
Rapid Biotic and Abiotic Transformation of Toxins produced by Ostreopsis. cf. ovata

Abstract
:1. Introduction
2. Results
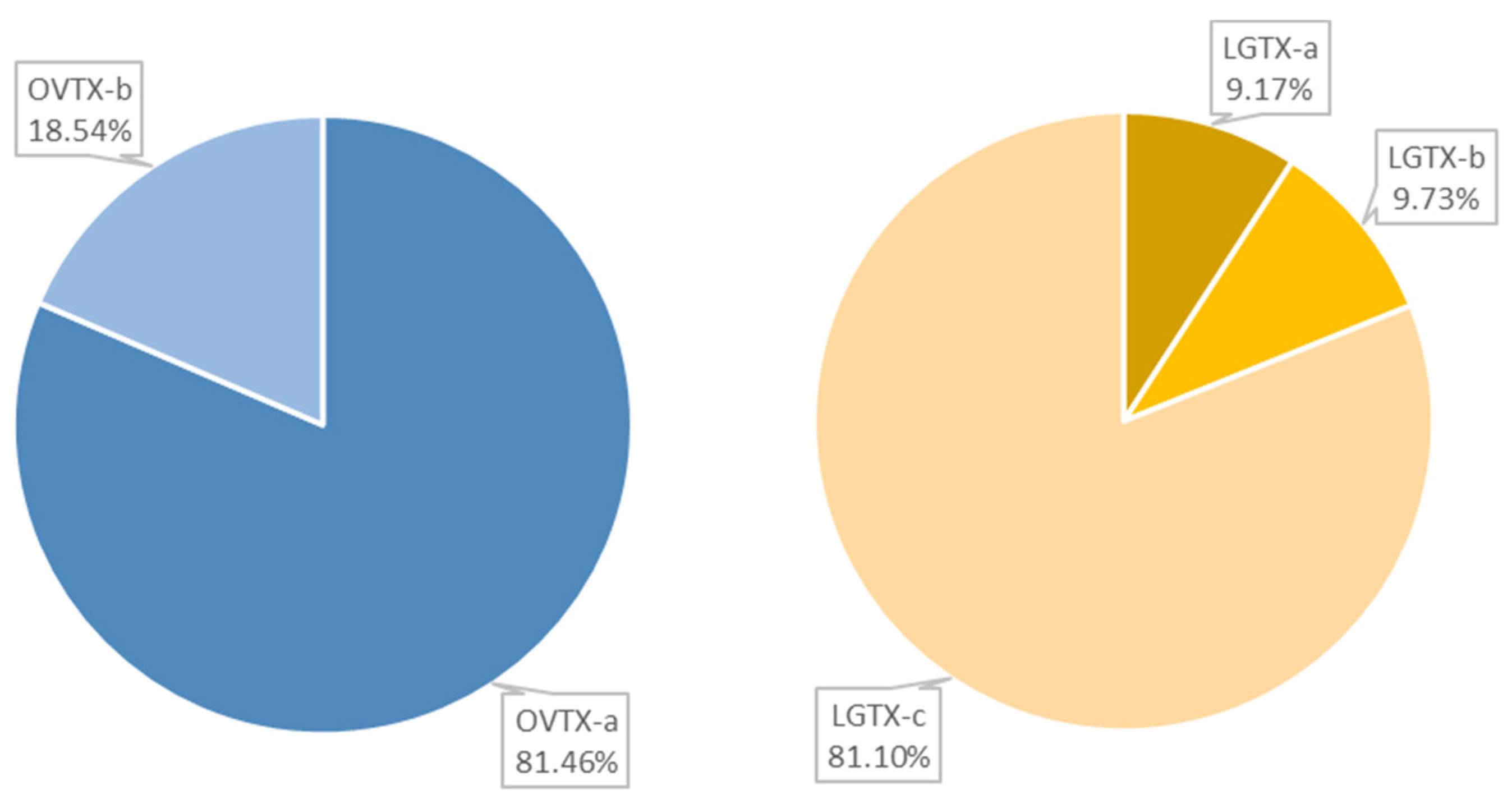
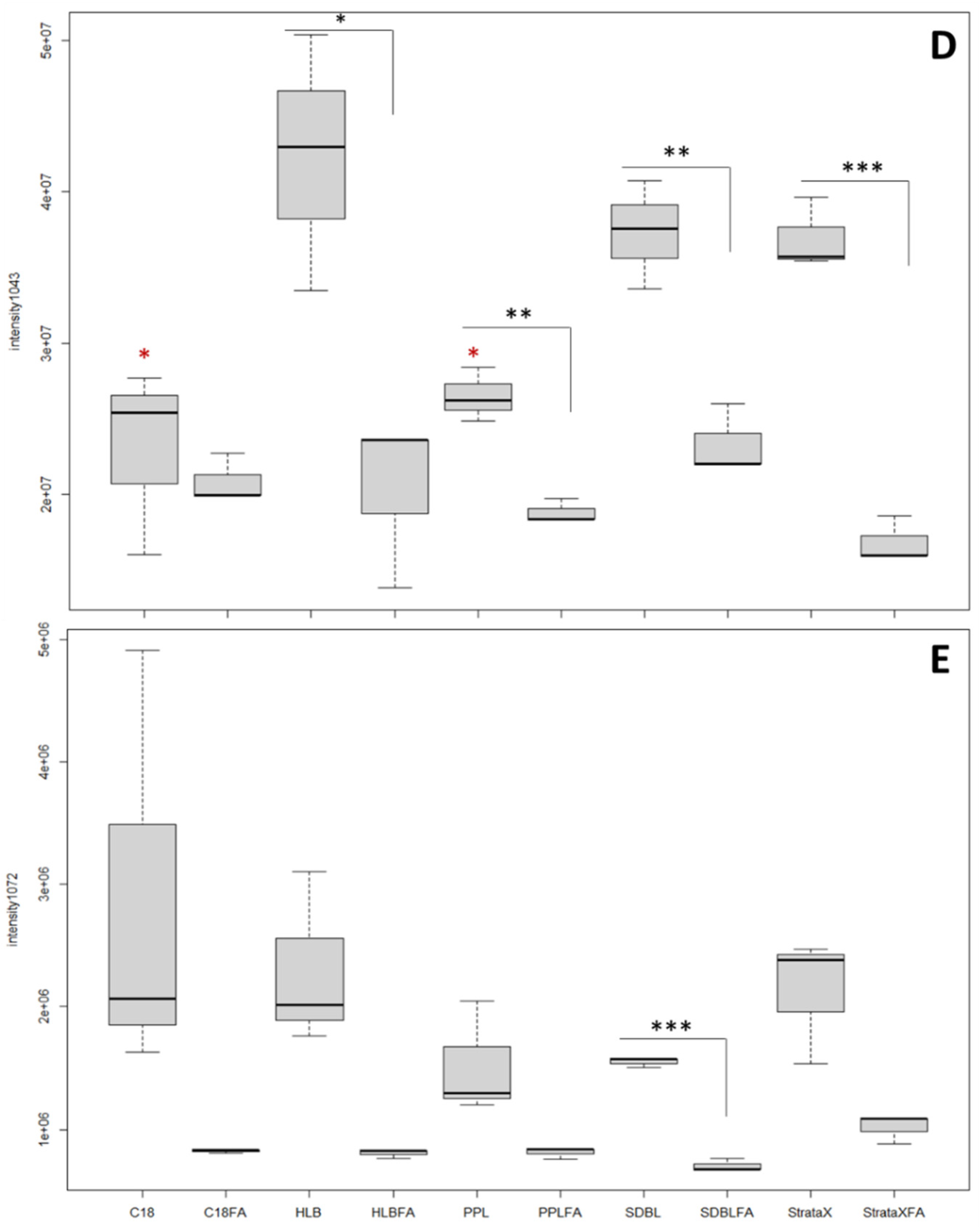
2.1. Cartridge Selection and Toxin Composition of O. cf. ovata’s Spent Medium
2.2. Biotic and Abiotic Transformation
2.3. Biotoxin By-Products
3. Discussion
3.1. Toxin Content and Spontaneous Stability
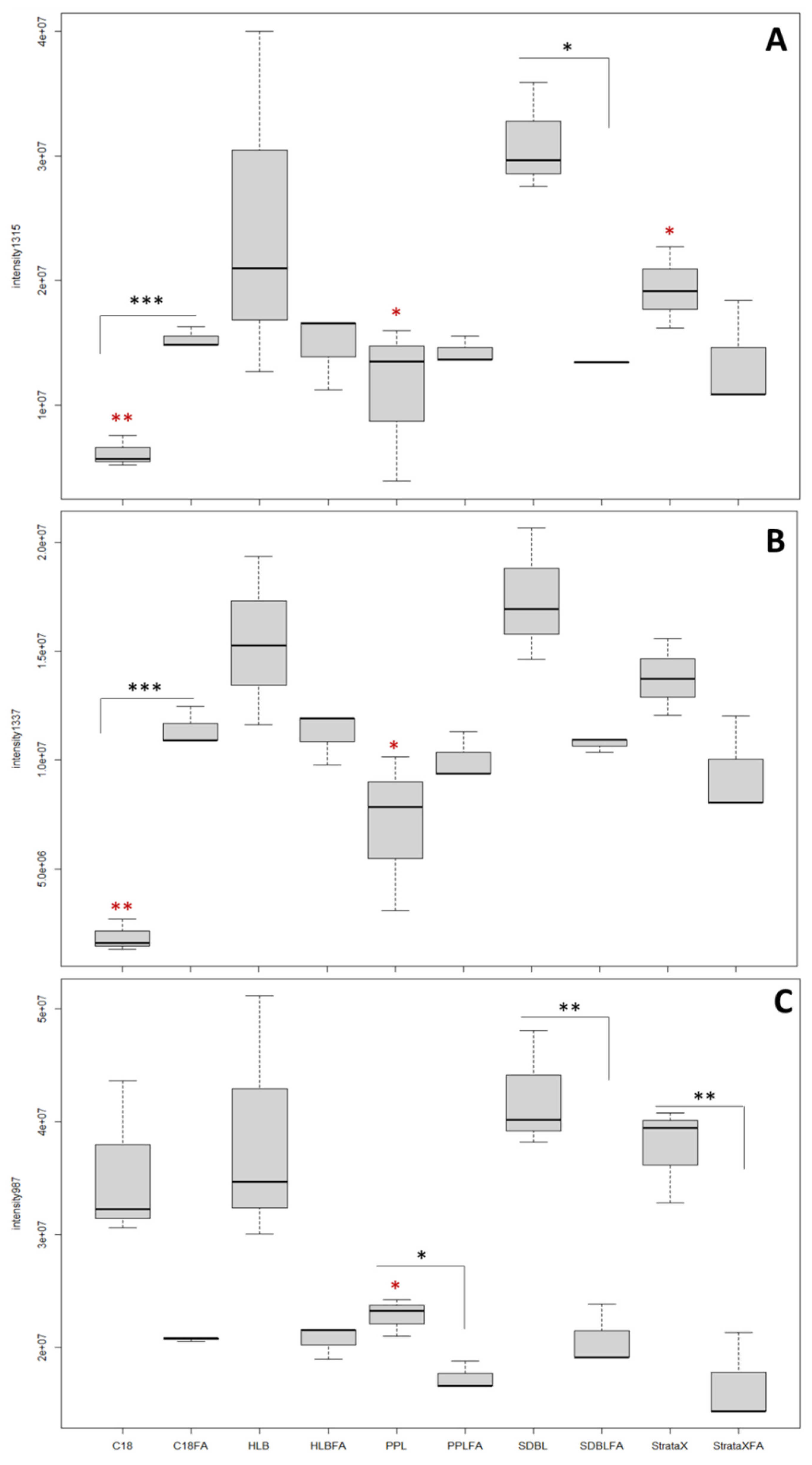
3.2. SDBL Cartridges Offer the Best Compromise for the Extraction of O. cf. ovata’s Toxins
3.3. Rapid Transformation of OVTX and LGTX by Bacteria
3.4. OVTXs Are more Sensitive to Light Than LGTX
4. Material and Methods
4.1. Microalgal Culture
4.2. Experimental Set up
4.2.1. Optimization of the Toxin Extraction
4.2.2. Biotic and Abiotic Transformation of the Toxins
4.3. Sample Treatment
4.4. Chemical Analysis
4.5. Bacterial Abundance
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Grillo, C.; Melchiorre, N. Putative Palytoxin and Its New Analogue, Ovatoxin-a, in Ostreopsis Ovata Collected along the Ligurian Coasts during the 2006 Toxic Outbreak. J. Am. Soc. Mass Spectrom. 2008, 19, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfannkuchen, M.; Godrijan, J.; Marić Pfannkuchen, D.; Iveša, L.; Kružić, P.; Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M. Toxin-Producing Ostreopsis Cf. Ovata Are Likely to Bloom Undetected along Coastal Areas. Environ. Sci. Technol. 2012, 46, 5574–5582. [Google Scholar] [CrossRef] [PubMed]
- Iddir-Ihaddaden, W.; Abdellah, K.O.; Merbout, G.; Abtroun, R.; Alamir, B.; de Haro, L. Health Impact of Unicellular Algae of the Ostreopsis Genus Blooms in Algeria during Summer 2009. Presse Med. 2013, 42, 1281–1283. [Google Scholar] [CrossRef] [PubMed]
- Tichadou, L.; Glaizal, M.; Armengaud, A.; Grossel, H.; Lemée, R.; Kantin, R.; Lasalle, J.-L.; Drouet, G.; Rambaud, L.; Malfait, P.; et al. Health Impact of Unicellular Algae of the Ostreopsis Genus Blooms in the Mediterranean Sea: Experience of the French Mediterranean Coast Surveillance Network from 2006 to 2009. Clin. Toxicol. 2010, 48, 839–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Guerrini, F.; Pezzolesi, L.; Pistocchi, R.; et al. Isolation and Structure Elucidation of Ovatoxin-a, the Major Toxin Produced by Ostreopsis Ovata. J. Am. Chem. Soc. 2012, 134, 1869–1875. [Google Scholar] [CrossRef] [PubMed]
- Brissard, C.; Herve, F.; Sibat, M.; Sechet, V.; Hess, P.; Amzil, Z.; Herrenknecht, C. Characterization of Ovatoxin-h, a New Ovatoxin Analog, and Evaluation of Chromatographic Columns for Ovatoxin Analysis and Purification. J. Chromatogr. A 2015, 1388, 87–101. [Google Scholar] [CrossRef] [Green Version]
- García-Altares, M.; Tartaglione, L.; Dell’Aversano, C.; Carnicer, O.; de la Iglesia, P.; Forino, M.; Diogène, J.; Ciminiello, P. The Novel Ovatoxin-g and Isobaric Palytoxin (so Far Referred to as Putative Palytoxin) from Ostreopsis Cf. Ovata (NW Mediterranean Sea): Structural Insights by LC-High Resolution MSn. Anal. Bioanal. Chem. 2015, 407, 1191–1204. [Google Scholar] [CrossRef]
