
The Inhibitory Effect of Phycocyanin Peptide on Pulmonary Fibrosis In Vitro




Abstract
:1. Introduction
2. Results
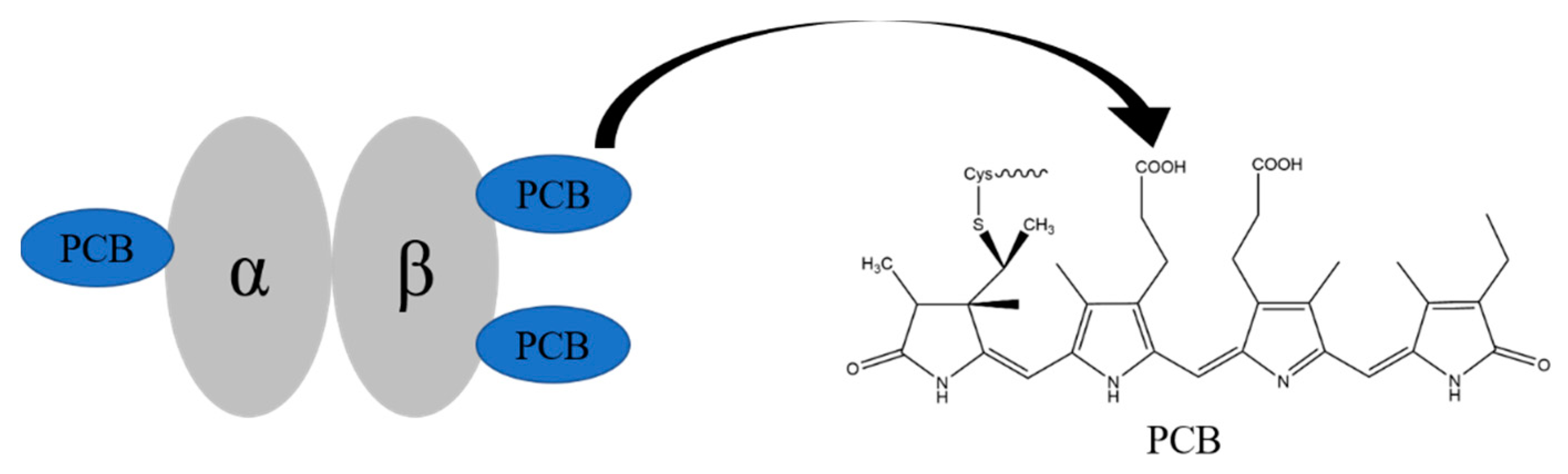
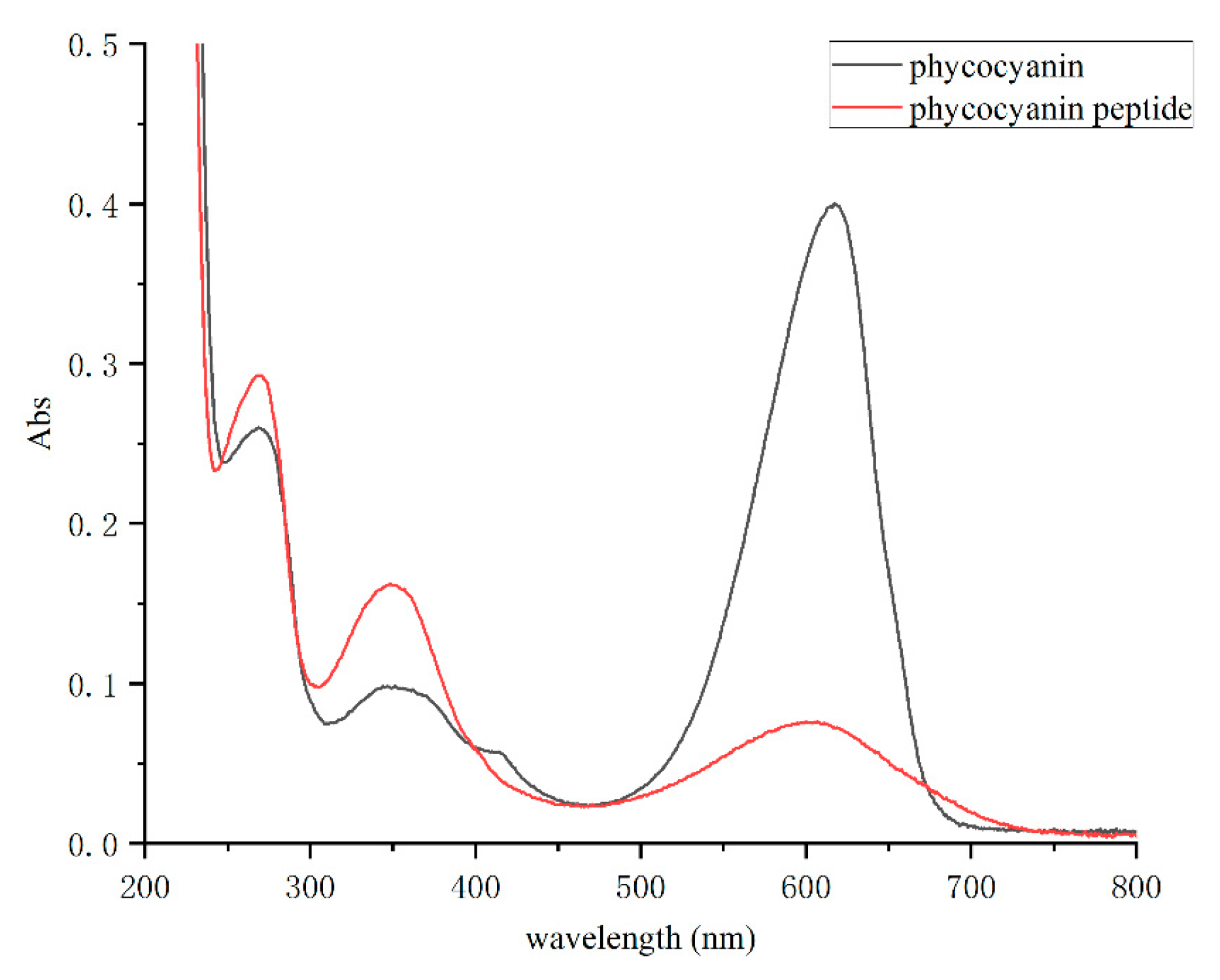
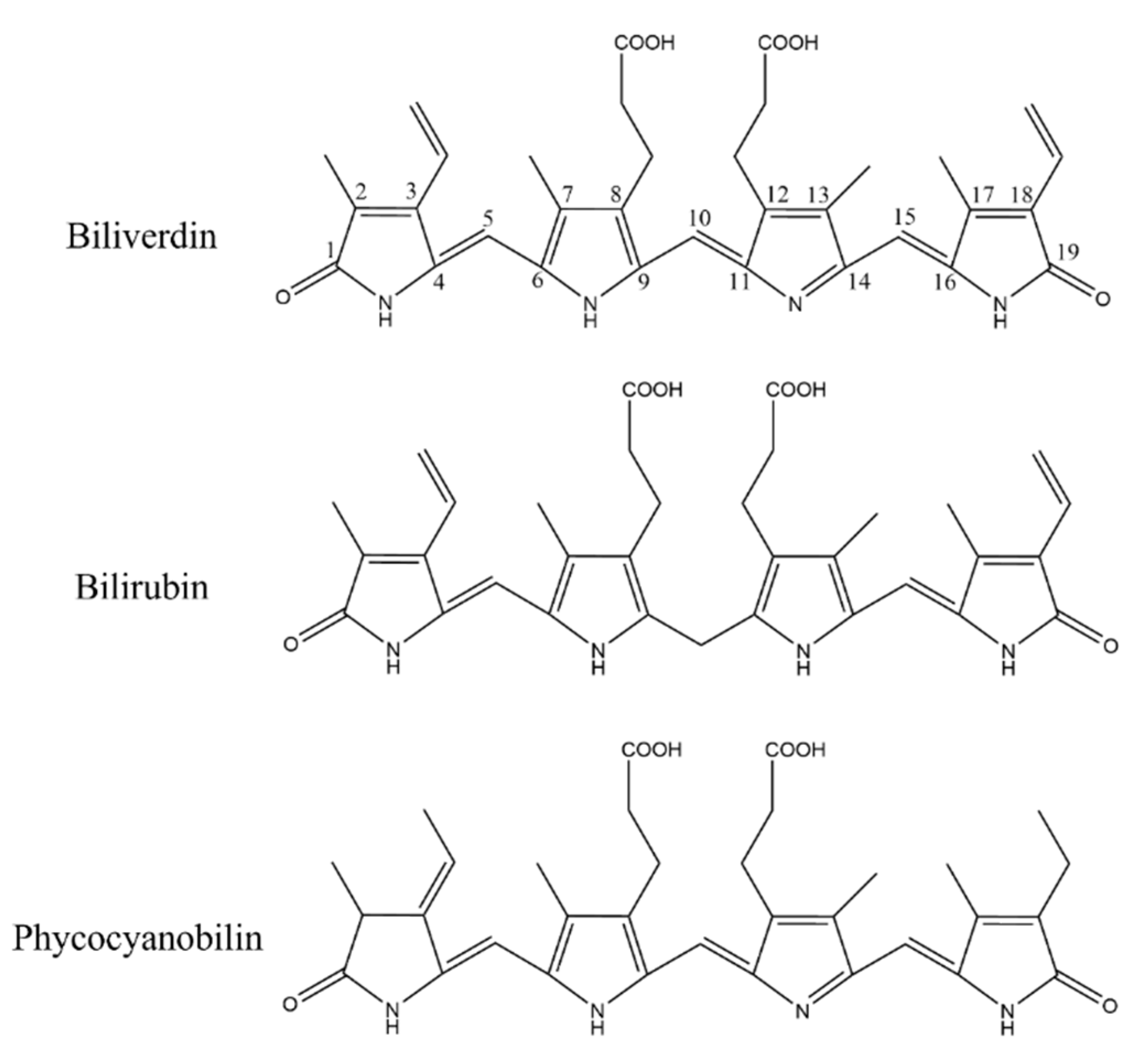
2.1. Enzymatic Hydrolysis of Phycocyanin and Its Absorption Spectrum and Molecular Weight Determination
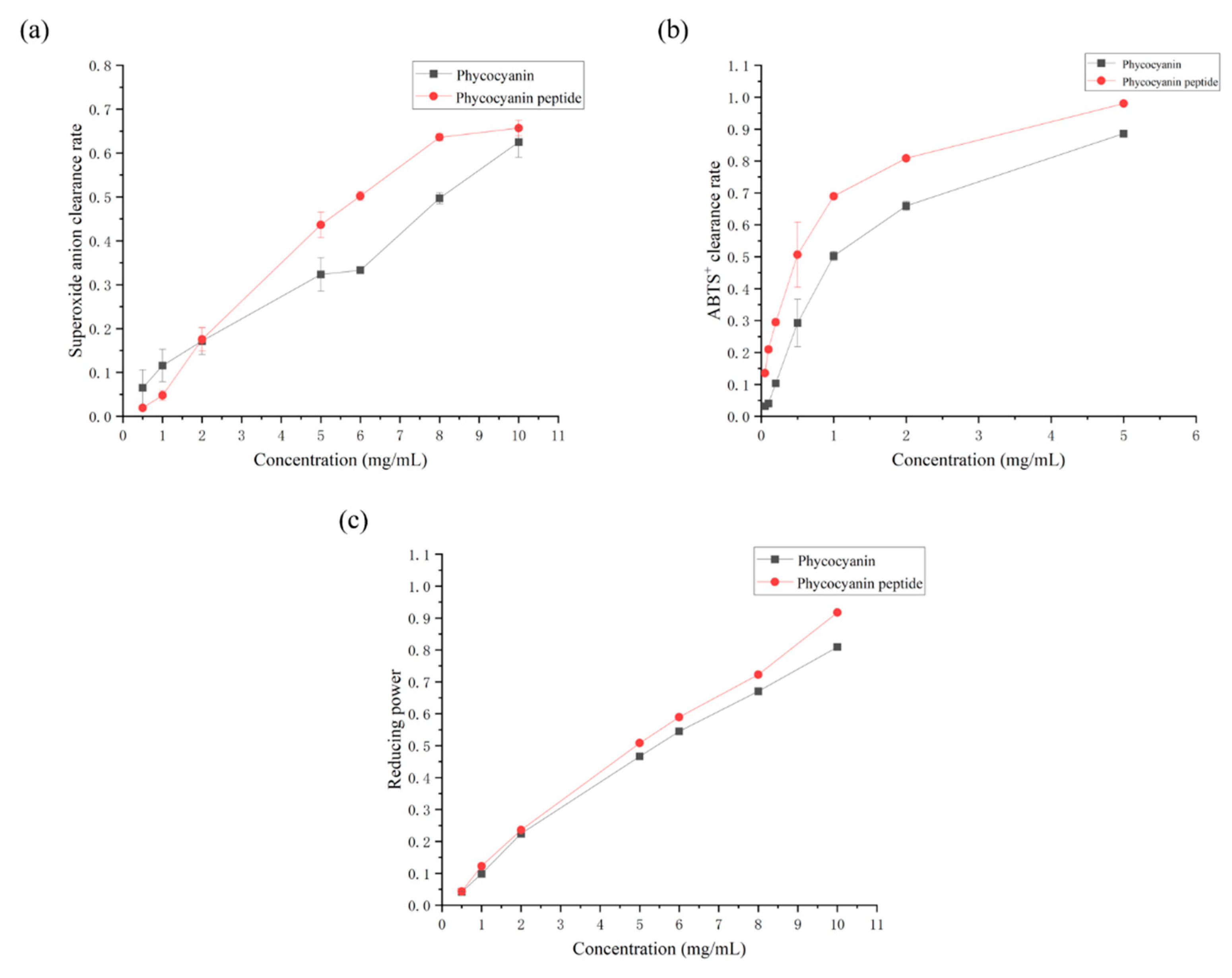
2.2. Antioxidant Activity of Phycocyanin Peptide
2.2.1. Superoxide Anion Scavenging Rate
2.2.2. ABTS+ Scavenging Rate
2.2.3. Total Reducing Power

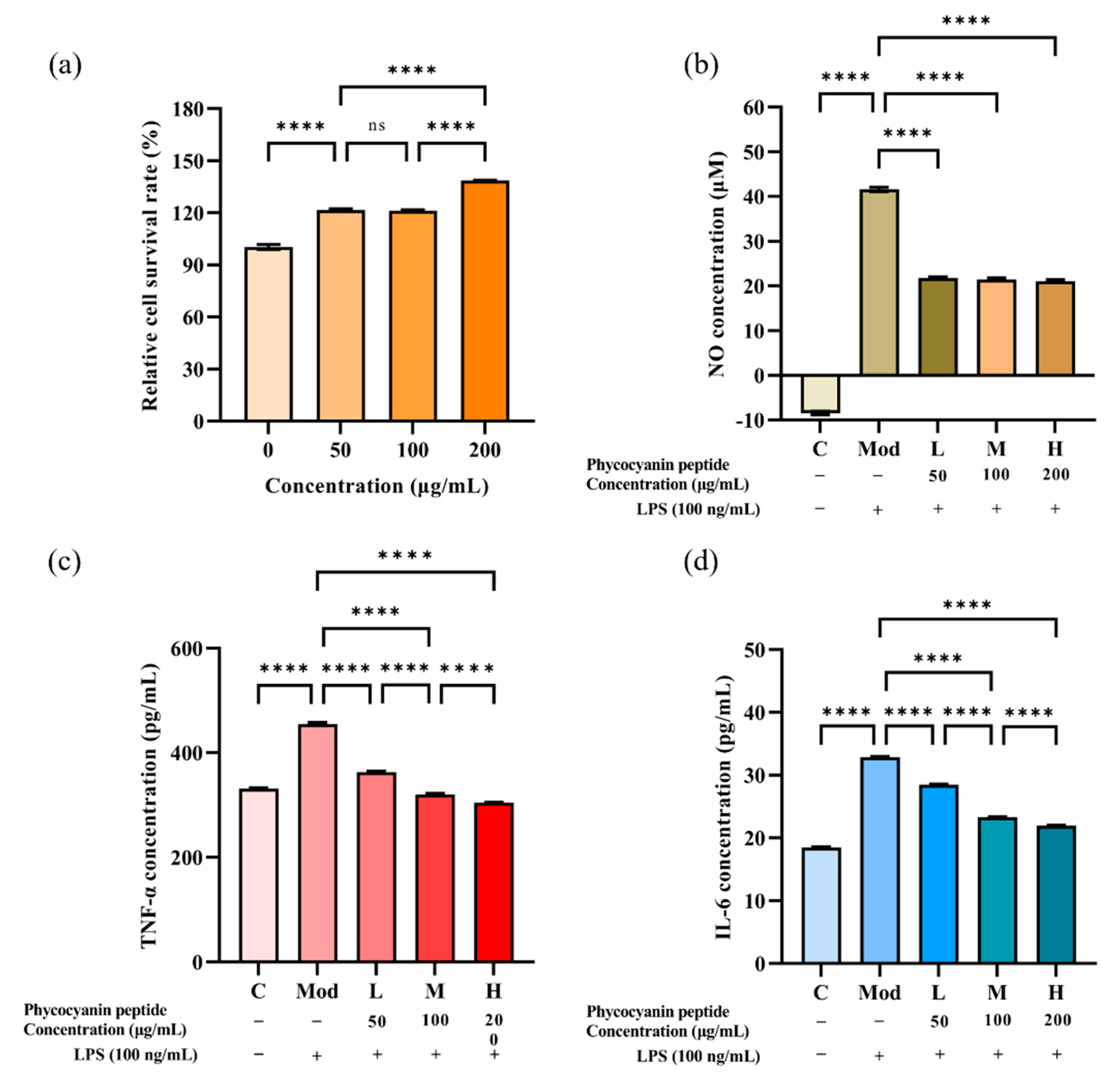
2.3. Anti-Inflammatory Activity of Phycocyanin Peptide
2.3.1. Effect of Phycocyanin Peptide on Survival Rate of RAW264.7 Cells
2.3.2. Inhibitory Effect of Phycocyanin Peptide on NO
2.3.3. Inhibitory Effect of Phycocyanin Peptide on TNF-α and IL-6

2.4. Anti-Pulmonary Fibrosis Activity of Phycocyanin Peptide
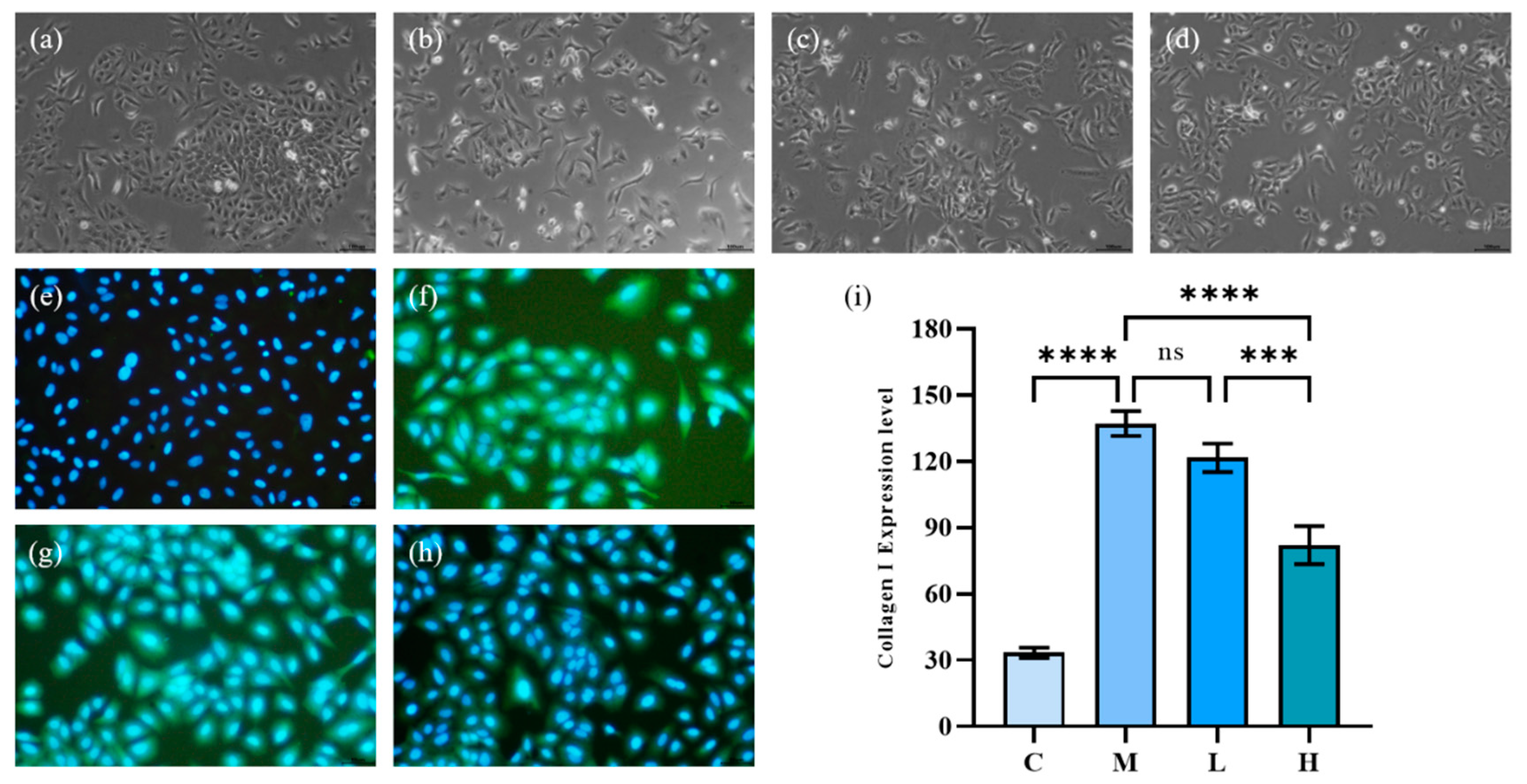
2.4.1. A549 Cell Morphology and Collagen I Expression
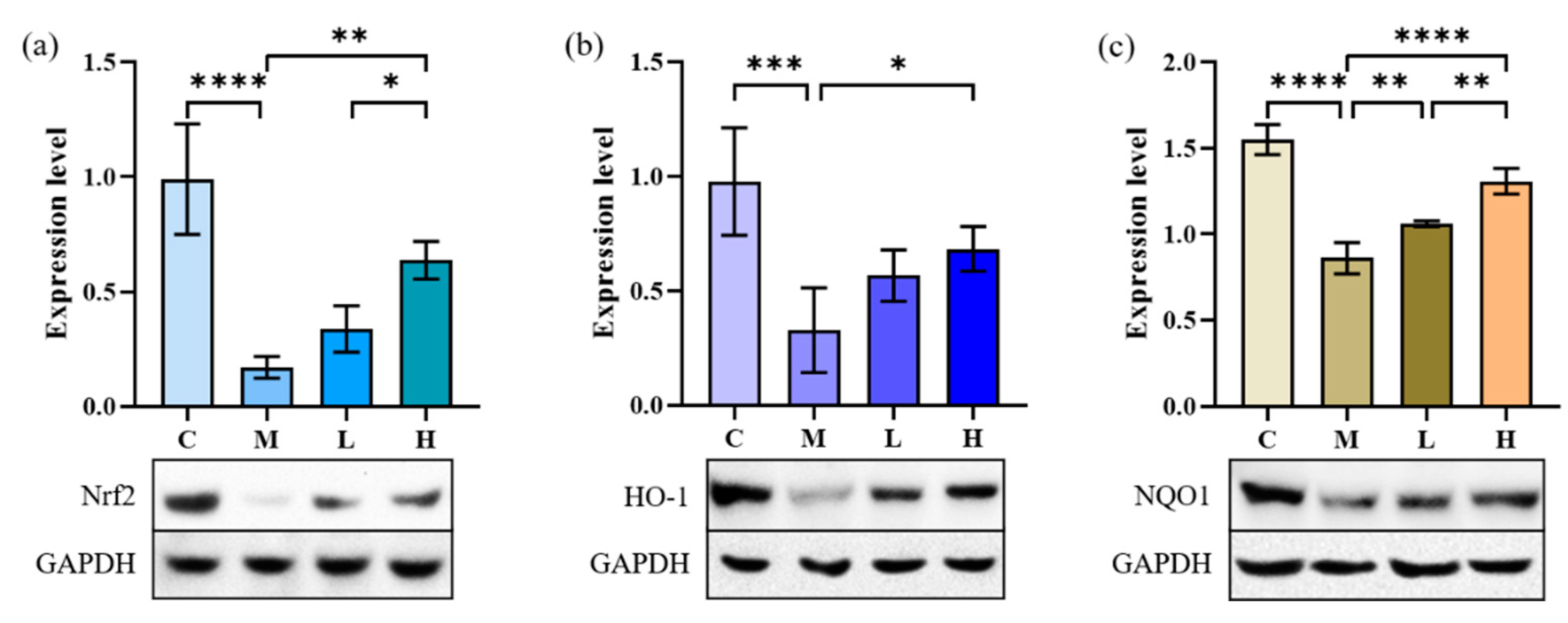
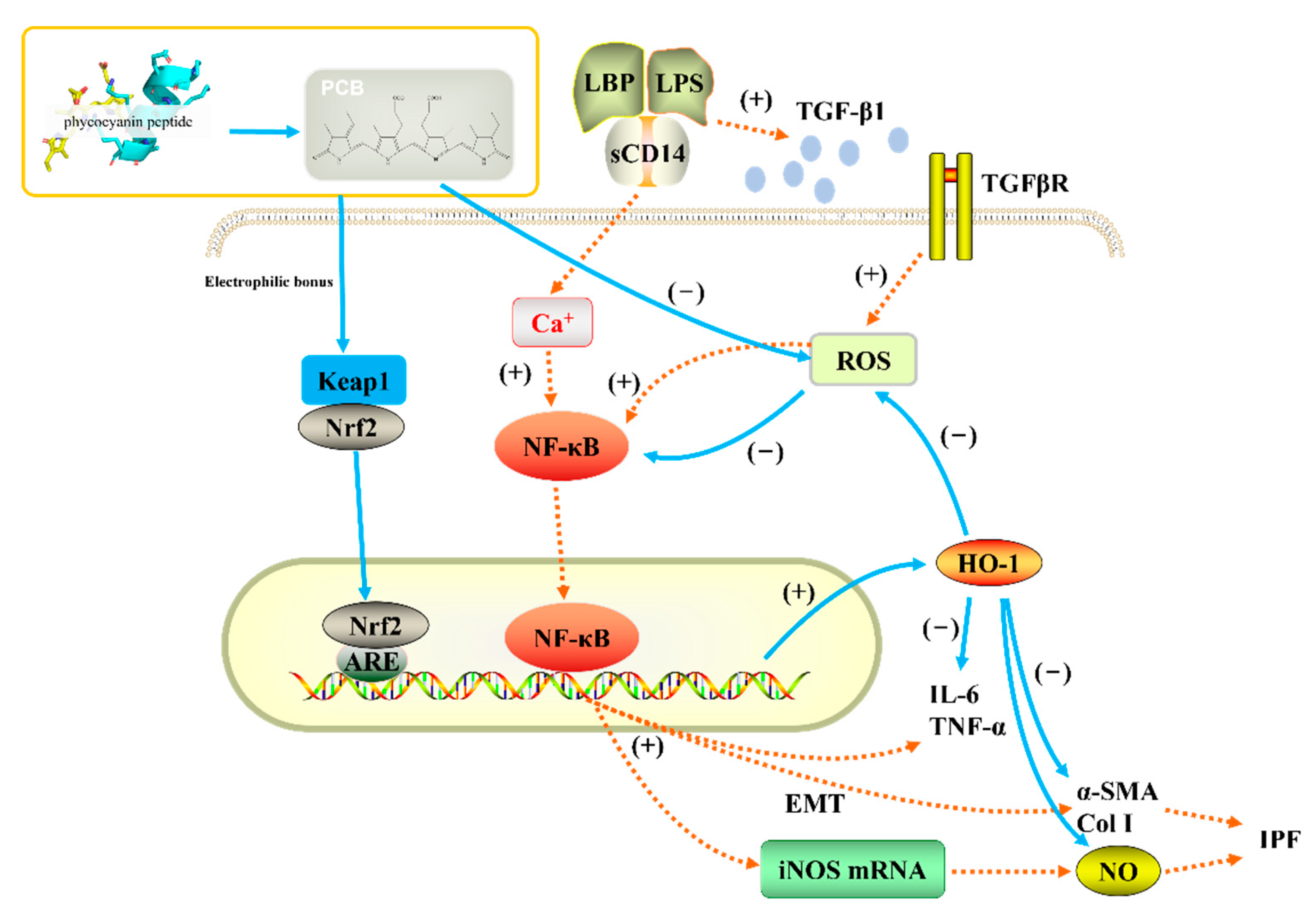
2.4.2. Nrf2, NQO1 and HO-1 Expression
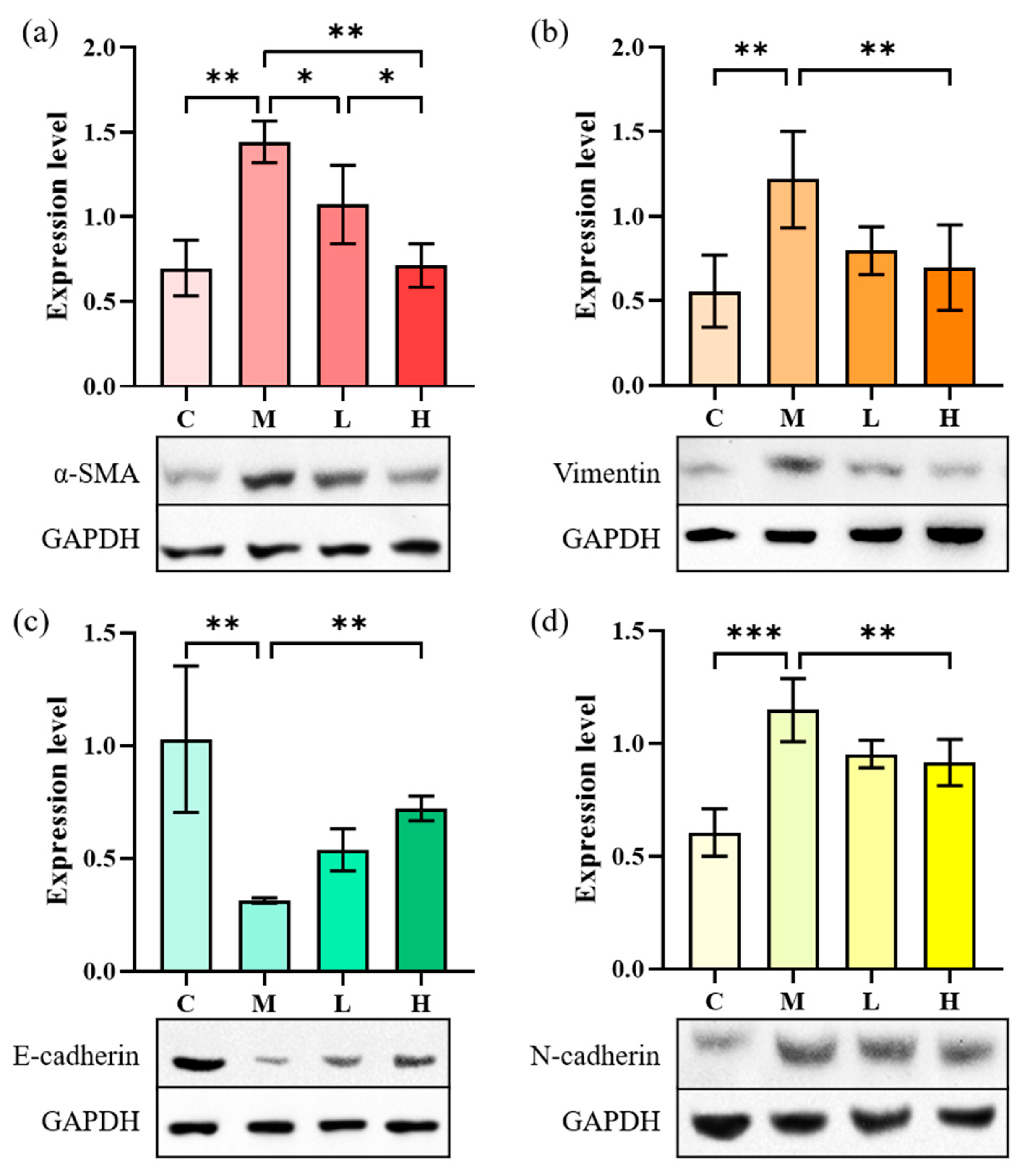
2.4.3. EMT-Related Proteins Expression
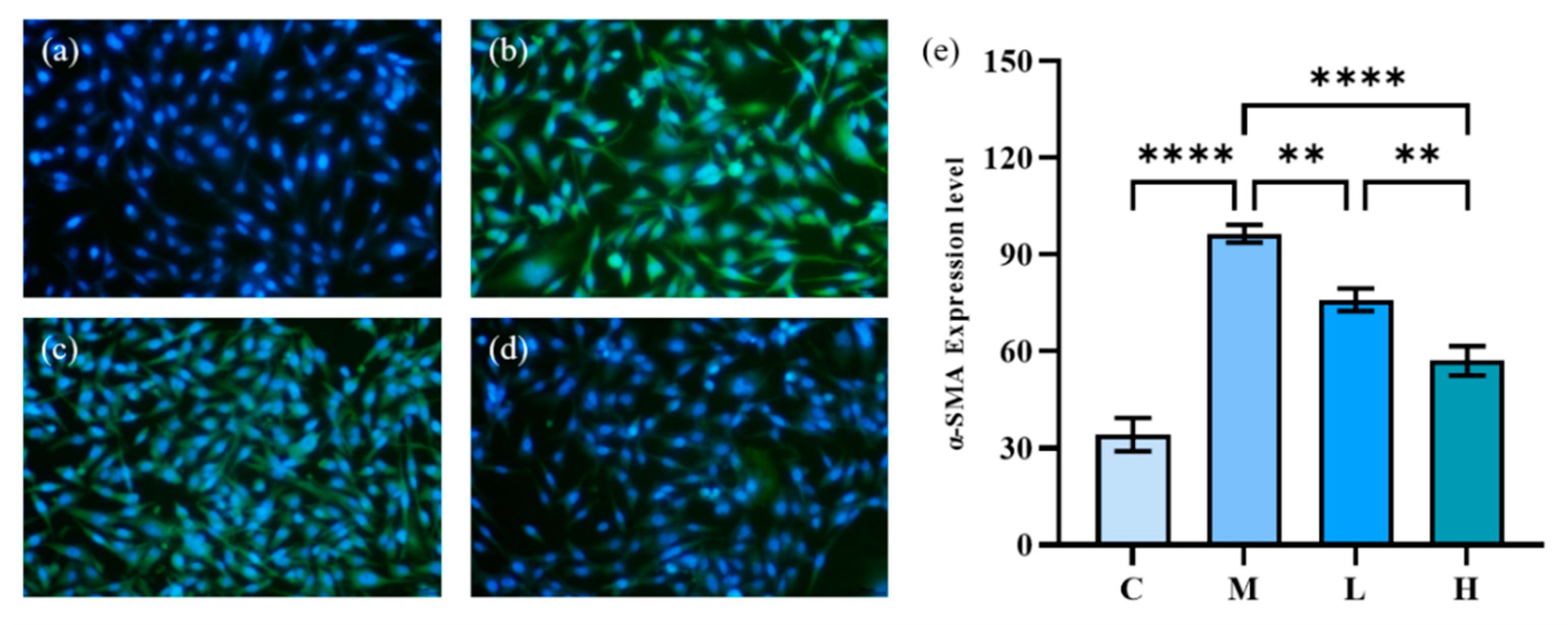
2.4.4. HFL-1 Cell α-SMA Expression
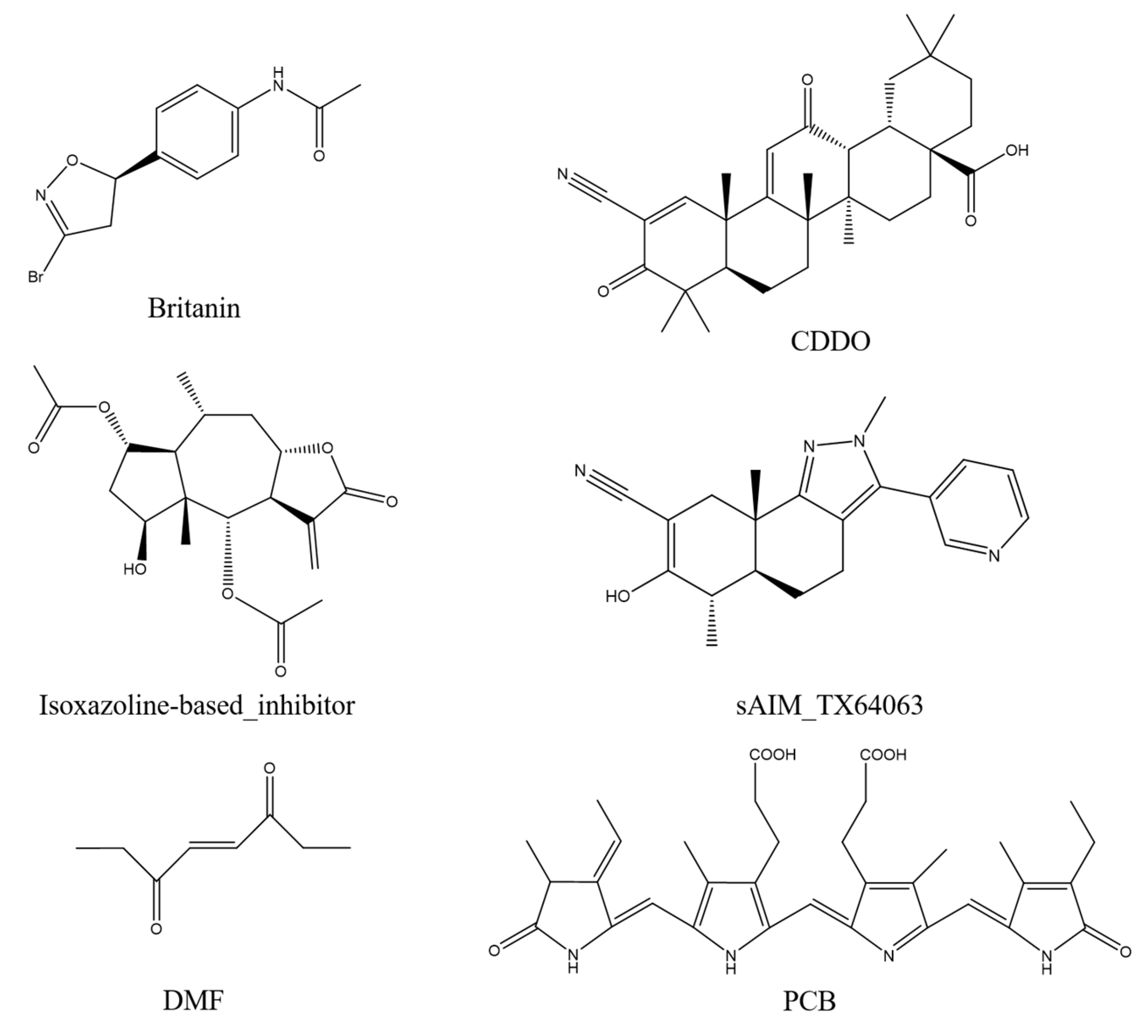
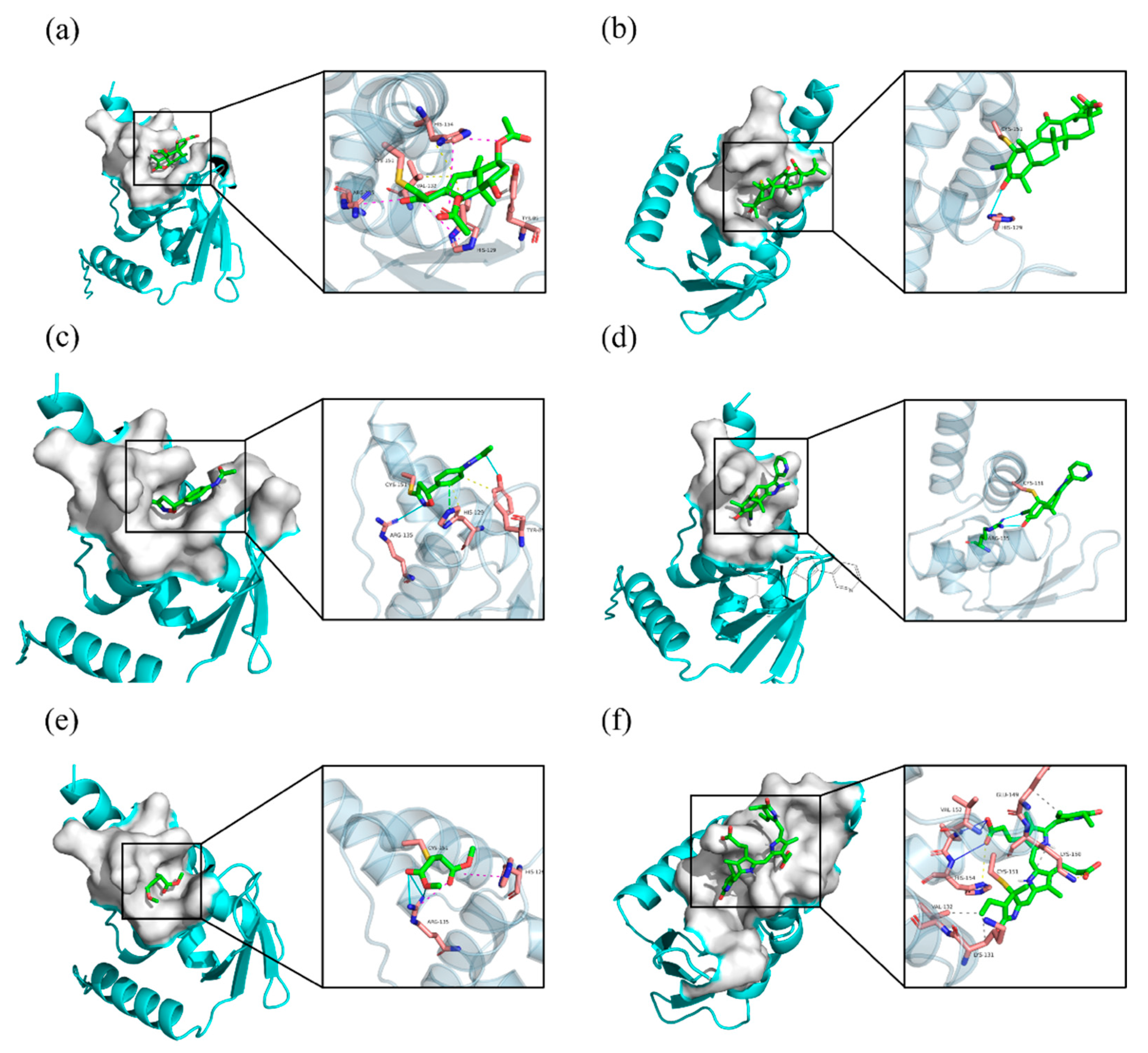
2.5. Covalent Docking
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Enzymatic Hydrolysis of Phycocyanin
4.3. Absorption Spectrum and Molecular Weight Distribution Measurement
4.4. Antioxidant Activity of Phycocyanin Peptide
4.4.1. Superoxide Anion Clearance
4.4.2. ABTS+ Clearance
4.4.3. Reducing Power
4.5. Anti-Inflammatory Activity of Phycocyanin Peptide
4.5.1. Cell Viability
4.5.2. Cytokines
4.6. Determination of Anti-Pulmonary Fibrosis Activity of Phycocyanin Peptide
4.6.1. Observation of A549 Cell Morphology
4.6.2. Immunofluorescence
4.6.3. Western Blot
4.7. Covalent Docking Analyze
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xin, W.; Jing, W.; Guichuan, H.; Yishi, L.; Shuliang, G. miR-320a-3P alleviates the epithelial-mesenchymal transition of A549 cells by activation of STAT3/SMAD3 signaling in a pulmonary fibrosis model. Mol. Med. Rep. 2021, 23, 357. [Google Scholar]
