 53.45 ± 3.27%, 36.45 ± 5.15%, 33.38 ± 4.61%, and 19.48 ± 3.95% at 0.1, 1, 10, and 20 μM, respectively [80] Significantly reduced the levels of COX-2 protein to 82.72 ± 6.17% and 66.23 ± 3.27% at 0.1, 1, 10, and 20 μM, respectively [80] Significantly increased the levels of TGF-β protein to 137.75 ± 5.97%, 149.82 ± 6.15%, 142.71 ± 4.57%, and 138.02 ± 5.15% at 0.1, 1, 10, and 20 μM, respectively [80] Exhibited analgesic properties in a rat model at 80 mg/kg [80] Exerted anti-neuroinflammatory and analgesic effects in a rat model [31] Anti-acne capabilities [33] |
| Isosinulaflexiolide K (37) | Cembrane-type diterpene | New | Sinularia flexibilis | Significantly reduced the levels of iNOS and COX-2 proteins to 30.9 ± 4.1% and 47.1 ± 3.8%, respectively, at 10 μM [32] |
| Sinulaflexiolide K (39) | Cembrane-type diterpene | Known | Sinularia flexibilis | Significantly reduced the levels of iNOS and COX-2 proteins to 37.4 ± 5.9% and 51.4 ± 5.6%, respectively, at 10 μM [32] |
| (−)-Sandensolide (40) | Cembrane-type diterpene | Known | Sinularia flexibilis | Significantly reduced the levels of iNOS and COX-2 proteins to 61 ± 3.4% and 51.9 ± 7.2%, respectively, at 10 μM [32] |
| 11-Dehydrosinulariolide (41) | Cembrane-type diterpene | Known | Sinularia flexibilis | Reduced the cell viability to 70% and significantly induced both the early and late apoptosis of CAL-27 cells at a concentration of 1.5 µg/mL [74] Reduced the cell viability to 60% and significantly induced both the early and late apoptosis of Ca9-22 cells at a concentration of 3.0 µg/mL [75] Inhibited cell migration of CAL-27 and Ca9-22 cells in a dose-dependent manner [74,75] Exhibited a dose-dependent (2–8 μg/mL) cytotoxicity against A2058 cells (IC50 = 5.8 μg/mL) [76] Exhibited a dose-dependent anti-migratory effect against A2058 cells with the suppression rates of approximately 32%, 51% and 73% for 2, 4, and 6 μg/mL, respectively [76] Exhibited cytotoxic against H1688 and H146 cell (after 12, 24, and 48 h of exposure, IC50 = >50, 29.8 ± 3.4, and 19.1 ± 2.4 µM respectively, for H1688 cells, and > 50, 43.5 ± 6.6, and 25.1 ± 2.6 µM, respectively, for H146 cells) [77] Significantly suppressed the H1688 tumor growth in a mouse xenograft model with an intraperitoneal injection regimen of 10 mg/kg x 3 times/week for 22 days [77] Significantly reduced the levels of iNOS and COX-2 proteins to 31.9 ± 5.1% and 49 ± 5.6%, respectively, at 10 μM [32] Anti-acne capabilities [33] Neuroprotective effect [81] |
| Sinulariolide (42) | Cembrane-type diterpene | Known | Sinularia flexibilis | Exhibited weak cytotoxicity in MCF-7 cells (ED50 = 16.9 μg/mL) [29] Exhibited cytotoxicity against KB and PS cell lines (ED50 = 20 and 7 μg/mL, respectively) [82] Inhibited cell proliferation, suppressed A375 melanoma cell migration, and elevated early and late apoptosis in a concentration-dependent manner [83] Significantly reduced the levels of iNOS and COX-2 proteins to 47.7 ± 6.3% and 52.2 ± 5.1%, respectively, at 10 μM [32] |