- Ternon, E.; Glukhov, E.; Trytten, E.; Lemée, R.; Gerwick, W.H. On the Hunt for New Toxin Families Produced by a Mediterranean Strain of the Benthic Dinoflagellate Ostreopsis Cf. Ovata. Toxins 2022, 14, 234. [Google Scholar] [CrossRef]
- Lewis, E.R.; Schwartz, S.E. Sea Salt Aerosol Production: Mechanisms, Methods, Measurements, and Models; American Geophysical Union: Washington, DC, USA, 2004. [Google Scholar]
- Bertram, T.H.; Cochran, R.E.; Grassian, V.H.; Stone, E.A. Sea Spray Aerosol Chemical Composition: Elemental and Molecular Mimics for Laboratory Studies of Heterogeneous and Multiphase Reactions. Chem. Soc. Rev. 2018, 47, 2374–2400. [Google Scholar] [CrossRef] [PubMed]
- Van Acker, E.; Huysman, S.; De Rijcke, M.; Asselman, J.; De Schamphelaere, K.A.C.; Vanhaecke, L.; Janssen, C.R. Phycotoxin-Enriched Sea Spray Aerosols: Methods, Mechanisms, and Human Exposure. Environ. Sci. Technol. 2021, 55, 6184–6196. [Google Scholar] [CrossRef]
- Pezzolesi, L.; Guerrini, F.; Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Pistocchi, R. Influence of Temperature and Salinity on Ostreopsis Cf. Ovata Growth and Evaluation of Toxin Content through HR LC-MS and Biological Assays. Water Res. 2012, 46, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Ternon, E.; Pavaux, A.-S.; Marro, S.; Thomas, O.P.; Lemée, R. Allelopathic Interactions between the Benthic Toxic Dinoflagellate Ostreopsis Cf. Ovata and a Co-Occurring Diatom. Harmful Algae 2018, 75, 35–44. [Google Scholar] [CrossRef]
- Ternon, E.; Pavaux, A.-S.; Peltekis, A.; Gemin, M.-P.; Jauzein, C.; Bailleul, B.; Lemée, R.; Thomas, O.P. Assessment of the Allelochemical Activity of Ostreopsis Cf. Ovata and the Ovatoxins towards Competitive Benthic Microalgae. Aquat. Ecol. 2022, 56, 475–491. [Google Scholar] [CrossRef]
- Medina-Pérez, N.I.; Dall’Osto, M.; Decesari, S.; Paglione, M.; Moyano, E.; Berdalet, E. Aerosol Toxins Emitted by Harmful Algal Blooms Susceptible to Complex Air–Sea Interactions. Environ. Sci. Technol. 2021, 55, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Magno, G.S.; Tartaglione, L.; Grillo, C.; Melchiorre, N. The Genoa 2005 Outbreak. Determination of Putative Palytoxin in Mediterranean Ostreopsis Ovata by a New Liquid Chromatography Tandem Mass Spectrometry Method. Anal. Chem. 2006, 78, 6153–6159. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Benedettini, G.; Onorari, M.; Serena, F.; Battocchi, C.; et al. First Finding of Ostreopsis Cf. Ovata Toxins in Marine Aerosols. Environ. Sci. Technol. 2014, 48, 3532–3540. [Google Scholar] [CrossRef]
- Longnecker, K.; Soule, M.C.K.; Kujawinski, E.B. Dissolved Organic Matter Produced by Thalassiosira Pseudonana. Mar. Chem. 2015, 168, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Shibl, A.A.; Isaac, A.; Ochsenkuhn, M.A.; Cárdenas, A.; Fei, C.; Behringer, G.; Arnoux, M.; Drou, N.; Santos, M.P.; Gunsalus, K.C. Diatom Modulation of Select Bacteria through Use of Two Unique Secondary Metabolites. Proc. Natl. Acad. Sci. USA 2020, 117, 27445–27455. [Google Scholar] [CrossRef] [PubMed]