- Elko, E.; Anathy, V.; van der Velden, J.; van der Vliet, A.; Janssen-Heininger, Y. Oxidation state of peroxiredoxin 4 in lungs from patients with pulmonary fibrosis and mouse models of fibrosis. Free Radic. Biol. Med. 2018, 128, S25. [Google Scholar] [CrossRef]
- Qingbo, S.; Bingjing, L.; Hanmei, X.; Jialiang, H. Pathogenesis of Pulmonary Fibrosis and Progress in Drug Development. Prog. Pharm. Sci. 2018, 42, 868–873. [Google Scholar]
- Zhenhua, L.; Shouchun, P.; Jian, K.; Xianming, H.; Runjiang, Y. Efect of corticosteroids upon the prognosis of idiopathic pulmonary fibrosis. Natl. Med. J. China 2010, 90, 804–807. [Google Scholar]
- Antonio Rodriguez-Portal, J. Efficacy and Safety of Nintedanib for the Treatment of Idiopathic Pulmonary Fibrosis: An Update. Drugs RD 2018, 18, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skold, C.M.; Bendstrup, E.; Myllarniemi, M.; Gudmundsson, G.; Sjaheim, T.; Hilberg, O.; Altraja, A.; Kaarteenaho, R.; Ferrara, G. Treatment of idiopathic pulmonary fibrosis: A position paper from a Nordic expert group. J. Intern. Med. 2017, 281, 149–166. [Google Scholar] [CrossRef] [PubMed]
- Peltzer, N.; Walczak, H. Cell Death and Inflammation—A Vital but Dangerous Liaison. Trends Immunol. 2019, 40, 387–402. [Google Scholar] [CrossRef]
- Ndiaye, F.; Vuong, T.; Duarte, J.; Aluko, R.E.; Matar, C. Anti-oxidant, anti-inflammatory and immunomodulating properties of an enzymatic protein hydrolysate from yellow field pea seeds. Eur. J. Nutr. 2012, 51, 29–37. [Google Scholar] [CrossRef]
- Mao, X.Y.; Cheng, X.; Wang, X.; Wu, S.J. Free-radical-scavenging and anti-inflammatory effect of yak milk casein before and after enzymatic hydrolysis. Food Chem. 2011, 126, 484–490. [Google Scholar] [CrossRef]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage Activation and Polarization: Nomenclature and Experimental Guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Et Biophys. Acta-Mol. Cell Res. 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenhunen, R.; Marver, H.S.; Schmid, R. The Enzymatic Conversion of Heme to Bilirubin by Microsomal Heme Oxygenase. Proc. Natl. Acad. Sci. USA 1968, 61, 748–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jingjing, W.; Hui, Z.; Jianqiang, L. The anti-inflammatory effects of heme oxygenase-1 on emphysema model. Chin. J. Tuberc. Respir. Dis. 2015, 38, 379–383. [Google Scholar]