| 3,4:8,11-Bisepoxy-7-acetoxycembra-15(17)-en-1,12-olide (43) | Cembrane-type diterpene | Known | Sinularia flexibilis | Significantly reduced the levels of iNOS and COX-2 proteins to 25.7 ± 5.2% and 55.3 ± 8.2%, respectively, at 10 μM [32] Anti-acne capabilities [33] |
| Dihydrosinularin (46) | Cembrane-type diterpene | Known | Sinularia flexibilis | Exhibited cytotoxicity against KB and PS cell lines (ED50 = 16 and 1.1 μg/mL, respectively) [82] |
| 11,12-Epoxy-13,14-dihydroxycembrene-C (48) | Cembrane-type diterpene | Known | Sinularia gibberosa | Exhibited significant antibacterial activity at a concentration of 25 μg/disk [34] |
| Flaccidoxide (49) | Cembrane-type diterpene | Known | Sinularia gibberosa | Exhibited significant antibacterial activity at a concentration of 50 μg/disk [34] |
| Columnariol A (56) | Cembrane-type diterpene | New | Litophyton columnaris | Exhibited moderate cytotoxicity toward LNCaP cells (IC50 = 9.80 μg/mL) [35] Significantly reduced the levels of iNOS and COX-2 at 50 μM [35] |
| Columnariol B (57) | Cembrane-type diterpene | New | Litophyton columnaris | Significantly reduced the levels of iNOS and COX-2 at 50 μM [35] |
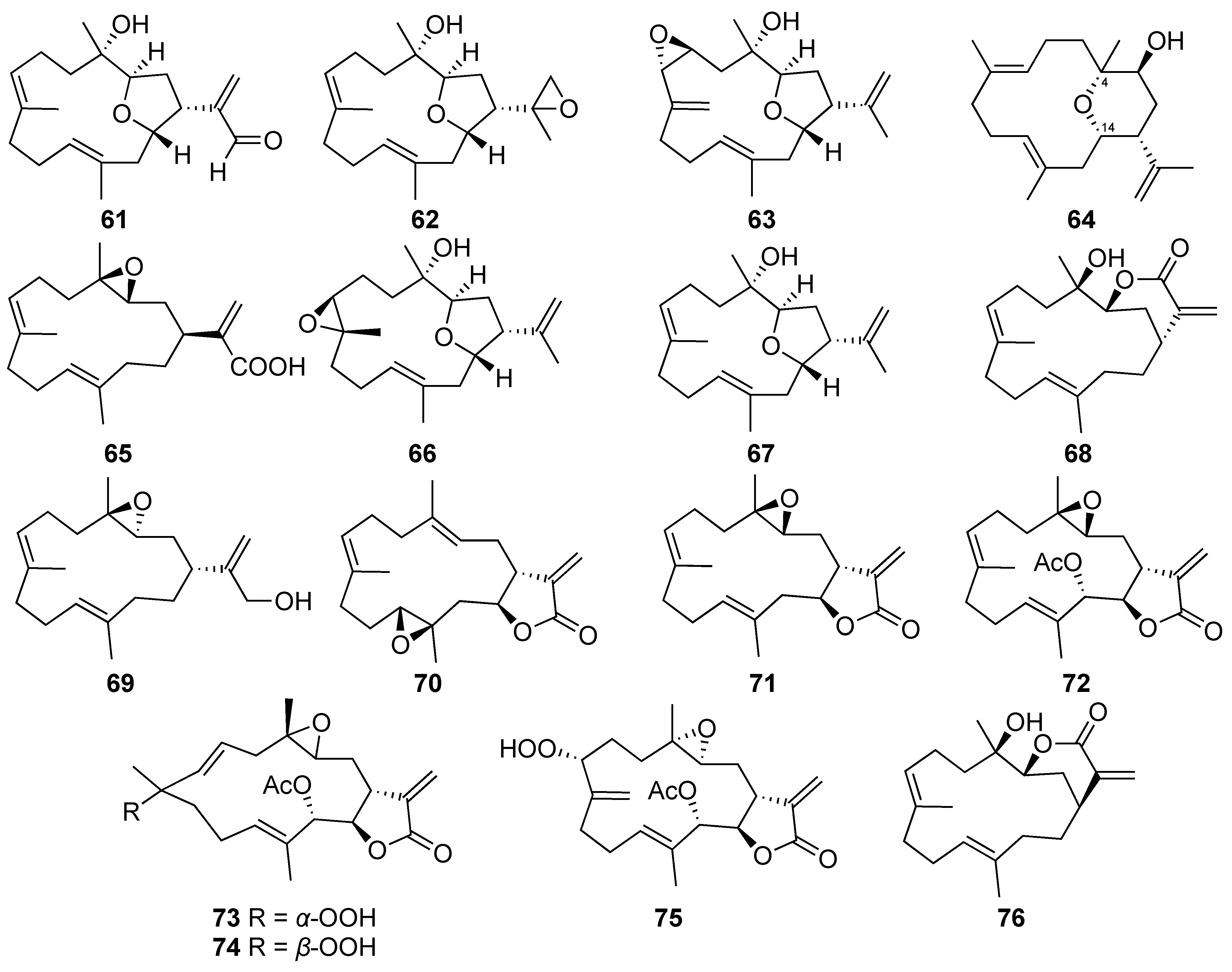
| Culobophylin A (61) | Cembrane-type diterpenoid | New | Lobophytum crassum | Exhibited cytotoxicity against the HL60, MDA-MB-231, DLD-1, and HCT-116 cells (IC50 = 3.0, 16.8, 4.6, and 16.3 μg/mL, respectively) [37] |
| Culobophylin B (62) | Cembrane-type diterpenoid | New | Lobophytum crassum | Exhibited cytotoxicity against HL60, DLD-1 and HCT-116 cells (IC50 = 6.8, 16.2, and 16.7 μg/mL, respectively) [37] |
| Lobophylin B (67) | Cembrane-type diterpenoid | Known | Lobophytum crassum | Exhibited cytotoxicity against K562, Molt 4, and Sup-T1 cells (IC50 = 16.3, 12.3, and 4.6 μM, respectively) [7] |
| Lobocrassin B (68) | Cembrane-type diterpenoid | Known | Lobophytum crassum | Exhibited cytotoxicity against K562, Molt 4, U937, and Sup-T1 cells (IC50 =3.3, 2.3, 5.2, and 6.2 μM, respectively) [7] |
| Lobocrassin C (69) | Cembrane-type diterpenoid | Known | Lobophytum crassum | Exhibited cytotoxicity toward Sup-T1 cells (IC50 = 35.8 μM) [7] |
| Crassocolide E (70) | Cembrane-type diterpenoid | Known | Lobophytum crassum | Exhibited cytotoxicity against K562, Molt 4, U937, and Sup-T1 cells (IC50 = 11.3, 6.2, 15.8, and 5.2 μM, respectively) [7] |
| Sarcocrassolide (71) | Cembrane-type diterpenoid | Known | Lobophytum crassum | Exhibited cytotoxicity against K562, Molt 4, U937, and Sup-T1 cells (IC50 = 18.1, 8.4, 4.4, and 8.3 μM, respectively) [7] |