- Heal, K.R.; Durham, B.; Boysen, A.; Carlson, L.; Qin, W.; Ribalet, F.; White, A.; Bundy, R.M.; Armbrus, V.; Ingalls, A.E. Marine Community Metabolomes Carry Fingerprints of Phytoplankton Community Composition. mSystems 2021, 6, e01334-20. [Google Scholar] [CrossRef]
- Paul, C.; Mausz, M.A.; Pohnert, G. A Co-Culturing/Metabolomics Approach to Investigate Chemically Mediated Interactions of Planktonic Organisms Reveals Influence of Bacteria on Diatom Metabolism. Metabolomics 2013, 9, 349–359. [Google Scholar] [CrossRef]
- Wichard, T.; Beemelmanns, C. Role of Chemical Mediators in Aquatic Interactions across the Prokaryote–Eukaryote Boundary. J. Chem. Ecol. 2018, 44, 1008–1021. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-González, F.X.; Widner, B.; Holderman, N.R.; Glushka, J.; Edison, A.S.; Kujawinski, E.B.; Moran, M.A. Resource Partitioning of Phytoplankton Metabolites That Support Bacterial Heterotrophy. ISME J. 2021, 15, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Cottrell, M.T.; Kirchman, D.L. Natural Assemblages of Marine Proteobacteria and Members of the Cytophaga-Flavobacter Cluster Consuming Low-and High-Molecular-Weight Dissolved Organic Matter. Appl. Environ. Microbiol. 2000, 66, 1692–1697. [Google Scholar] [CrossRef] [Green Version]
- Alonso, C.; Pernthaler, J. Roseobacter and SAR11 Dominate Microbial Glucose Uptake in Coastal North Sea Waters. Environ. Microbiol. 2006, 8, 2022–2030. [Google Scholar] [CrossRef] [PubMed]
- Elifantz, H.; Dittel, A.I.; Cottrell, M.T.; Kirchman, D.L. Dissolved Organic Matter Assimilation by Heterotrophic Bacterial Groups in the Western Arctic Ocean. Aquat. Microb. Ecol. 2007, 50, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Mou, X.; Vila-Costa, M.; Sun, S.; Zhao, W.; Sharma, S.; Moran, M.A. Metatranscriptomic Signature of Exogenous Polyamine Utilization by Coastal Bacterioplankton. Environ. Microbiol. Rep. 2011, 3, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Sáez, L.; Gasol, J.M. Seasonal Variations in the Contributions of Different Bacterial Groups to the Uptake of Low-Molecular-Weight Compounds in Northwestern Mediterranean Coastal Waters. Appl. Environ. Microbiol. 2007, 73, 3528–3535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donovan, C.J.; Ku, J.C.; Quilliam, M.A.; Gill, T.A. Bacterial Degradation of Paralytic Shellfish Toxins. Toxicon 2008, 52, 91–100. [Google Scholar] [CrossRef]
- Shetty, K.G.; Huntzicker, J.V.; Rein, K.S.; Jayachandran, K. Biodegradation of Polyether Algal Toxins–Isolation of Potential Marine Bacteria. J. Environ. Sci. Health Part A 2010, 45, 1850–1857. [Google Scholar] [CrossRef] [Green Version]
- Christoffersen, K.; Lyck, S.; Winding, A. Microbial Activity and Bacterial Community Structure during Degradation of Microcystins. Aquat. Microb. Ecol. 2002, 27, 125–136. [Google Scholar] [CrossRef]
- Mopper, K.; Kieber, D.J.; Stubbins, A. Marine Photochemistry of Organic Matter: Processes and Impacts. In Biogeochemistry of Marine Dissolved Organic Matter; Academic Press: Cambridge, UK, 2015; pp. 389–450. [Google Scholar]
