2.3.1. Radical Scavenging Activity
The superoxide anion radical is a physiological free radical with extreme importance for human body. When there is an overproduction of this ROS during aerobic respiration, or when the endogenous detoxification mechanisms fail or are insufficient, there is an increased risk of oxidative damage with serious deleterious effects. In this sense, finding mechanisms to inhibit the deleterious effects of O
2•− is of key importance, not only in the field of cosmetics, but also considering the amelioration and prevention of a wide array of diseases. The O
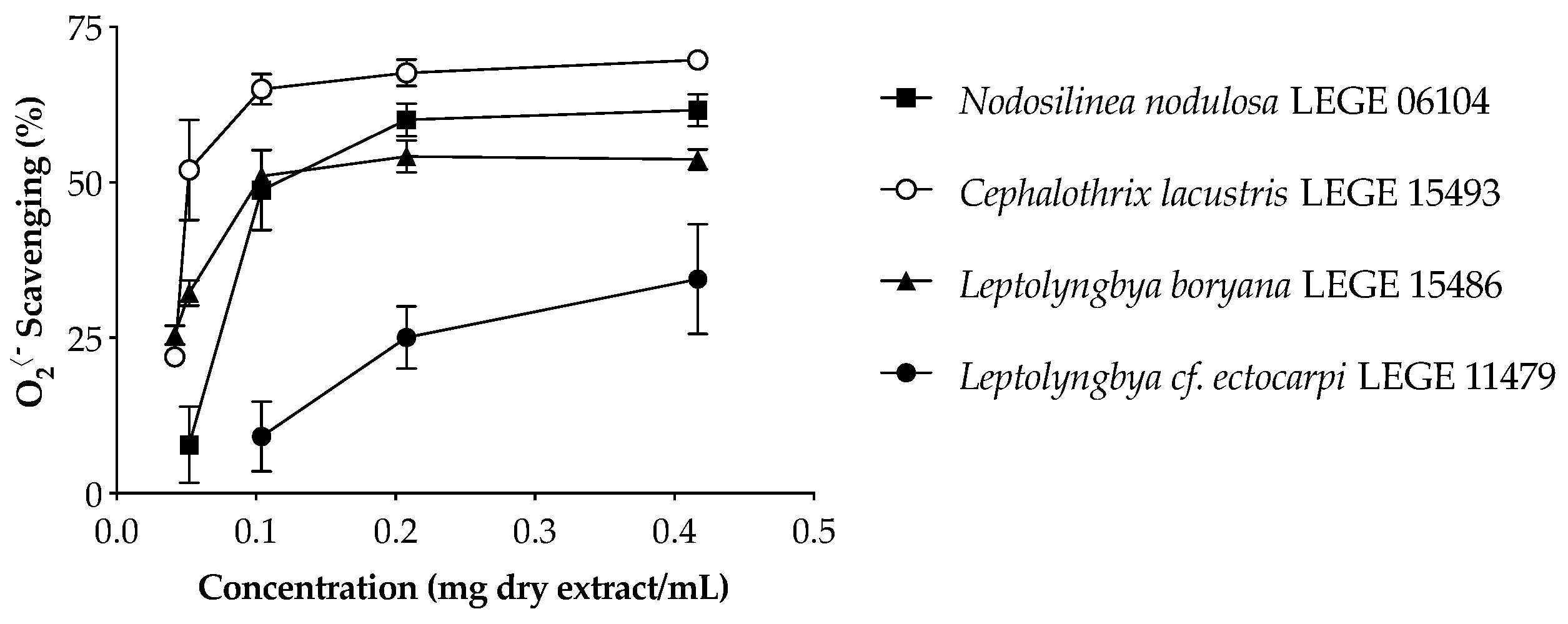
2•− scavenging behavior of cyanobacteria extracts is displayed in
Figure 1, and the IC values are summarized in
Table 6.
The aqueous extracts were significantly more effective in O
2•− scavenging than the acetone extracts (
Table 6) and presented a dose-dependent activity (
Figure 1), with the freshwater strains standing out from the marine strains.
Cephalothrix lacustris LEGE 15493 was the most effective strain, presenting the lowest IC
50 value (65.5 µg dry extract/mL,
p < 0.05), followed by
Leptolyngbya boryana LEGE 15486 and
Nodosilinea nodulosa LEGE 06104.
Leptolyngbya cf.
ectocarpi LEGE 11479 was the only strain which did not reach the IC
50 for the aqueous extract (
Table 6). On the other hand, the acetone extract of this strain was the most effective in O
2•− scavenging.