| 13-Acetoxysarcocrassolide (72) | Cembrane-type diterpenoid | Known | Lobophytum crassum | Exhibited cytotoxicity against K562, Molt 4, U937, and Sup-T1 cells (IC50 = 3.3, 1.2, 7.1, and 1.5 μM, respectively) [7] |
| Sarcocrassocolide F (73) | Cembrane-type diterpenoid | Known | Lobophytum crassum | Exhibited cytotoxicity toward K562, Molt 4, U937, and Sup-T1 cells (IC50 =12.3, 4.8, 10.9, and 6.1 μM, respectively) [7] |
| Sarcocrassocolide G (74) | Cembrane-type diterpenoid | Known | Lobophytum crassum | Exhibited cytotoxicity against K562, Molt 4, U937, and Sup-T1 cells (IC50 =13.0, 7.0, 23.3, and 6.6 μM, respectively) [7] |
| Sarcocrassocolide M (75) | Cembrane-type diterpenoid | Known | Lobophytum crassum | Exhibited cytotoxicity against K562, Molt 4, U937, and Sup-T1 cells (IC50 =15.3, 11.6, 32.0, and 10.2 μM, respectively) [7] |
| 14-Deoxycrassin (76) | Cembrane-type diterpenoid | Known | Lobophytum crassum | Exhibited cytotoxicity against K562, Molt 4, U937, and Sup-T1 cells (IC50 =4.5, 2.9, 7.0, and 4.5 μM, respectively) [7] |
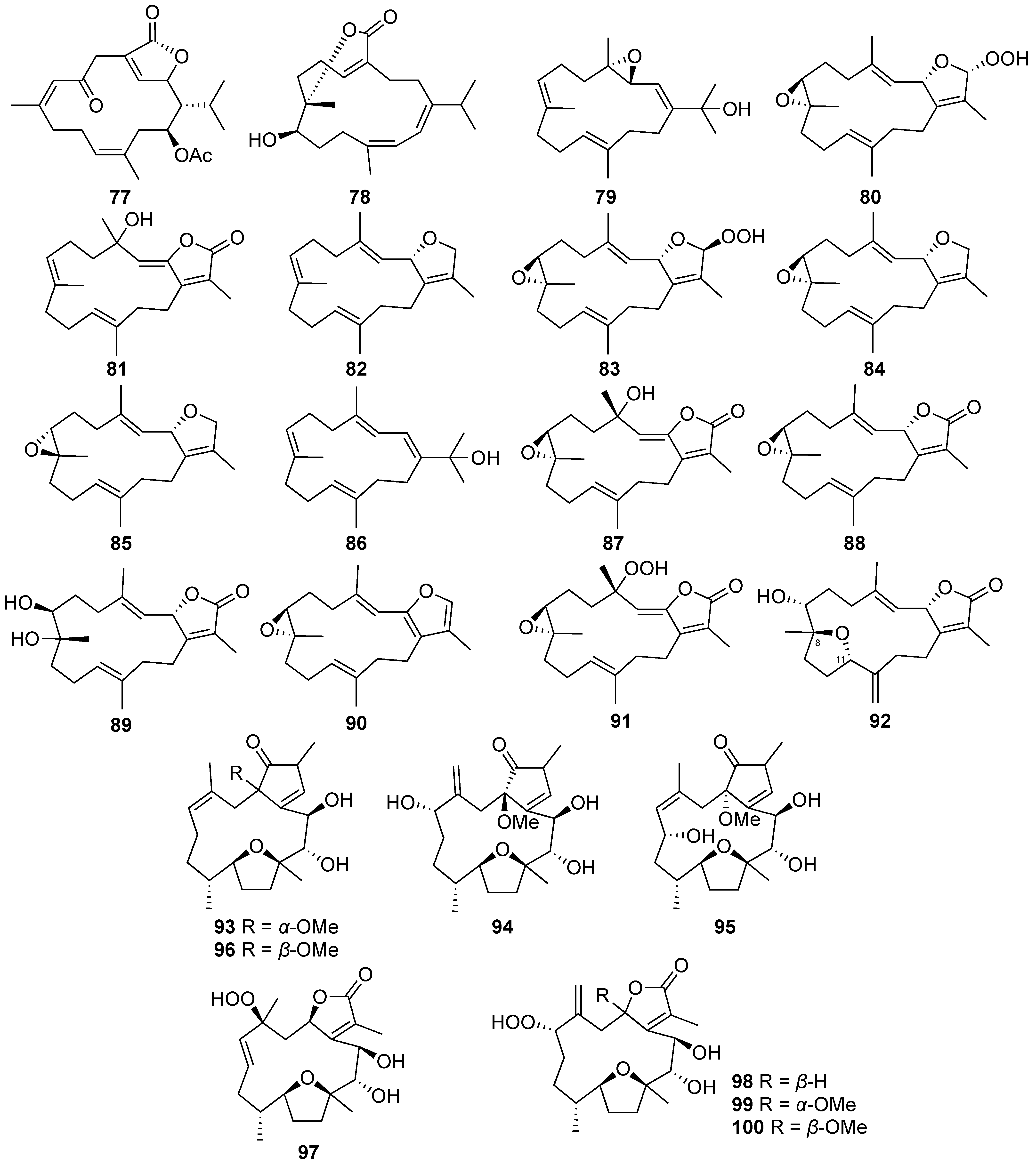
| Isosarcophytonolide D (77) | Cembrane-type diterpenoid | Known | Sarcophyton digitatum Sarcophyton glaucum | Exhibited cytotoxicity against MCF-7 carcinoma cell lines (IC50 = 10.9 ± 4.3 μg/mL) [8] Inhibited LPS-induced IL-1β production to 56 ± 1% at 10 µg/mL (IC50 = 14.9 ± 5.1 µg/mL) [8] Exhibited cytotoxicity against HL-60, CCRF-CEM, and MOLT-4 cell lines (ED50 = 13.0 ± 1.9, 15.3 ± 2.5, and 17.2 ± 3.1 μg/mL, respectively) [38] |
| Sarcotenusene A (79) | Cembrane-type diterpenoid | New | Sarcophyton tenuispiculatum | Exhibited cytotoxicity against MCF-7 cells (IC50 = 34.3 ± 3.7 μM) [9] |
| Sarcophytonin F (83) | Cembrane-type diterpenoid | Known | Sarcophyton tenuispiculatum | Exhibited cytotoxicity against MCF-7 and MDA-MB-231 cells (IC50 = 30.1 ± 3.1 and 38.6 ± 5.0 μM, respectively) [9] |
| (2S,7S,8S)-Sarcophytoxide (84) | Cembrane-type diterpenoid | Known | Sarcophyton tenuispiculatum | Exhibited cytotoxicity against MCF-7 and HepG2 cells (IC50 = 37.6 ± 4.2 and 35.2 ± 4.4 μM, respectively) [9] |
| (2S,7R,8R)-Sarcophytoxide (85) | Cembrane-type diterpenoid | Known | Sarcophyton tenuispiculatum | Exhibited cytotoxicity against MCF-7 and HepG2 cells (IC50 = 33.3 ± 3.5 and 28.6 ± 3.4 μM, respectively) [9] |
| 3,4-Dihydro-4α-hydroxy-∆2-sarcophine (87) | Cembrane-type diterpenoid | Known | Sarcophyton tenuispiculatum | Exhibited cytotoxicity against MCF-7 and HepG2 cells (IC50 = 24.3 ± 3.0 and 34.5 ± 4.2 μM, respectively) [9] |
| (+)-7α,8β-Dihydroxydeepoxysarcophine (89) | Cembrane-type diterpenoid | Known | Sarcophyton tenuispiculatum | Inhibited IL-1β production to 56 ± 1% in LPS-stimulated murine macrophage J774A.1 cell at 30 µM [9] |
| A hydroperoxide obtained by autoxidation of dihydrofuranocembranoid (91) | Cembrane-type diterpenoid | Known | Sarcophyton tenuispiculatum | Exhibited cytotoxicity against MCF-7 and HepG2 cells (IC50 = 27.2 ± 4.0 and 36.4 ± 5.3 μM, respectively) [9] |