- Kujawinski, E.B.; Del Vecchio, R.; Blough, N.V.; Klein, G.C.; Marshall, A.G. Probing Molecular-Level Transformations of Dissolved Organic Matter: Insights on Photochemical Degradation and Protozoan Modification of DOM from Electrospray Ionization Fourier Transform Ion Cyclotron Resonance Mass Spectrometry. Mar. Chem. 2004, 92, 23–37. [Google Scholar] [CrossRef]
- Medeiros, P.M.; Seidel, M.; Powers, L.C.; Dittmar, T.; Hansell, D.A.; Miller, W.L. Dissolved Organic Matter Composition and Photochemical Transformations in the Northern North Pacific Ocean. Geophys. Res. Lett. 2015, 42, 863–870. [Google Scholar] [CrossRef]
- Stubbins, A.; Spencer, R.G.M.; Chen, H.; Hatcher, P.G.; Mopper, K.; Hernes, P.J.; Mwamba, V.L.; Mangangu, A.M.; Wabakanghanzi, J.N.; Six, J. Illuminated Darkness: Molecular Signatures of Congo River Dissolved Organic Matter and Its Photochemical Alteration as Revealed by Ultrahigh Precision Mass Spectrometry. Limnol. Oceanogr. 2010, 55, 1467–1477. [Google Scholar] [CrossRef]
- Kieber, R.J.; Zhou, X.; Mopper, K. Formation of Carbonyl Compounds from UV-Induced Photodegradation of Humic Substances in Natural Waters: Fate of Riverine Carbon in the Sea. Limnol. Oceanogr. 1990, 35, 1503–1515. [Google Scholar] [CrossRef]
- Cooper, W.J.; Zika, R.G.; Petasne, R.G.; Fischer, A.M. Sunlight-Induced Photochemistry of Humic Substances in Natural Waters: Major Reactive Species. In Aquatic Humic Substances; Advances in Chemistry; American Chemical Society: Washington, DC, USA, 1988; Volume 219, pp. 333–362. ISBN 978-0-8412-1428-6. [Google Scholar]
- Vione, D.; Minella, M.; Maurino, V.; Minero, C. Indirect Photochemistry in Sunlit Surface Waters: Photoinduced Production of Reactive Transient Species. Chem. A Eur. J. 2014, 20, 10590–10606. [Google Scholar] [CrossRef] [PubMed]
- Stubbins, A.; Dittmar, T. Illuminating the Deep: Molecular Signatures of Photochemical Alteration of Dissolved Organic Matter from North Atlantic Deep Water. Mar. Chem. 2015, 177, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Stevens, D.K.; Krieger, R.I. Stability Studies on the Cyanobacterial Nicotinic Alkaloid Anatoxin-A. Toxicon 1991, 29, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Hardman, R.C. Harmful Algal Blooms in the Gulf of Mexico: Brevetoxin Degradation and Derivative Formation via Photochemical Processes; University of North Carolina at Wilmington: Wilmington, NC, USA, 2002; 110p. [Google Scholar]
- Bates, S.S.; Léger, C.; Wells, M.L.; Hardy, K. Photodegradation of Domoic Acid. Can. Tech. Rep. Fish. Aquat. Sci./Rapp. Tech. Can. Sci. Halieut. Aquat 2003, 30–35. [Google Scholar]
- Bouillon, R.; Knierim, T.L.; Kieber, R.J.; Skrabal, S.A.; Wright, J.L.C. Photodegradation of the Algal Toxin Domoic Acid in Natural Water Matrices. Limnol. Oceanogr. 2006, 51, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Cohu, S.; Mangialajo, L.; Thibaut, T.; Blanfuné, A.; Marro, S.; Lemée, R. Proliferation of the Toxic Dinoflagellate Ostreopsis Cf. Ovata in Relation to Depth, Biotic Substrate and Environmental Factors in the North West Mediterranean Sea. Harmful Algae 2013, 24, 32–44. [Google Scholar] [CrossRef]