- Otterbein, L.E.; Kolls, J.K.; Mantell, L.L.; Cook, J.L.; Alam, J.; Choi, A.M. Exogenous administration of heme oxygenase-1 by gene transfer provides protection against hyperoxia-induced lung injury. J. Clin. Investig. 1999, 103, 1047–1054. [Google Scholar] [CrossRef] [PubMed]
- Otterbein, L.; Sylvester, S.L.; Choi, A.M. Hemoglobin provides protection against lethal endotoxemia in rats: The role of heme oxygenase-1. Am. J. Respir. Cell Mol. Biol. 1995, 13, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Shixiang, Z. Studies on the Anti-Hepatic Fibrosis Mechanism of Phycocyanin from Spirulina In Vitro and In Vivo. Master’s Thesis, University of Chinese Academy of Sciences, Beijing, China, 2020. [Google Scholar]
- Romay, C.; Armesto, J.; Remirez, D.; Gonzalez, R.; Ledon, N.; Garcia, I. Antioxidant and anti-inflammatory properties of C-phycocyanin from blue-green algae. Inflamm. Res. 1998, 47, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yu, Y.; Li, W.; Liu, B.; Jiao, X.; Song, X.; Lv, C.; Qin, S. Phycocyanin attenuates pulmonary fibrosis via the TLR2-MyD88-NF-kappa B signaling pathway. Sci. Rep. 2017, 7, 5843. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Li, W.; Qin, S. Therapeutic effect of phycocyanin on acute liver oxidative damage caused by X-ray. Biomed. Pharmacother. 2020, 130, 110553. [Google Scholar] [CrossRef]
- Campbell, N.K.; Fitzgerald, H.K.; Dunne, A. Regulation of inflammation by the antioxidant haem oxygenase 1. Nat. Rev. Immunol. 2021, 21, 411–425. [Google Scholar] [CrossRef]
- Nam, J.; Lee, Y.; Yang, Y.; Jeong, S.; Kim, W.; Yoo, J.W.; Moon, J.O.; Lee, C.; Chung, H.Y.; Kim, M.S.; et al. Is it worth expending energy to convert biliverdin into bilirubin? Free Radic. Bio. Med. 2018, 124, 232–240. [Google Scholar] [CrossRef]
- Collin, F. Chemical Basis of Reactive Oxygen Species Reactivity and Involvement in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deffert, C.; Cachat, J.; Krause, K.H. Phagocyte NADPH oxidase, chronic granulomatous disease and mycobacterial infections. Cell. Microbiol. 2014, 16, 1168–1178. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.R. Protein Oxidation and Aging. Science 1992, 257, 1220–1224. [Google Scholar] [CrossRef] [PubMed]