| Briaviotriol A (94) | Cembrane-type diterpenoid | New | Briareum violaceum | Reduced the levels of iNOS to 67.7 ± 2.4% at 10 μM [40] |
| Briaviotriol B (95) | Cembrane-type diterpenoid | New | Briareum violaceum | Reduced the levels of iNOS to 79.5 ± 9.4% at 10 μM [40] |
| Briaviodiol A (96) | Cembrane-type diterpenoid | Known | Briareum violaceum | Reduced the levels of iNOS to 61.9 ± 7.3% at 10 μM [40] |
| Briaviodiol B (97) | Cembrane-type diterpenoid | New | Briareum violaceum | Reduced the level of iNOS to 43 ± 6% at 10 μM [41] |
| Briaviodiol D (99) | Cembrane-type diterpenoid | New | Briareum violaceum | Reduced the level of iNOS to 61 ± 7% at 10 μM [41] |
| Briaviodiol E (100) | Cembrane-type diterpenoid | New | Briareum violaceum | Reduced the level of iNOS to 46 ± 10% at 10 μM [41] |
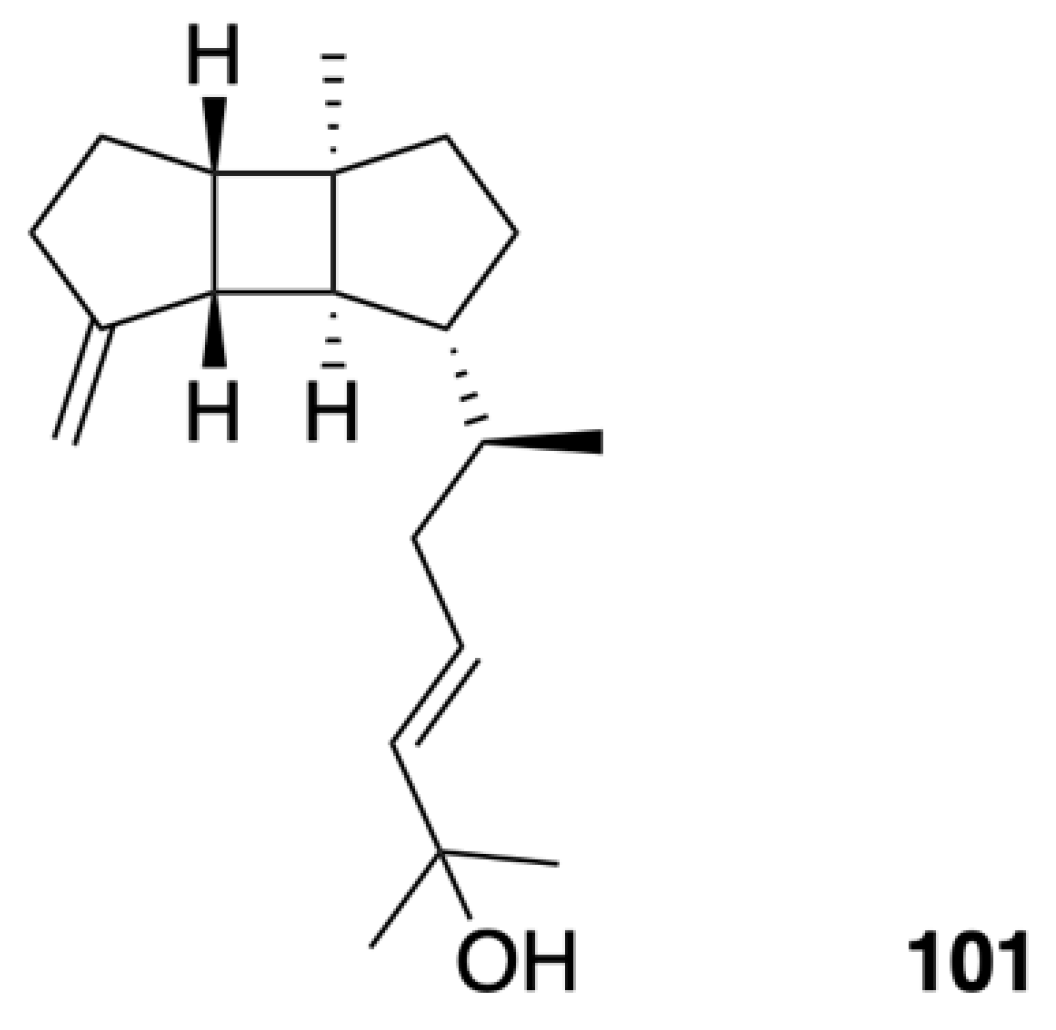
| Leptoclalin A (101) | Spatane-type diterpenoid | New | Sinularia leptoclados | Exhibited weak cytotoxicity against human tumor cell lines T-47D (IC50 = 15.4 μg/mL) and K-562 (IC50 = 12.8 μg/mL) [43] |
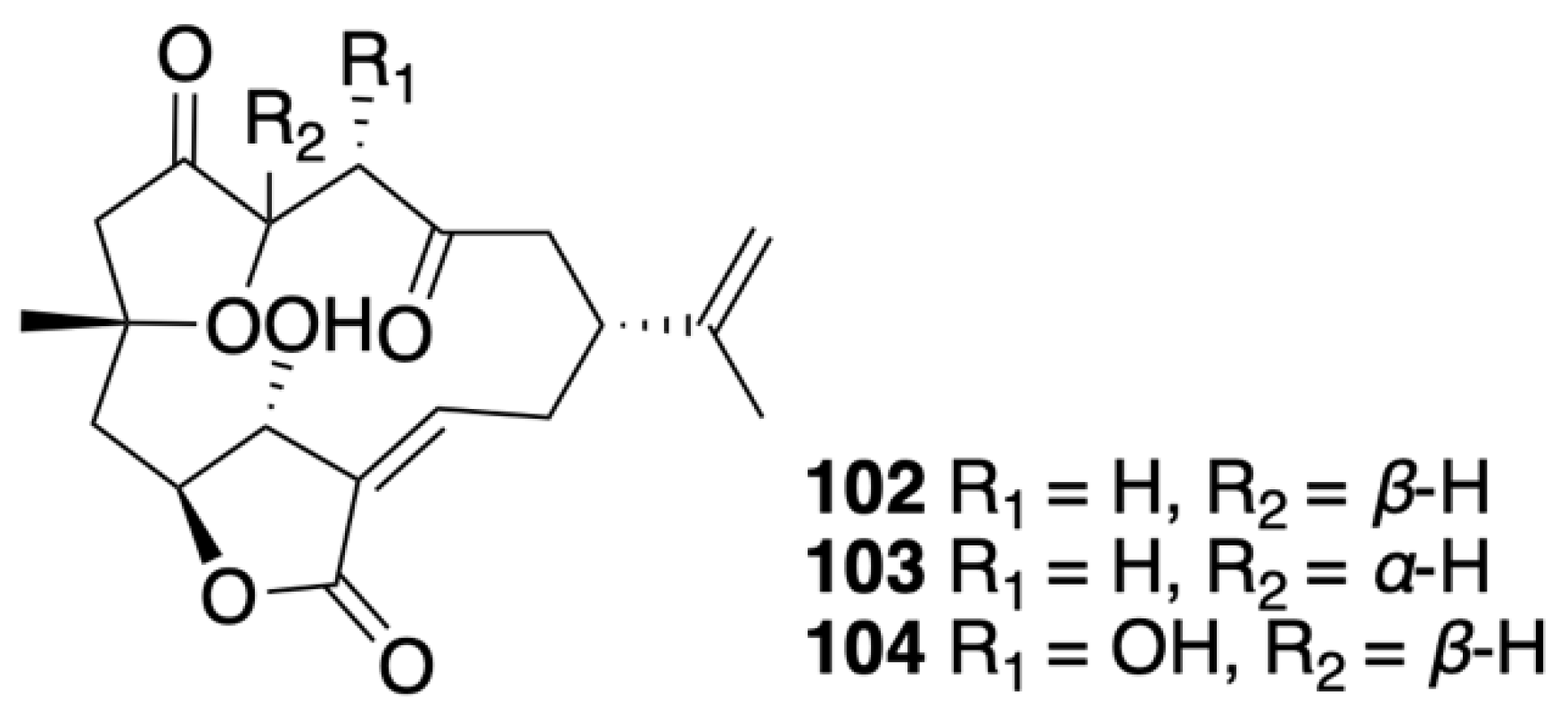
| 5-epi-Sinuleptolide (102) | Norcembranoidal diterpene | Known | Sinularia numerosa | Exhibited cytotoxicity against CCRF-CEM and HL-60 cells (IC50 = 11.07 and 11.11 μg/mL, respectively) [45] |
| 4α-Hydroxy-5-epi-sinuleptolide (104) | Norcembranoidal diterpene | New | Sinularia numerosa | Exhibited cytotoxicity against CCRF-CEM, HL-60, K-562, and U-937 cells (IC50 = 4.21, 10.38, 18.07, and 10.08 μg/mL, respectively) [45] |
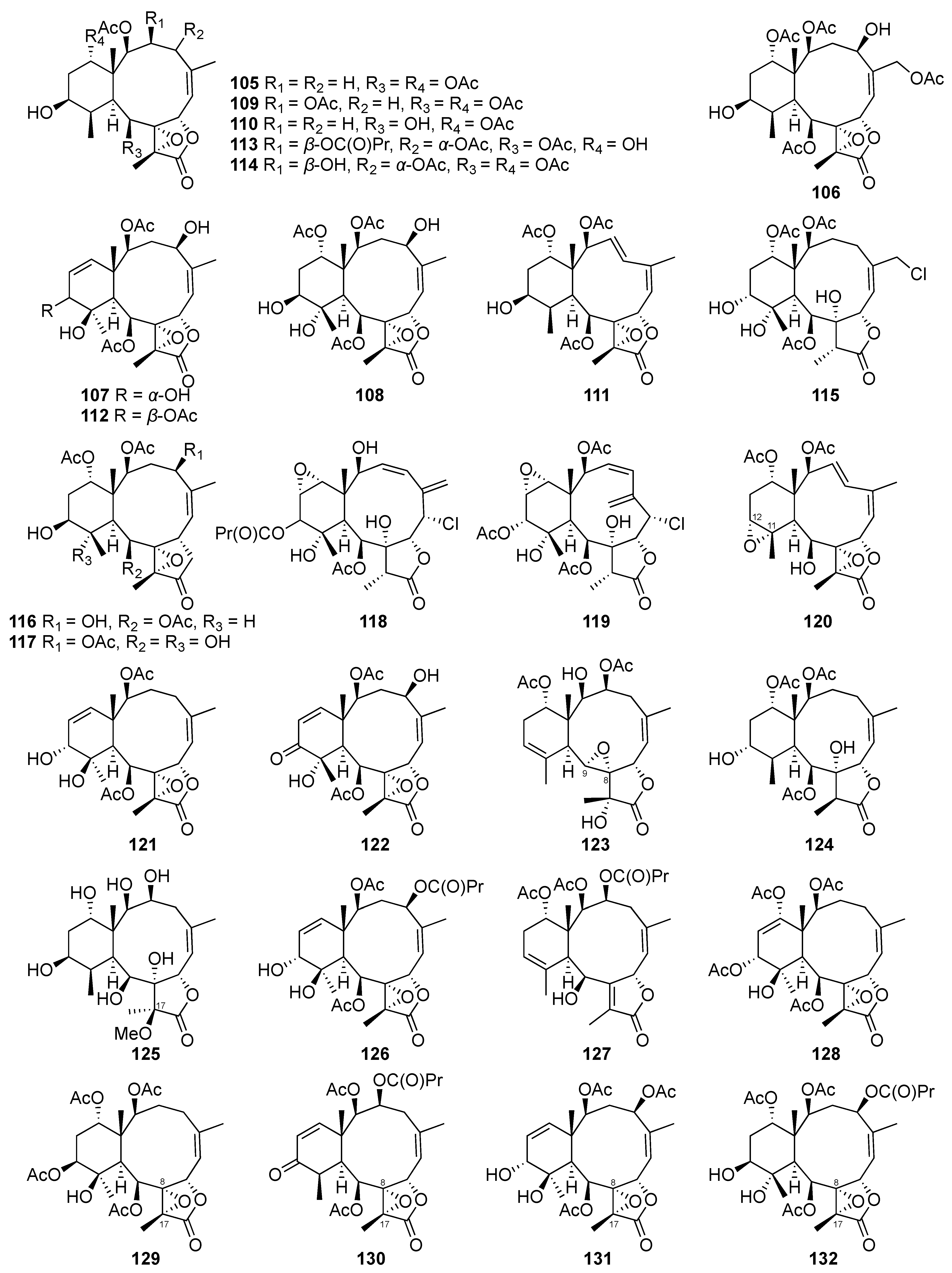
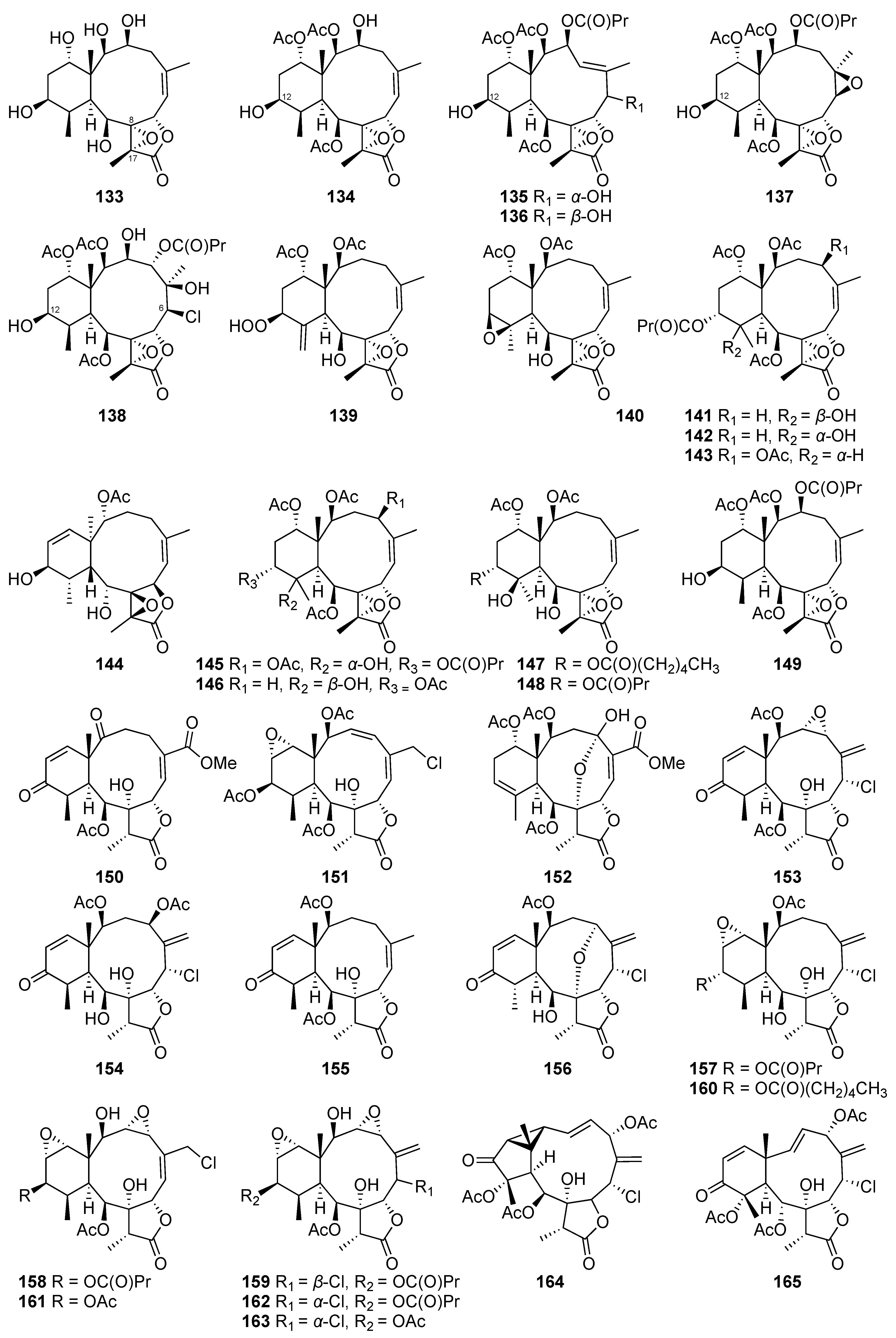
| Excavatolide C (109) | Briarane-type diterpene | Known | Briareum stechei | Showed a 15.47% inhibitory effect on superoxide anion generation by human neutrophils at 10 μg/mL [48] |
| Briaexcavatin P (114) | Briarane-type diterpene | New | Briareum stechei | Showed a 14.99% inhibitory effect on superoxide anion generation by human neutrophils at 10 μg/mL [49] |
| Briaexcavatin S (117) | Briarane-type diterpene | New | Briareum stechei | Exhibited weak cytotoxicity toward CCRF-CEM cells (ED50 = 37.8 μg/mL) [50] |
| Briaexcavatin V (120) | Briarane-type diterpene | New | Briareum stechei | Showed 11.39 ± 1.26% and 23.27 ± 8.65% inhibitory effects on superoxide anion generation and elastase release by human neutrophils at 10 μg/mL, respectively [52] |
| Briaexcavatin X (122) | Briarane-type diterpene | New | Briareum stechei | Displayed a 13.69 ± 3.84% inhibitory effect on superoxide anion generation by human neutrophils at 10 μg/mL [52] |
| Briaexcavatin Y (123) | Briarane-type diterpene | New | Briareum stechei | Displayed a 17.47 ± 0.85% inhibitory effect on superoxide anion generation by human neutrophils at 10 μg/mL [52] |