- Carnicer, O.; Guallar, C.; Andree, K.B.; Diogène, J.; Fernández-Tejedor, M. Ostreopsis Cf. Ovata Dynamics in the NW Mediterranean Sea in Relation to Biotic and Abiotic Factors. Environ. Res. 2015, 143, 89–99. [Google Scholar] [CrossRef]
- Pavaux, A.-S.; Velasquez-Carjaval, D.; Drouet, K.; Lebrun, A.; Hiroux, A.; Marro, S.; Christians, E.; Castagnetti, S.; Lemee, R. Daily Variations of Ostreopsis Cf. Ovata Abundances in NW Mediterranean Sea. Harmful Algae 2021, 110, 102144. [Google Scholar] [CrossRef] [PubMed]
- Gémin, M.-P.; Réveillon, D.; Hervé, F.; Pavaux, A.-S.; Tharaud, M.; Séchet, V.; Bertrand, S.; Lemée, R.; Amzil, Z. Toxin Content of Ostreopsis Cf. Ovata Depends on Bloom Phases, Depth and Macroalgal Substrate in the NW Mediterranean Sea. Harmful Algae 2020, 92, 101727. [Google Scholar] [CrossRef]
- Dittmar, T.; Koch, B.; Hertkorn, N.; Kattner, G. A Simple and Efficient Method for the Solid-Phase Extraction of Dissolved Organic Matter (SPE-DOM) from Seawater. Limnol. Oceanogr. Methods 2008, 6, 230–235. [Google Scholar] [CrossRef]
- Zendong, Z.; Herrenknecht, C.; Abadie, E.; Brissard, C.; Tixier, C.; Mondeguer, F.; Sechet, V.; Amzil, Z.; Hess, P. Extended Evaluation of Polymeric and Lipophilic Sorbents for Passive Sampling of Marine Toxins. Toxicon 2014, 91, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Brissard, C.; Herrenknecht, C.; Sechet, V.; Herve, F.; Pisapia, F.; Harcouet, J.; Lemee, R.; Chomerat, N.; Hess, P.; Amzil, Z. Complex Toxin Profile of French Mediterranean Ostreopsis Cf. Ovata Strains, Seafood Accumulation and Ovatoxins Prepurification. Mar. Drugs 2014, 12, 2851–2876. [Google Scholar] [CrossRef] [Green Version]
- Tartaglione, L.; Dello Iacovo, E.; Mazzeo, A.; Casabianca, S.; Ciminiello, P.; Penna, A.; Dell’Aversano, C. Variability in Toxin Profiles of the Mediterranean Ostreopsis Cf. Ovata and in Structural Features of the Produced Ovatoxins. Environ. Sci. Technol. 2017, 51, 13920–13928. [Google Scholar] [CrossRef]
- Cancelada, L.; Torres, R.R.; Garrafa Luna, J.; Dorrestein, P.C.; Aluwihare, L.I.; Prather, K.A.; Petras, D. Assessment of Styrene-Divinylbenzene Polymer (PPL) Solid-Phase Extraction and Non-Targeted Tandem Mass Spectrometry for the Analysis of Xenobiotics in Seawater. Limnol. Oceanogr. Methods 2022, 20, 89–101. [Google Scholar] [CrossRef]
- Seidel, M.; Yager, P.L.; Ward, N.D.; Carpenter, E.J.; Gomes, H.R.; Krusche, A.V.; Richey, J.E.; Dittmar, T.; Medeiros, P.M. Molecular-Level Changes of Dissolved Organic Matter along the Amazon River-to-Ocean Continuum. Mar. Chem. 2015, 177, 218–231. [Google Scholar] [CrossRef]
- Medeiros, P.M.; Seidel, M.; Gifford, S.M.; Ballantyne, F.; Dittmar, T.; Whitman, W.B.; Moran, M.A. Microbially-Mediated Transformations of Estuarine Dissolved Organic Matter. Front. Mar. Sci. 2017, 4, 69. [Google Scholar] [CrossRef] [Green Version]