A comparison of the antioxidant capacities among different cyanobacteria is challenging due to the different methods applied. Morone and co-workers evaluated the radical scavenging ability of ethanol (70%
v/v) extracts of different cyanobacteria strains [
26], the lowest IC
50 value being 822.70 µg/mL for
Phormidium sp. LEGE 05292, while no activity was detected for
Nodosilinea nodulosa LEGE 06102. Lopes and co-workers reported O
2•− scavenging activity for
Nodosilinea (
Leptolynbbya)
antarctica LEGE13457 and
Cuspidothrix issatschenkoi LEGE 03282 acetone extracts, with IC
25 values of 319 and 286 µg/mL, and no activity for
Leptolynbbya-like sp. LEGE 13412 [
27]. The authors also reported a higher effectiveness in the acetone extracts when compared to ethanol extracts. Another study conducted by Amaro and co-workers revealed IC
50 values of 1394 and 826 µg/mL for
Gloeothece sp. and
Scenedesmus obliquus (M2-1), respectively [
37]. The results obtained herein for acetone extracts seem less promising than those previously reported, even though they are within the same order of magnitude. On the other hand, the aqueous extracts revealed an enormous potential worthy of further exploitation regarding cosmetic applications in the field of skin aging.
2.3.2. Enzymes Inhibition
MMPs are a family of extracellular zinc-dependent enzymes, whose main function is to remodel and degrade the ECM [
38], a gel-like material essential to hold cells together and provide a pathway for nutrients and oxygen [
39]. Alterations in the ECM components, such as collagen and elastin, induced by MMPs are the basis of skin damage and wrinkle formation [
40]. Together with these enzymes, linked with skin structure and wrinkle formation, another assumes a crucial role in the aging process due to its activity in melanogenesis: tyrosinase. Among other factors, UVR exposure causes an accumulation of an abnormal amount of melanin, due to increased ROS production. These reactive species affect the activity of melanocytes, which increases the conversion of tyrosine into melanin by oxidation, leading to hyperpigmentation and irregular skin patches [
41].
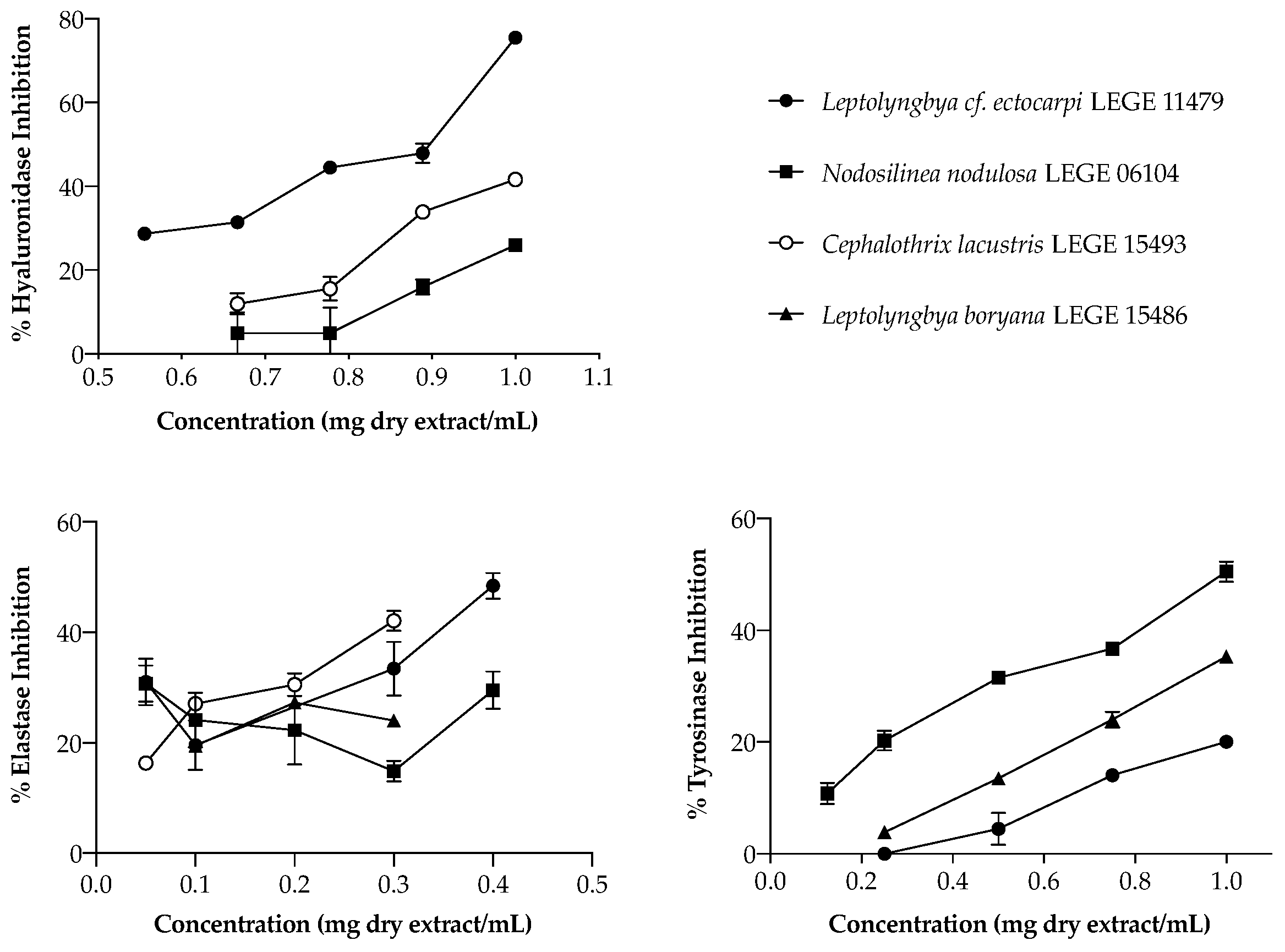
In the search for natural alternatives to commercial anti-aging ingredients, focusing the enzymes mentioned above, the activity of cyanobacteria extracts was explored (
Figure 2). Regarding HAase, only three strains were able to inhibit this enzyme, acting in a dose-dependent manner: the aqueous extract of
Leptolyngbya cf.
ectocarpi LEGE 11479 (IC
50 = 863 µg/mL), and the acetone extracts of
Cephalothrix lacustris LEGE 15493 and
Nodosilinea nodulosa LEGE 06104, the latter two only reaching IC