| Excavatoid E (127) | Briarane-type diterpene | New | Briareum stechei | Exhibited 26.22 ± 0.50% and 12.95 ± 6.99% inhibitory effects on elastase release and superoxide anion generation by human neutrophils at 10 μg/mL, respectively [54] |
| Excavatoid F (128) | Briarane-type diterpene | New | Briareum stechei | Exhibited a 30.63 ± 4.68% inhibitory effect on elastase release by human neutrophils at 10 μg/mL [54] |
| Excavatoid H (130) | Briarane-type diterpene | New | Briareum stechei | Exhibited cytotoxicity against CCRF-CEM cells (ED50 = 13.1 μg/mL) [55] |
| Excavatoid I (131) | Briarane-type diterpene | New | Briareum stechei | Displayed 38.3% and 21.8% inhibitory effects on elastase release and superoxide anion generation by human neutrophils at 10 μg/mL, respectively [55] |
| Excavatoid L (134) | Briarane-type diterpene | New | Briareum stechei | Displayed 42.44 ± 2.38% and 31.25 ± 0.07% inhibitory effects on superoxide anion generation and elastase release by human neutrophils at 10 μg/mL, respectively [56] |
| Excavatoid M (135) | Briarane-type diterpene | New | Briareum stechei | Displayed 14.85 ± 3.66% and 16.96 ± 2.93% inhibitory effects on superoxide anion generation and elastase release by human neutrophils at 10 μg/mL, respectively [56] |
| Excavatoid N (136) | Briarane-type diterpene | New | Briareum stechei | Displayed 10.90 ± 0.50% and 22.21 ± 3.34% inhibitory effects on superoxide anion generation and elastase release by human neutrophils at 10 μg/mL, respectively [56] |
| Excavatoid O (137) | Briarane-type diterpene | New | Briareum stechei | Displayed a 16.9% inhibitory effect on elastase release by human neutrophils at 10 μg/mL [57] |
| Excavatoid P (138) | Briarane-type diterpene | New | Briareum stechei | Displayed a 16.1% inhibitory effect on elastase release by human neutrophils at 10 μg/mL [57] |
| Briarenolide D (139) | Briarane-type diterpene | New | Briareum stechei | Showed moderate cytotoxicity toward DLD-1 and CCRF-CEM cells (ED50 = 9.6 and 6.9 μg/mL, respectively) [47] |
| 2β-Acetoxy-2-(debutyryloxy)stecholide E (140) | Briarane-type diterpene | Known | Briareum stechei | Showed cytotoxicity against P-388 and HT-29 cell lines (ED50 = 0.61 and 6.96 μg/mL, respectively) [78] |
| Briaviolide L (142) | Briarane-type diterpene | New | Briareum violaceum | Reduced the levels of iNOS and COX-2 to 46.68% and 61.81%, respectively, at 33.7 μM [58] |
| Briaviolide O (145) | Briarane-type diterpene | New | Briareum violaceum | Reduced the levels of iNOS and COX-2 to 10.53 ± 1.38% and 84.31 ± 2.14%, respectively, at 10 μM [59] |
| Briaviolide P (146) | Briarane-type diterpene | New | Briareum violaceum | Reduced the level of COX-2 to 87.83 ± 3.36% at 10 μM [59] |
| Briaviolide Q (147) | Briarane-type diterpene | New | Briareum violaceum | Reduced the level of iNOS to 26.4 ± 1.5% at 10 μM [60] |