- Hui, W.; Yan, Z. The Method of ABTS Assay for Screening and Evaluating Antioxidant. Guangzhou Chem. Ind. 2012, 40, 41–43. [Google Scholar]
- Park, J.T.; Johnson, M.J. A Submicrodetermination of Glucose. J. Biol. Chem. 1949, 181, 149–151. [Google Scholar] [CrossRef]
- MacMicking, J.; Xie, Q.W.; Nathan, C. Nitric oxide and macrophage function. Annu. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef]
- Kraakman, M.J.; Kammoun, H.L.; Allen, T.L.; Deswaerte, V.; Henstridge, D.C.; Estevez, E.; Matthews, V.B.; Neill, B.; White, D.A.; Murphy, A.J.; et al. Blocking IL-6 trans-Signaling Prevents High-Fat Diet-Induced Adipose Tissue Macrophage Recruitment but Does Not Improve Insulin Resistance. Cell Metab. 2015, 21, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Elliott, S.; Cawston, T. The clinical potential of matrix metalloproteinase inhibitors in the rheumatic disorders. Drug Aging 2001, 18, 87–99. [Google Scholar] [CrossRef]
- Jung, K.A.; Kwak, M.K. The Nrf2 System as a Potential Target for the Development of Indirect Antioxidants. Molecules 2010, 15, 7266–7291. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.H.; Peng, W.; Zhang, Z.Y.; Pei, X.; Sun, Z.K.; Ou, Y. A phycocyanin derived eicosapeptide attenuates lung fibrosis development. Eur. J. Pharmacol. 2021, 908, 14. [Google Scholar] [CrossRef] [PubMed]
- Szapiel, S.V.; Elson, N.A.; Fulmer, J.D.; Hunninghake, G.W.; Crystal, R.G. Bleomycin-induced interstitial pulmonary disease in the nude, athymic mouse. Am. Rev. Respir. Dis. 1979, 120, 893–899. [Google Scholar]
- Linker, R.A.; Lee, D.-H.; Ryan, S.; van Dam, A.M.; Conrad, R.; Bista, P.; Zeng, W.; Hronowsky, X.; Buko, A.; Chollate, S.; et al. Fumaric acid esters exert neuroprotective effects in neuroinflammation via activation of the Nrf2 antioxidant pathway. Brain 2011, 134, 678–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Zhu, L.; Yuan, X.; Chen, H.; Xiong, R.; Zhang, S.; Cheng, H.; Shen, Y.; An, H.; Li, T.; et al. Britanin Ameliorates Cerebral Ischemia-Reperfusion Injury by Inducing the Nrf2 Protective Pathway. Antioxid. Redox Signal. 2017, 27, 754–768. [Google Scholar] [CrossRef] [PubMed]