- Myers, T.L.; Prince, E.K.; Naar, J.; Kubanek, J. Loss of Waterborne Brevetoxins from Exposure to Phytoplankton Competitors. Harmful Algae 2008, 7, 762–771. [Google Scholar] [CrossRef]
- Redshaw, C.H.; Sutter, D.; Myers, T.L.; Naar, J.; Kubanek, J. Tracking Losses of Brevetoxins on Exposure to Phytoplankton Competitors: Mechanistic Insights. Aquat. Toxicol. 2010, 100, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith Elizabeth, A.; Faye, G.; Ferguson Carolyn, M.J. Gallacher Susan Biotransformations of Paralytic Shellfish Toxins by Bacteria Isolated from Bivalve Molluscs. Appl. Environ. Microbiol. 2001, 67, 2345–2353. [Google Scholar] [CrossRef] [Green Version]
- Guidi, F.; Pezzolesi, L.; Vanucci, S. Microbial Dynamics during Harmful Dinoflagellate Ostreopsis Cf. Ovata Growth: Bacterial Succession and Viral Abundance Pattern. MicrobiologyOpen 2018, 7, e00584. [Google Scholar] [CrossRef]
- Vanucci, S.; Guidi, F.; Pistocchi, R.; Long, R.A. Phylogenetic Structure of Bacterial Assemblages Co-Occurring with Ostreopsis Cf. Ovata Bloom. Harmful Algae 2016, 55, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Chiswell, R.K.; Shaw, G.R.; Eaglesham, G.; Smith, M.J.; Norris, R.L.; Seawright, A.A.; Moore, M.R. Stability of Cylindrospermopsin, the Toxin from the Cyanobacterium, Cylindrospermopsis Raciborskii: Effect of PH, Temperature, and Sunlight on Decomposition. Environ. Toxicol. 1999, 14, 155–161. [Google Scholar] [CrossRef]
- Mazur-Marzec, H.; Meriluoto, J.; Pliński, M. The Degradation of the Cyanobacterial Hepatotoxin Nodularin (NOD) by UV Radiation. Chemosphere 2006, 65, 1388–1395. [Google Scholar] [CrossRef]
- Welker, M.; Steinberg, C. Rates of Humic Substance Photosensitized Degradation of Microcystin-LR in Natural Waters. Environ. Sci. Technol. 2000, 34, 3415–3419. [Google Scholar] [CrossRef]
- Timko, S.; Maydanov, A.; Pittelli, S.; Conte, M.; Cooper, W.; Koch, B.; Schmitt-Kopplin, P.; Gonsior, M. Depth-Dependent Photodegradation of Marine Dissolved Organic Matter. Front. Mar. Sci. 2015, 2, 66. [Google Scholar] [CrossRef] [Green Version]
- Kaminski, A.; Bober, B.; Lechowski, Z.; Bialczyk, J. Determination of Anatoxin-a Stability under Certain Abiotic Factors. Harmful Algae 2013, 28, 83–87. [Google Scholar] [CrossRef]
- Song, W.; Yan, S.; Cooper, W.J.; Dionysiou, D.D.; O’Shea, K.E. Hydroxyl Radical Oxidation of Cylindrospermopsin (Cyanobacterial Toxin) and Its Role in the Photochemical Transformation. Environ. Sci. Technol. 2012, 46, 12608–12615. [Google Scholar] [CrossRef]
- Rosario-Ortiz, F.L.; Canonica, S. Probe Compounds to Assess the Photochemical Activity of Dissolved Organic Matter. Environ. Sci. Technol. 2016, 50, 12532–12547. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Blasiak, L.C.; Karolin, J.O.; Powell, R.J.; Geddes, C.D.; Hill, R.T. Phosphorus Sequestration in the Form of Polyphosphate by Microbial Symbionts in Marine Sponges. Proc. Natl. Acad. Sci. USA 2015, 112, 4381–4386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, K.M.; Mitch, W.A. Halogen Radicals Contribute to Photooxidation in Coastal and Estuarine Waters. Proc. Natl. Acad. Sci. USA 2016, 113, 5868–5873. [Google Scholar] [CrossRef] [Green Version]