| Excavatolide Z (148) | Briarane-type diterpene | Known | Briareum violaceum | Reduced the level of iNOS to 66.2 ± 9.6% at 10 μM [60] |
| Excavatolide B (149) | Briarane-type diterpene | Known | Briareum stechei | Showed significant dose-dependent inhibition of iNOS gene expression in RAW 264.7 murine macrophages at doses of 1, 10, 25, and 50 μM [19] Significantly inhibited COX-2 gene expression in RAW 264.7 murine macrophages at 25 and 50 μM [61] Reduced the infiltration of inflammatory cells and iNOS protein expression to ameliorate the pain behavior and inflammatory response in carrageenan-induced inflammatory rats at 15 or 60 mg/kg [61] Anti-rheumatic effect: Reduced osteoclastogenesis via the downregulation of the inflammatory factors IL-17A and M-CSF to influence the MAPK and HO-1/HMGB-1 pathways in AIA and CIA rats [62] |
| Briarenol P (151) | Briarane-type diterpene | New | Briareum stechei | Reduced the expressions of iNOS and β-actin to 88.24 ± 7.51% and 85.40 ± 5.35% at 10 μM [46] |
| Briarenol S (154) | Briarane-type diterpene | New | Briareum stechei | Inhibited the release of iNOS to 78.50% [63] |
| Briarenol T (155) | Briarane-type diterpene | New | Briareum stechei | Inhibited the release of iNOS to 79.95% [63] |
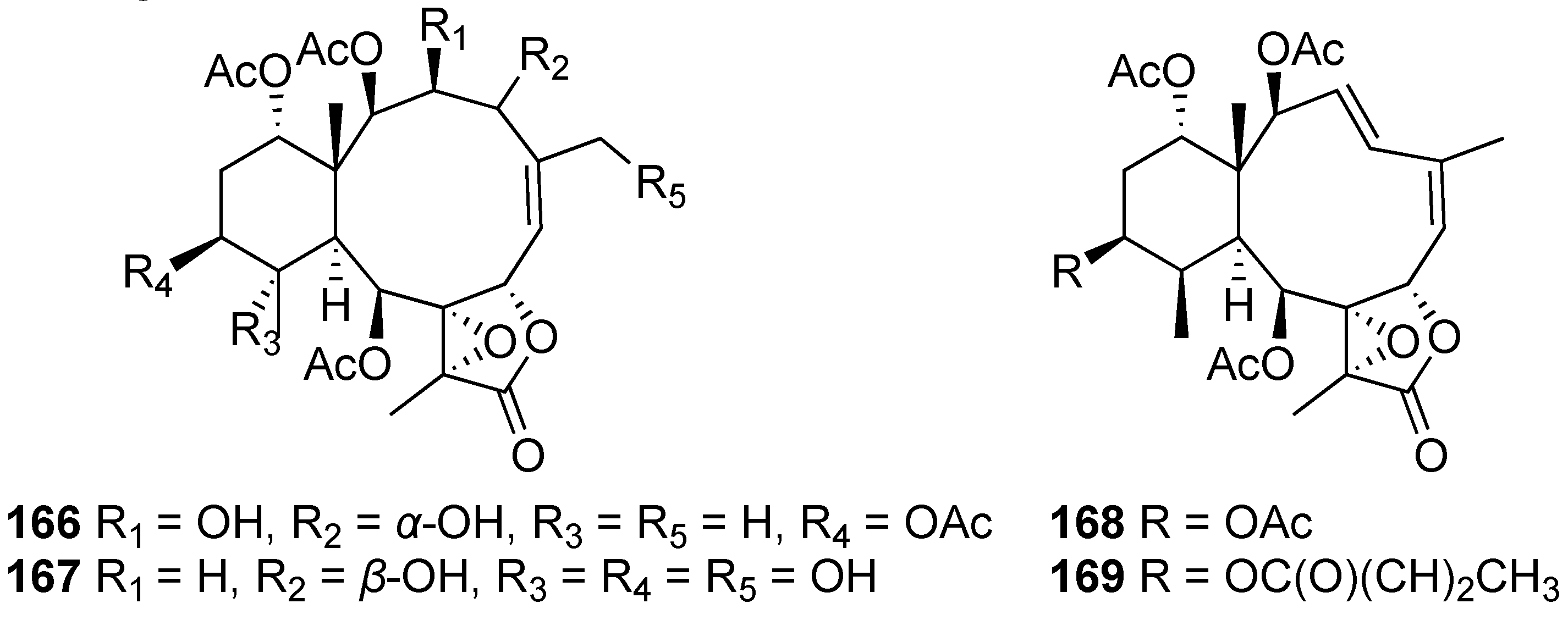
| Briavioid A (166) | Briarane-type diterpene | New | Briareum violaceum | Reduced the release of iNOS protein to 77.50% at 10 μM [65] |
| Briavioid B (167) | Briarane-type diterpene | New | Briareum violaceum | Reduced the release of iNOS protein to 80.24% at 10 μM [65] |
| Briavioid C (168) | Briarane-type diterpene | New | Briareum violaceum | Reduced the release of iNOS protein to 71.30% at 10 μM [65] |
| Excavatolide F (169) | Briarane-type diterpene | Known | Briareum violaceum | Significantly reduced the release of iNOS protein to 28.60% at 10 μM [65] |
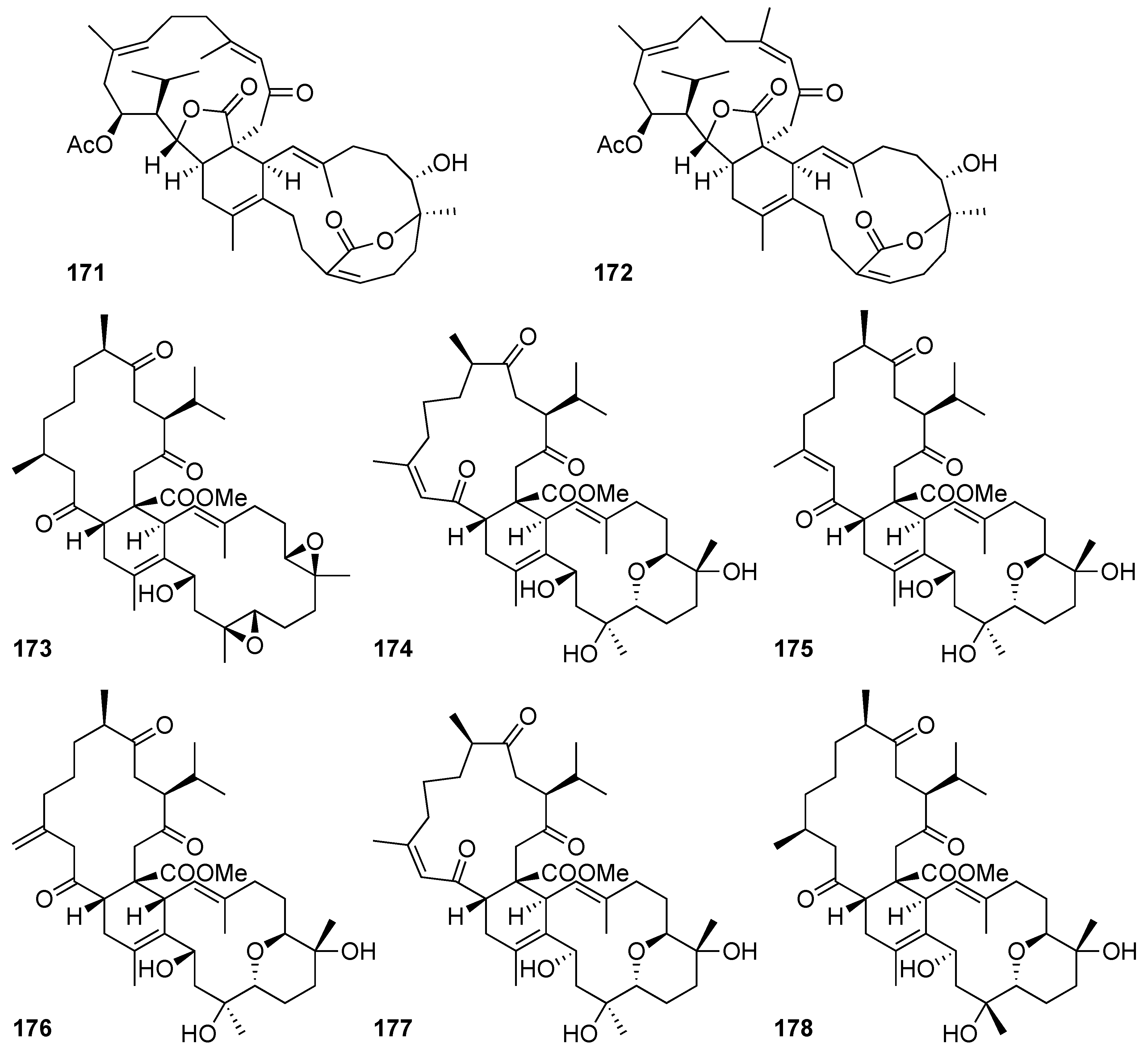
| Glaucumolide A (171) | Biscembranoid | New | Sarcophyton digitatum Sarcophyton glaucum | Exhibited cytotoxicity against MCF-7, HepG2, and HeLa carcinoma cell lines (IC50 = 10.1 ± 3.3, 14.9 ± 3.5, and 17.1 ± 4.5 μg/mL, respectively) [8] Inhibited LPS-induced IL-1β production to 68 ± 1% at 10 µg/mL (IC50 = 10.7 ± 2.7 µg/mL) [8] Exhibited cytotoxicity against HL-60, CCRF-CEM, MOLT-4, and K-562 cell lines (ED50 = 6.6 ± 1.2, 7.4 ± 1.5, 11.0 ± 2.8, and 19.2 ± 2.3 μg/mL, respectively) [38] Inhibited superoxide anion generation and elastase release in human neutrophils (IC50 = 2.79 ± 0.66 and 3.97± 0.10 µM, respectively) [38] Significantly reduced the levels of iNOS and COX-2 to 59.4 ± 9.0 and 66.5 ± 4.4%; 31.3 ± 6.5 and 78.3 ± 5.0%; and −2.6 ± 2.7 and −0.5 ± 3.2% at concentrations of 5, 10, and 20 μM, respectively [38] |