- Cleasby, A.; Yon, J.; Day, P.J.; Richardson, C.; Tickle, I.J.; Williams, P.A.; Callahan, J.F.; Carr, R.; Concha, N.; Kerns, J.K.; et al. Structure of the BTB Domain of Keap1 and Its Interaction with the Triterpenoid Antagonist CDDO. PLoS ONE 2014, 9, e98896. [Google Scholar] [CrossRef]
- Pinto, A.; El Ali, Z.; Moniot, S.; Tamborini, L.; Steegborn, C.; Foresti, R.; De Micheli, C. Effects of 3-Bromo-4,5-dihydroisoxazole Derivatives on Nrf2 Activation and Heme Oxygenase-1 Expression. Chemistryopen 2018, 7, 858–864. [Google Scholar] [CrossRef]
- Huerta, C.; Jiang, X.; Trevino, I.; Bender, C.F.; Ferguson, D.A.; Probst, B.; Swinger, K.K.; Stoll, V.S.; Thomas, P.J.; Dulubova, I.; et al. Characterization of novel small-molecule NRF2 activators: Structural and biochemical validation of stereospecific KEAP1 binding. Biochim. Et Biophys. Acta-Gen. Subj. 2016, 1860, 2537–2552. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ma, J.; Fang, Q.; Guo, T.; Li, X. Protective effects of Nostoc sphaeroides Kütz against cyclophosphamide-induced immunosuppression and oxidative stress in mice. Toxin Rev. 2019, 40, 1118–1127. [Google Scholar] [CrossRef]
- Leung, P.O.; Lee, H.H.; Kung, Y.C.; Tsai, M.F.; Chou, T.C. Therapeutic Effect of C-Phycocyanin Extracted from Blue Green Algae in a Rat Model of Acute Lung Injury Induced by Lipopolysaccharide. Evid.-Based Compl. Alt. Med. 2013, 2013, 916590. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.J.; Yang, Y.H.; Liang, Y.C.; Chiu, C.J.; Chu, K.H.; Chou, H.N.; Chiang, B.L. A Novel Phycobiliprotein Alleviates Allergic Airway Inflammation by Modulating Immune Responses. Am. J. Resp. Crit. Care 2011, 183, 15–25. [Google Scholar] [CrossRef]
- Xia, D.; Liu, B.; Xin, W.Y.; Liu, T.S.; Sun, J.Y.; Liu, N.N.; Qin, S.; Du, Z.N. Protective effects of C-phycocyanin on alcohol-induced subacute liver injury in mice. J. Appl. Phycol. 2016, 28, 765–772. [Google Scholar] [CrossRef]