25 (832 and 995 µg/mL, respectively). Even though these values seem high, their range of activity was similar to that of the reference drug di-sodium cromoglicate (DSCG) (IC
50 = 1105 µg/mL). Moreover, it is worth mentioning that, for the highest concentration tested (1 mg/mL),
Leptolyngbya cf.
ectocarpi LEGE 11479 inhibited this enzyme in 80% (
Figure 2), which makes this extract promising as a cosmetic ingredient.
Few studies have reported on the potential effect of cyanobacteria compounds and extracts on hyaluronidase activity, and any comparison on extracts’ biological potential should take into account that cyanobacterial metabolism suffers significant variation depending on the cultivation conditions, influencing extracts’ chemical composition. Morone and co-workers [
26] evaluated the effect of ethanol extracts of
Tychonema sp. LEGE 07196 and
Cyanobium sp. LEGE 07175 on hyaluronidase, and found a stronger inhibitory activity, with IC
50 values of 182.74 and 208.36 µg/mL, respectively. The activity of ethanol extracts has also been reported for an insoluble fraction of
Spirulina platensis, with an IC
50 of 150 µg/mL [
42]. Another study, conducted by Yamaguchi and Koketsu [
19], showed that
Nostochopsis lobatus MAC0804NAN produced a large amount of polysaccharides with a high inhibitory effect (IC
50 = 7.18 µg/mL). It was also reported that an
Arthrospira-derived peptide may be involved in hyaluronidase inhibition [
43]. Together, these results support the potential of cyanobacteria compounds and extracts as anti-aging ingredients for cosmeceutical applications.
Considering elastase, only acetone extracts presented interesting bioactivity.
Leptolyngbya cf.
ectocarpi LEGE 11479 was again the most active (
Figure 2), being the only strain to reach the IC
50, with a value of 391 µg/mL.
Nodosilinea nodulosa LEGE 06104,
Cephalothrix lacustris LEGE 15493, and
Leptolyngbya boryana LEGE 15486 only reached IC
25, with values of 126, 86, and 99 µg/mL, respectively. As for hyaluronidase, marine strains have shown to be the most promising.
To the best of our knowledge, there are no previous reports on elastase inhibitory activity for the strains explored herein. Regarding other strains, it was discovered that
Nostoc minutum produced microviridins-type peptides and nostopeptins, with IC
50 = 1.3 and 11.0 µg/mL [
44,
45]. Microviridins B and C obtained from
Microcystis aeruginosa also inhibited elastase effectively, with IC
50 values of 0.044 and 0.084 µg/mL [
46]. Most of the available data regarding elastase inhibition focus on isolated compounds, so comparisons with the extracts from our strains are difficult to make.