| Glaucumolide B (172) | Biscembranoid | New | Sarcophyton digitatum Sarcophyton glaucum | Exhibited cytotoxicity against MCF-7, MDA-MB-231, and HepG2 carcinoma cell lines (IC50 = 9.4 ± 3.0, 17.8 ± 4.5, and 14.9 ± 4.2 μg/mL, respectively) [8] Exhibited cytotoxicity against HL-60, CCRF-CEM, MOLT-4, and K-562 cell lines (ED50 = 3.8 ± 0.9, 5.3 ± 1.4, 11.0 ± 2.2, and 12.6 ± 0.7 μg/mL, respectively) [38] Inhibited superoxide anion generation and elastase release in human neutrophils (IC50 = 2.79 ± 0.32 and 3.97± 0.10 µM, respectively) [38] Significantly reduced the levels of iNOS and COX-2 to 75.9 ± 3.5 and 64.3 ± 6.9%; and 43.4 ± 5.0 and 6.0 ± 3.6% at concentrations of 10 and 20 μM, respectively [38] |
| Ximaolide A (173) | Biscembranoid | Known | Sarcophyton glaucum | Reduced the level of COX-2 expression to 22.0 ± 6.5% in LPS-treated macrophage cells at 20 μM [38] |
| Sardigitolide B (175) | Biscembranoid | New | Sarcophyton digitatum | Exhibited cytotoxicity against MCF-7 and MDA-MB-231 carcinoma cell lines (IC50 = 9.6 ± 3.0 and 14.8 ± 4.0 μg/mL, respectively) [8] |
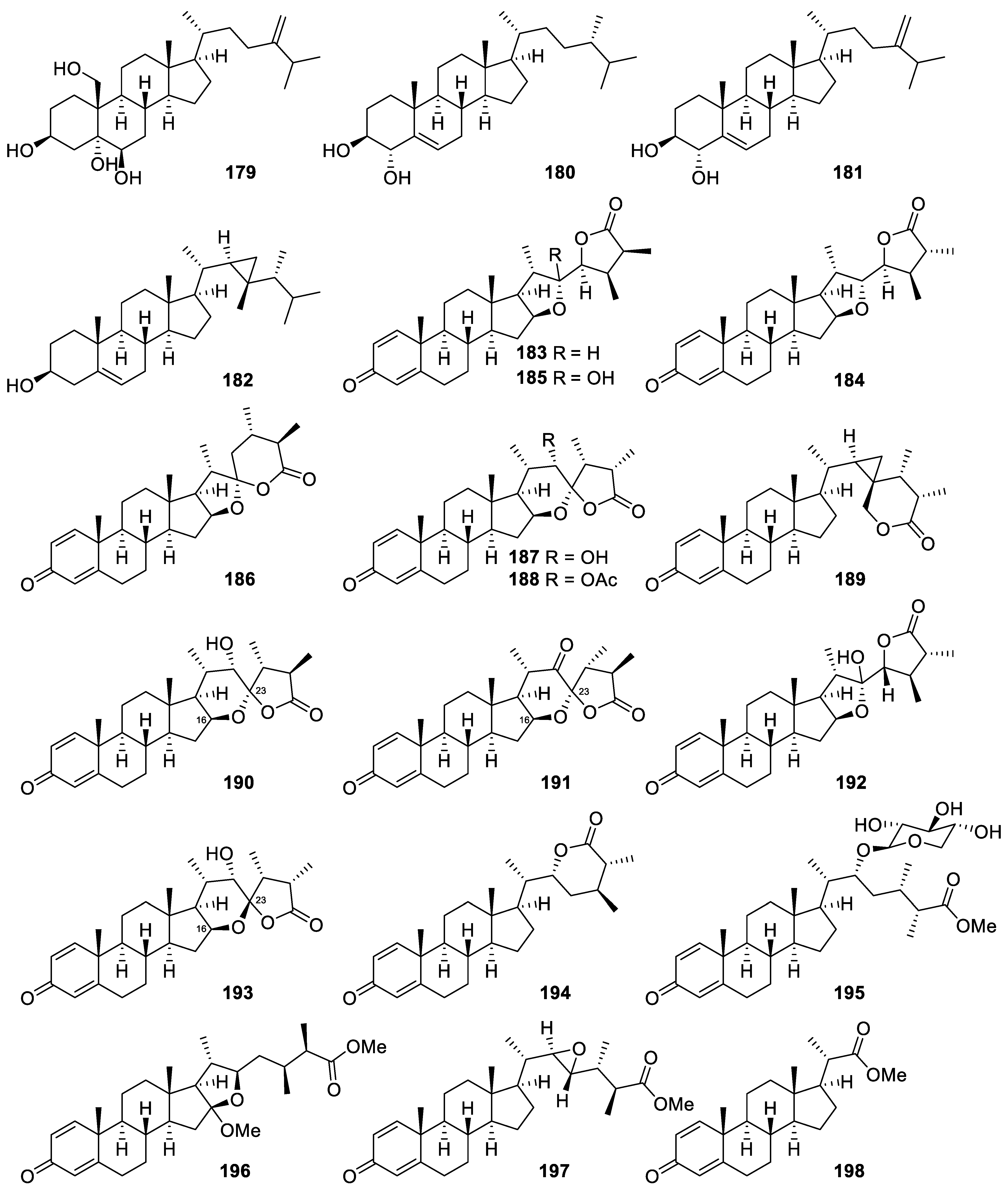
| Nephalsterol A (179) | Sterol | Known | Litophyton columnaris | Exhibited cytotoxicity against MOLT-4, SUP-T1, U-937, DLD-1, LNCaP, and MCF7 cells (IC50 = 22.5, 32.4, 38.6, 44.2, 11.6, and 9.8 μM, respectively) [36] |
| (24S)-24-Methylcholest-5-en-3β,4α-diol (180) | Sterol | New | Sinularia sandensis | Reduced the release of iNOS to 89.52 ± 3.37% at 10 μM [67] |
| Gorgosterol (182) | Sterol | Known | Sinularia sandensis | Reduced the release of iNOS to 87.34 ± 2.48% at 10 μM [67] |
| Sinubrasolide A (183) | Withanolidal steroid | New | Sinularia brassica | Exhibited cytotoxicity against P388, MOLT-4, K-562, and HT-29 cells (IC50 = 29.9 ± 3.0, 12.1 ± 1.1, 8.7 ± 1.4, and 18.7 ± 2.5 μM, respectively) [69] Showed potent inhibitory effect against superoxide anion generation and elastase release in fMLP/CB-stimulated cells (IC50 = 3.5 ± 0.9 and 1.4 ± 0.1 μM, respectively) [69] |
| Sinubrasolide B (184) | Withanolidal steroid | New | Sinularia brassica | Exhibited cytotoxicity against P388, MOLT 4, and HT-29 cancer cell lines (ED50 = 9.1 ± 1.4, 4.8 ± 0.9, and 4.8 ± 0.7 μM, respectively) [68] |
| Sinubrasolide E (187) | Withanolidal steroid | New | Sinularia brassica | Showed cytotoxicity against MOLT 4 and HT-29 cell lines (ED50 = of 9.9 ± 1.8 and 7.5 ± 1.5 μM, respectively) [68] |
| Sinubrasolide H (190) | Withanolidal steroid | New | Sinularia brassica | Exhibited cytotoxicity against P388, MOLT-4, K-562, and HT-29 cells (IC50 = 39.8 ± 7.7, 28.6 ± 5.9, 29.7 ± 8.6, and 24.4 ± 6.2 μM, respectively) [69] |
| Sinubrasolide J (192) | Withanolidal steroid | New | Sinularia brassica | Exhibited cytotoxicity against P388, MOLT-4, K-562, and HT-29 cells (IC50 = 18.7 ± 3.1, 17.2 ± 1.5, 12.6 ± 3.1, and 11.2 ± 1.1 μM, respectively) [69] |