- Okada, K. HO1 and PcyA proteins involved in phycobilin biosynthesis form a 1:2 complex with ferredoxin-1 required for photosynthesis. FEBS Lett. 2009, 583, 1251–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, N.G.; Kappas, A. Pharmacological and clinical aspects of heme oxygenase. Pharmacol. Rev. 2008, 60, 79–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuji, N.; Tomohisa, T.; Yasuki, H. Heme oxygenase-1 and anti-inflammatory M2 macrophages. Arch. Biochem. Biophys. 2014, 564, 83–88. [Google Scholar]
- Chanda, D.; Otoupalova, E.; Smith, S.R.; Volckaert, T.; De Langhe, S.P.; Thannickal, V.J. Developmental pathways in the pathogenesis of lung fibrosis. Mol. Asp. Med. 2019, 65, 56–69. [Google Scholar] [CrossRef]
- Younis, T.; Khan, M.R.; Sajid, M.; Majid, M.; Zahra, Z.; Shah, N.A. Fraxinus xanthoxyloides leaves reduced the level of inflammatory mediators during in vitro and in vivo studies. BMC Complement. Altern. Med. 2016, 16, 230. [Google Scholar] [CrossRef] [Green Version]
- Strasky, Z.; Zemankova, L.; Nemeckova, I.; Rathouska, J.; Wong, R.J.; Muchova, L.; Subhanova, I.; Vanikova, J.; Vanova, K.; Vitek, L.; et al. Spirulina platensis and phycocyanobilin activate atheroprotective heme oxygenase-1: A possible implication for atherogenesis. Food Funct. 2013, 4, 1586–1594. [Google Scholar] [CrossRef]
- Qing-An, Z.; Xi, W.; Yun, S.; Xue-Hui, F.; Francisco, G.M.J. Optimization of Pyrogallol Autoxidation Conditions and Its Application in Evaluation of Superoxide Anion Radical Scavenging Capacity for Four Antioxidants. J. AOAC Int. 2016, 99, 504–511. [Google Scholar]
- Xuelian, M.; Jia, L.; Yingying, L.; Liangchao, Z.; Chengcheng, G.; Dan, W.; Jing, L.; Changlan, C. Inhibitory effect of cordycepin on macrophage hyperactivation induced by lipopolysaccharide. Sci. Technol. Food Ind. 2017, 38, 297–301, 306. [Google Scholar]
- Xing, M.; Guang, W.; Rui, T.; Yu, L.; Lanxin, X.; Cheng, T.; Wei, L.; Chao, C. Antioxidant Properties of Five Phycocyanins with Different Purities. Food Sci. 2021, 42, 17–23. [Google Scholar]
- Klompong, V.; Benjakul, S.; Kantachote, D.; Shahidi, F. Antioxidative activity and functional properties of protein hydrolysate of yellow stripe trevally (Selaroides leptolepis) as influenced by the degree of hydrolysis and enzyme type. Food Chem. 2007, 102, 1317–1327. [Google Scholar] [CrossRef]