- Bouillon, R.-C.; Kieber, R.J.; Skrabal, S.A.; Wright, J.L.C. Photochemistry and Identification of Photodegradation Products of the Marine Toxin Domoic Acid. Mar. Chem. 2008, 110, 18–27. [Google Scholar] [CrossRef]
- Jin, H.; Lian, L.; Zhou, H.; Yan, S.; Song, W. Mechanistic Consideration of the Photochemical Transformation of Domoic Acid (Algal Toxin) in DOM-Rich Brackish Water. Chemosphere 2018, 209, 328–337. [Google Scholar] [CrossRef]
- Moore, R.E.; Woolard, F.X.; Bartolini, G. Periodate Oxidation of N-(p-Bromobenzoyl)Palytoxin. J. Am. Chem. Soc. 1980, 102, 7370–7372. [Google Scholar] [CrossRef]
- Moore, R.E.; Bartolini, G. Structure of Palytoxin. J. Am. Chem. Soc. 1981, 103, 2491–2494. [Google Scholar] [CrossRef]
- Selwood, A.I.; van Ginkel, R.; Harwood, D.T.; McNabb, P.S.; Rhodes, L.R.; Holland, P.T. A Sensitive Assay for Palytoxins, Ovatoxins and Ostreocins Using LC-MS/MS Analysis of Cleavage Fragments from Micro-Scale Oxidation. Toxicon 2012, 60, 810–820. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Ryther, J.H. Studies of Marine Planktonic Diatoms; I. Cyclotella Nana Hustedt, and Detonula Confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J. A Cross-Platform Toolkit for Mass Spectrometry and Proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef] [PubMed]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular Framework for Processing, Visualizing, and Analyzing Mass Spectrometry-Based Molecular Profile Data. BMC Bioinform. 2010, 11, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nothias, L.-F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M. Feature-Based Molecular Networking in the GNPS Analysis Environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef]
- Gasol, J.M.; del Giorgio, P.A. Using Flow Cytometry for Counting Natural Planktonic Bacteria and Understanding the Structure of Planktonic Bacterial Communities. Sci. Mar. 2000, 64, 197–224. [Google Scholar] [CrossRef] [Green Version]
- Chomérat, N.; Antajan, E.; Auby, I.; Bilien, G.; Carpentier, L.; Casamajor, M.-N.d.; Ganthy, F.; Hervé, F.; Labadie, M.; Méteigner, C.; et al. First Characterization of Ostreopsis cf. Ovata (Dinophyceae) and Detection of Ovatoxins during a Multispecific and Toxic Ostreopsis Bloom on French Atlantic Coast. Mar. Drugs 2022, 20, 461. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | m/z | RT | Ion | Molecular Formula | Score Formula (%) | GNPS | Diagnostic Fragments | ||
|---|---|---|---|---|---|---|---|---|---|
| BACTERIA | Lost | 1 | 1072.0619 | 4.48 | [M + 2H]2+ | LGTX-c | - | singleton | 1057.55 ( + losses water); 979.50 ( + losses water); 655.31; 590.78; 350.47 |
| 2 | 647.4052 | 5.79 | [M + H]+ | C39H54N2O6 | 85.79 | singleton | 629.39; 603.41; 550.82; 497.23; 456.27; 398.26; 316.22; 250.14; 192.13 | ||
| 3 | 997.6001 | 4.59 | [M + H]+ | nd | - | cluster | 920.46; 911.45; 902.45; 893.44; 884.44 (all dicharged = loss water); 865.55; 464.26; 420.23; 360.21; 302.21 | ||