| Sinubrasolide K (193) | Withanolidal steroid | New | Sinularia brassica | Exhibited cytotoxicity against P388, MOLT-4, K-562, and HT-29 cells (IC50 = 18.3 ± 2.6, 13.7 ± 3.3, 17.4 ± 3.3, and 20.5 ± 3.7 μM, respectively) [69] |
| Sinubrasone B (196) | Non-withanolidal Steroid | New | Sinularia brassica | Showed significant cytotoxicity against P388D1, MOLT-4, K-562, and HT-29 cells (IC50 = 9.7 ± 1.2, 6.0 ± 0.4, 5.2 ± 0.8, and 7.6 ± 2.3 μM, respectively) [70] |
| Sinubrasone C (197) | Non-withanolidal Steroid | New | Sinularia brassica | Showed significant cytotoxicity against P388D1, MOLT-4, K-562, and HT-29 cells (IC50 = 5.7 ± 1.8, 5.3 ± 1.3, 12.1 ± 2.4, and 10.4 ± 2.2 μM, respectively) [70] Inhibited elastase release in fMLP/CB-induced human neutrophils (IC50 = 6.6 ± 1.1 μM) [70] |
| Sinubrasone D (198) | Non-withanolidal Steroid | New | Sinularia brassica | Inhibited superoxide anion generation and elastase release in fMLP/CB-induced human neutrophils (IC50 = 8.4 ± 1.1 and 6.5 ± 1.1 μM) [70] |
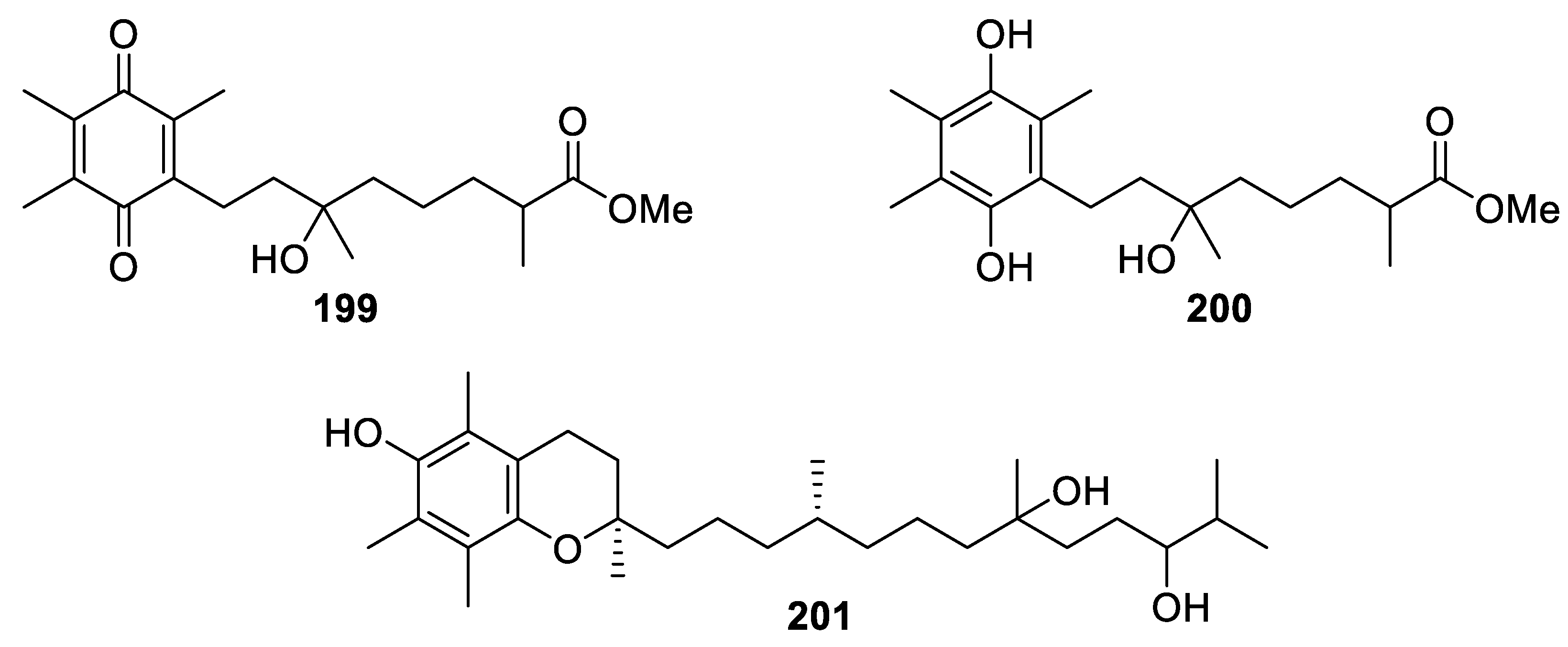
| Flexibilisquinone (199) | Quinone derivative | New | Sinularia flexibilis | Significantly suppressed the levels of iNOS and COX-2 at 5–20 µM and 20 µM, respectively [71] |
| Sarcotenuhydroquinone (200) | 1,4-dihydrobenzoquinone | New | Sarcophyton tenuispiculatum | Exhibited cytotoxicity toward MCF-7 and MDA-MB-231 cells (IC50 = 25.3 ± 2.8 and 36.4 ± 3.6 μM, respectively) [9] |
| Crassumtocopherol C (201) | α-tocopherol derivative | New | Lobophytum crassum | Exhibited cytotoxicity against K562 and Sup-T1 cells (IC50 = 34.0 and 23.3 μM, respectively) [7] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, N.B.A.; Chen, L.-Y.; El-Shazly, M.; Peng, B.-R.; Su, J.-H.; Wu, H.-C.; Lee, I.-T.; Lai, K.-H. Towards Sustainable Medicinal Resources through Marine Soft Coral Aquaculture: Insights into the Chemical Diversity and the Biological Potential. Mar. Drugs 2022, 20, 640. https://doi.org/10.3390/md20100640
Nguyen NBA, Chen L-Y, El-Shazly M, Peng B-R, Su J-H, Wu H-C, Lee I-T, Lai K-H. Towards Sustainable Medicinal Resources through Marine Soft Coral Aquaculture: Insights into the Chemical Diversity and the Biological Potential. Marine Drugs. 2022; 20(10):640. https://doi.org/10.3390/md20100640
Chicago/Turabian StyleNguyen, Ngoc Bao An, Lo-Yun Chen, Mohamed El-Shazly, Bo-Rong Peng, Jui-Hsin Su, Ho-Cheng Wu, I-Ta Lee, and Kuei-Hung Lai. 2022. "Towards Sustainable Medicinal Resources through Marine Soft Coral Aquaculture: Insights into the Chemical Diversity and the Biological Potential" Marine Drugs 20, no. 10: 640. https://doi.org/10.3390/md20100640
APA StyleNguyen, N. B. A., Chen, L. -Y., El-Shazly, M., Peng, B. -R., Su, J. -H., Wu, H. -C., Lee, I. -T., & Lai, K. -H. (2022). Towards Sustainable Medicinal Resources through Marine Soft Coral Aquaculture: Insights into the Chemical Diversity and the Biological Potential. Marine Drugs, 20(10), 640. https://doi.org/10.3390/md20100640