Uneven skin pigmentation, associated with both aging and UV exposure, remains a major concern of aging populations and cosmetic industries. The majority of the available studies are trifling and use mushroom tyrosinase as an enzymatic model, making it difficult to translate the results to the human environment, nevertheless, this enzyme has a high similarity and homology with human tyrosinase [
47]. Thus, the same enzymatic model was used herein to explore the potential of cyanobacteria extracts in tyrosinase inhibition. As for elastase, only acetone extracts were able to inhibit tyrosinase.
Nodosilinea nodulosa LEGE 06104 was the most effective (
Figure 2), being the only strain to reach the IC
50 (989.26 ± 4.3 µg/mL). Below that was
Leptolyngbya boryana LEGE 15486, with IC
25 = 784. 78 ± 4.33 µg/mL, and lastly
Leptolyngbya cf.
ectocarpi LEGE 11479, with the most promising results being found for the marine strains.
Morone and co-workers [
26] previously explored the activity of cyanobacteria ethanolic extracts in the same model, but found no activity. One interesting aspect of this is that one of the studied strains was
Nodosilinea nodulosa LEGE 06102, which showed the best results herein, once again emphasizing the importance of the extraction solvents in obtaining targeted bioactive extracts. As for the other enzymes explored, surveys focusing cyanobacteria are also scarce for tyrosinase. In work conducted by Yabuta and his team [
48], it was reported that the hot water extract of
Nostochopsis spp. significantly inhibited the tyrosinase activity (IC
50 = 250 µg/mL). This is a very interesting result, considering that it results from an aqueous extract, for which no activity was found in the present study. The authors attribute the result to the low molecular weight compounds released from PBPs by heat treatment, namely a biline moiety that acts as a potent peroxyl radical scavenger. Another study evaluated the inhibitory activity of
Arthrospira platensis ethanol (IC
50 = 14,000 µg/mL) and water (IC
50 = 72,000 µg/mL) extracts, where the values were attributed to the presence of phenolic compounds such as ferulic and caffeic acids in the ethanol extract [
49].
2.3.3. UV Protection
Even though a growing number of cosmetic companies incorporate sun blockers into their products, it is still difficult to convince consumers about the benefits of their daily use to slow down premature skin aging. If on the one hand, the daily application of sun blockers is a little ingrained habit, on the other hand, there is a certain fear in the use of synthetic substances, due to their unwelcome associated risks [
50]. Following on from this, research on natural photoprotectors has considerably increased in the last few years, considering their potential for biodegradability and lower toxicity, making them more beneficial to humans and the environment. In order to explore cyanobacteria extracts in this field, their capacity to act as sunscreens was evaluated in vitro for UVR-B, since it is the most harmful radiation. The results found for cyanobacteria aqueous and acetone extracts are presented in
Table 7.
Regarding acetone extracts, the most promising value (19.2) was found for
Leptolyngbya boryana LEGE 15486, followed by
Leptolyngbya cf.
ectocarpi LEGE 11479 (10.7), both at the lowest concentration tested, 200 µg/mL.
Nodosilinea nodulosa LEGE 06104 was the least promising among acetone extracts. In aqueous extracts, the most promising results were obtained for the highest concentration tested, with
Leptolyngbya boryana LEGE 15486 and
Cephalothrix lacustris LEGE 15493 standing out with in vitro SPF values of 17.1 and 14.9, respectively (
Table 7). Discussing previous studies in this field, Hossain and his team [
51] reported that the SPF value for
Cephalothrix komarekiana extract was 2.37. Another group found that the SPF of a methanol extract of
Aphanizomenon flos-aquae was 4 [
52]. However, the information about the concentration of the extract used by the authors was not available, making possible comparisons difficult.
The number of cyanobacteria strains explored in the field of cosmetics, especially regarding SPF, is very scarce, considering the potentialities of these resources. The results presented herein demonstrate that the species under study may be good options as biological photoprotectors, and possibly act as boosters to other sunscreens currently marketed, allowing a reduction in the concentration of synthetic sunscreens in the formulas. Therefore, it is crucial to increase research on these organisms, especially their bioactive extracts, that, being of significantly lower cost, higher yield, and faster obtainment than isolated compounds are more environmentally friendly and economically more attractive.


 ,
, 






{kind=link}
{kind=link}
{kind=link}