| 4 | 1013.5953 | 3.92 | [M + H]+ | nd | - | cluster | 881.55; 464.26; 420.23; 360.21; 302.21 | ||
| 5 | 459.3292 | 13.74 | [M + Na]+ | C23H48O7 | 85.3 | cluster | 415.29; 403.20; 371.27; 345.24; 305.15; 275.20; 261.11; 217.09 | ||
| 6 | 997.5004 | 3.85 | [M + 2H]2+ | nd | - | singleton | 1618.81?; 1098.55; 979.98; 824.39; 842.39; 806.37; 466.22; 448.21; 430.20; 360.21; 306.17 | ||
| 7 | 1007.5271 | 4.64 | [M + 2H]2+ | LGTX-a | - | - | 989.51 ( + water losses); 920.46 ( + water losses); 652.26; 540.61; 351.35 | ||
| 8 | 865.5573 | 4.99 | [M + H]+ | C45H76N4O12 | 52.02 | cluster | 847.54; 643.43; 602.40; 560.35; 464.26; 420.23; 360.21; 302.21; 244.17 | ||
| Gained | 1 | 863.4252 | 8.85 | [M + H]+ | nd | - | - | ||
| 2 | 888.8663 | 4.60 | [M + Fe]2+ | nd | - | - | 823.9066; 814.8993; 805.89; 796.89; 787.88; 778.89; 670.8027; 521.73; 464.26; 420.23 | ||
| PHOTOLYSIS | Lost | 1 | 343.0887 | 2.36 | [M + Na]+ | C11H20N4O3S2 | 59.46 | singleton | 336.06; 319.01; 277.00; 258.99; 240.98 |
| 2 | 997.5004 | 3.85 | [M + 2H]2+ | nd | - | singleton | 1618.81?; 1098.55; 979.98; 824.39; 842.39; 806.37; 466.22; 448.21; 360.21; | ||
| 3 | 329.1102 | 0.76 | [M + Na]+ | C13H24NO3S2 | 80.35 | singleton | 299.09; 281.10; 253.11; 219.12; 167.09; 139.09; 122.07; 110.07 | ||
| 4 | 647.4052 | 5.79 | [M + H]+ | C39H54N2O6 | 85.79 | singleton | 603.41; 550.82; 497.23; 456.27; 398.26; 316.22; 273.10; 192.13 | ||
| 5 | 387.0881 | 2.36 | [M + H]+ | nd | - | cluster | no MS2 | ||
| 6 | 865.5573 | 4.94 | [M + H]+ | C45H76N4O12 | 52.02 | cluster | 847.54; 643.43; 602.40; 560.35; 464.26; 420.23; 360.21; 302.21; 244.17 | ||
| 7 | 285.2904 | 6.09 | [M + H]+ | C17H36N2O | 100 | cluster | 263.0188; 240.2317 | ||
| 8 | 271.2422 | 8.45 | [M + H]+ | C20H30 | 100 | singleton | odd isotopic pattern | ||
| Gained | 1 | 309.2062 | 9.26 | [M + H]+ | C18H28O4 | 100 | cluster | 281.21; 277.17; 253.14; 221.111; 193.12; 175.11; 165.05; 115.03; 95.08; 85.02 | |
| 2 | 409.2070 | 8.26 | [M + H]+ | - | - | ||||
| 3 | 409.2070 | 9.26 | [M + H]+ | - | - | ||||
| 4 | 330.2275 | 7.29 | [M + H]+ | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ternon, E.; Thomas, O.P.; Lemée, R.; Gerwick, W.H. Rapid Biotic and Abiotic Transformation of Toxins produced by Ostreopsis. cf. ovata. Mar. Drugs 2022, 20, 748. https://doi.org/10.3390/md20120748
Ternon E, Thomas OP, Lemée R, Gerwick WH. Rapid Biotic and Abiotic Transformation of Toxins produced by Ostreopsis. cf. ovata. Marine Drugs. 2022; 20(12):748. https://doi.org/10.3390/md20120748
Chicago/Turabian StyleTernon, Eva, Olivier P. Thomas, Rodolphe Lemée, and William H. Gerwick. 2022. "Rapid Biotic and Abiotic Transformation of Toxins produced by Ostreopsis. cf. ovata" Marine Drugs 20, no. 12: 748. https://doi.org/10.3390/md20120748
APA StyleTernon, E., Thomas, O. P., Lemée, R., & Gerwick, W. H. (2022). Rapid Biotic and Abiotic Transformation of Toxins produced by Ostreopsis. cf. ovata. Marine Drugs, 20(12), 748. https://doi.org/10.3390/md20120748