- Gao, R.C.; Shu, W.H.; Shen, Y.; Sun, Q.C.; Bai, F.; Wang, J.L.; Li, D.J.; Li, Y.; Jin, W.G.; Yuan, L. Sturgeon protein-derived peptides exert anti-inflammatory effects in LPS-stimulated RAW264.7 macrophages via the MAPK pathway. J. Funct. Foods 2020, 72, 104044. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, Y.; Cao, F.; Qin, Y.; Li, W.; Zhang, J. Ganglioside GD1a suppresses LPS-induced pro-inflammatory cytokines in RAW264.7 macrophages by reducing MAPKs and NF-kappa B signaling pathways through TLR4. Int. Immunopharmacol. 2015, 28, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Pi, J.; Li, T.; Liu, J.; Su, X.; Wang, R.; Yang, F.; Bai, H.; Jin, H.; Cai, J. Detection of lipopolysaccharide induced inflammatory responses in RAW264.7 macrophages using atomic force microscope. Micron 2014, 65, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; Xin, Y.; Guo, Y.; Diao, Y.; Kou, X.; Luo, L.; Yin, Z. Ampelopsin reduces endotoxic inflammation via repressing ROS-mediated activation of PI3K/Akt/NF-kappa B signaling pathways. Int. Immunopharmacol. 2012, 12, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Bianco, G.; Forli, S.; Goodsell, D.S.; Olson, A.J. Covalent docking using autodock: Two-point attractor and flexible side chain methods. Protein Sci. 2016, 25, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP 2021: Expanding the scope of the protein-ligand interaction profiler to DNA and RNA. Nucleic Acids Res. 2021, 49, W530–W534. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicators | >10,000 | 10,000–5000 | 5000–3000 | 3000–2000 | 2000–1000 | 1000–500 | 500–180 | <180 |
|---|---|---|---|---|---|---|---|---|
| Peak area percentage (%, λ220 nm) | 0.72 | 0.39 | 0.58 | 1.81 | 6.83 | 20.56 | 50.14 | 18.96 |
| Number average molecular weight | 13,929 | 7536 | 3476 | 2444 | 1267 | 639 | 253 | – |
| Weight average molecular weight | 14,429 | 7826 | 3539 | 2477 | 1311 | 661 | 271 | – |
| total weight average molecular weight (Mw) | 583 | |||||||
| Seq Loc | Tgt Seq Mass | Sequence | Missed | Pred Mods |
|---|---|---|---|---|
| α(78-86) | 1646.8351 | QRGKDKCAR | 8 | PCB-α84 |
| β(80-82) | 849.3731 | AAC | 2 | PCB-β82 |
| β(81-84) | 1047.5212 | ACLR | 3 | PCB-β82 |
| β(80-84) | 1118.5583 | AACLR | 4 | PCB-β82 |
| β(81-86) | 1293.5886 | ACLRDM | 5 | PCB-β82 |
| β(80-86) | 1364.6257 | AACLRDM | 6 | PCB-β82 |
| β(152-155) | 980.395 | DCSA | 3 | PCB-β153 |
| β(152-156) | 1093.479 | DCSAL | 4 | PCB-β153 |
| β(148-155) | 1348.6009 | ITPGDCSA | 7 | PCB-β153 |
| β(148-156) | 1461.685 | ITPGDCSAL | 8 | PCB-β153 |
| β(147-155) | 1405.6224 | GITPGDCSA | 8 | PCB-β153 |
| Mode | Affinity (kcal/mol) | Clust. RMSD | Ref. RMSD | Clust. Size | RMSD Stdv | Energy Stdv | Best Run |
|---|---|---|---|---|---|---|---|
| Britanin | −3.4 | 0.0 | −1.0 | 7 | 0.4 | 0.2 | 003 |
| CDDO | −1.9 | 0.0 | −1.0 | 8 | 0.0 | 0.0 | 007 |
| Isoxazoline-based–inhibitor | −3.1 | 0.0 | −1.0 | 7 | 0.5 | 0.1 | 004 |
| DMF | −1.5 | 0.0 | −1.0 | 8 | 0.4 | 0.1 | 002 |
| sAIM_TX64063 | −2.1 | 0.0 | −1.0 | 8 | 0.2 | 0.1 | 001 |
| PCB | −3.8 | 0.0 | −1.0 | 1 | NA | NA | 002 |
| Control | Model | Low Dose | High Dose | |
|---|---|---|---|---|
| Medium | 1640 | 1640 | 1640 | 1640 |
| FBS | 10% | 10% | 10% | 10% |
| Temperature | 37 °C | 37 °C | 37 °C | 37 °C |
| CO2 | 5% | 5% | 5% | 5% |
| TGF-β1 | – | 10 ng/mL | 10 ng/mL | 10 ng/mL |
| Phycocyanin peptide | – | – | 10 µg/mL | 30 µg/mL |
| Sample size | 100 µL | 100 µL | 100 µL | 100 µL |
| Time | 72 h | 72 h | 72 h | 72 h |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, R.-Z.; Li, W.-J.; Zhang, J.-J.; Liu, Z.-Y.; Li, Y.; Liu, C.; Qin, S. The Inhibitory Effect of Phycocyanin Peptide on Pulmonary Fibrosis In Vitro. Mar. Drugs 2022, 20, 696. https://doi.org/10.3390/md20110696
Liu R-Z, Li W-J, Zhang J-J, Liu Z-Y, Li Y, Liu C, Qin S. The Inhibitory Effect of Phycocyanin Peptide on Pulmonary Fibrosis In Vitro. Marine Drugs. 2022; 20(11):696. https://doi.org/10.3390/md20110696
Chicago/Turabian StyleLiu, Run-Ze, Wen-Jun Li, Juan-Juan Zhang, Zheng-Yi Liu, Ya Li, Chao Liu, and Song Qin. 2022. "The Inhibitory Effect of Phycocyanin Peptide on Pulmonary Fibrosis In Vitro" Marine Drugs 20, no. 11: 696. https://doi.org/10.3390/md20110696
APA StyleLiu, R. -Z., Li, W. -J., Zhang, J. -J., Liu, Z. -Y., Li, Y., Liu, C., & Qin, S. (2022). The Inhibitory Effect of Phycocyanin Peptide on Pulmonary Fibrosis In Vitro. Marine Drugs, 20(11), 696. https://doi.org/10.3390/md20110696