
Emerging Marine Biotoxins in European Waters: Potential Risks and Analytical Challenges



Abstract
:1. Introduction
2. Emerging Marine Toxins in European Waters and Their Risks
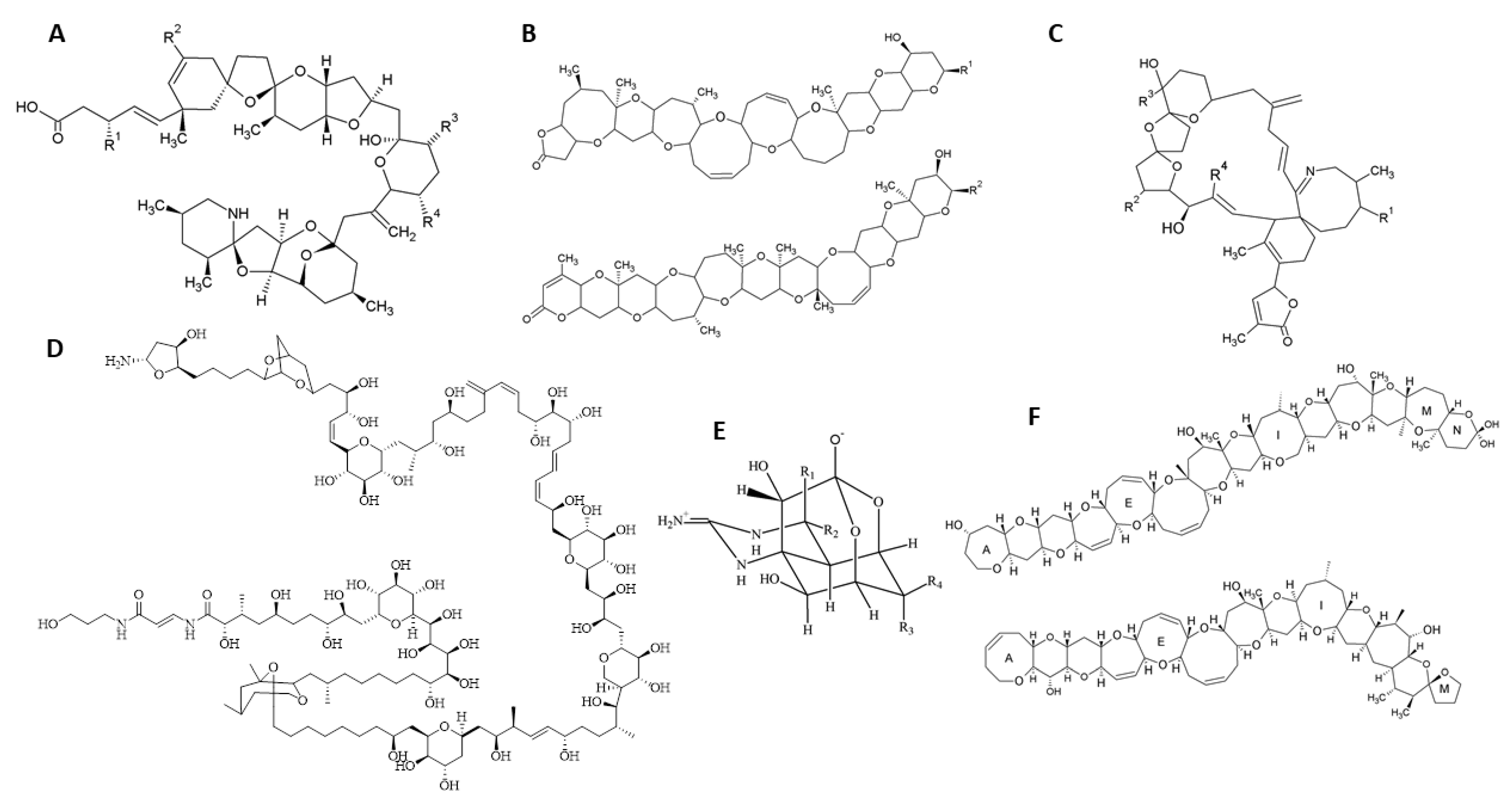
2.1. Imine Cyclic Toxins
2.1.1. Spirolides
2.1.2. Pinnatoxins
2.1.3. Gymnodimines
2.2. Azaspiracids
2.3. Palytoxins
2.4. Ciguatoxins

{kind=link}
{kind=link}
| Toxin | Report Location | Year | Vector/Uptake Route | Incident | No. Poisonings | Refs. |
|---|---|---|---|---|---|---|
| Imine Cyclic | ||||||
| PnTX-G | Ingril Lagoon (France) | 2010 | Mussels (Mytilus galloprovincialis) and clams (Venerupis decussata) | 1200 mg/kg of PnTX-G in mussels and clams | 0 | [60] |
| Azaspiracids | ||||||
| AZAs | Norway | 2005 | Viscera of the edible (brown) crab, C. pagurus, | Hospitalization of 2 persons after eating crabs containing AZA. | 2 | [7] |
| AZAs | Coast of Sweden | 2018 | Azadinium spp. | AZA levels above the regulatory limit | 0 | [7] |
| AZAs | North Sea coast, Netherlands, | 2020 | Phaeocystis globosa | Human fatalities: 5 persons playing water sports died. | 5 | [7] |
| Palytoxins | ||||||
| PlTXs | Genova (Italy) | 2005 | Ostreopsis sp./exposure to aerosols | Hospitalization of several hundred persons. | >100 | [99] |
| PlTX, ovatoxin-a | Ligurian Coasts | 2006 | O. ovata | Human toxic outbreak. Bathing was forbidden | Few cases | [100] |
| PlTXs | Almeria (Spain) | 2006 | Ostreopsis spp./exposure to aerosols | Epidemic outbreak with respiratory symptoms | >100 | [101] |
| PlTXs | French Mediterranean coast | 2006–2009 | 9 blooms Ostreopsis spp. | Respiratory irritation in 47 swimmers. Hospitalization of an 8-year-old girl (dyspnea). | 48 | [94] |
| Ciguatoxins | ||||||
| CTXs | Madeira archipelago, Portugal | 2007–2008 | No vectors were identified | Hospitalization of 6 persons exhibiting CP symptomatology | 6 | [124] |
| CTXs | Madeira archipelago, Portugal | 2008 | Amberkacl (Seriola spp.) | Hospitalization of 11 persons after consumption of the contaminated fish (CTX concentration NDA) | 11 | [124] |
| CTXs | Spain | 2012 | Amberjack (Seriola spp.) and Grouper (Epinephelus sp.) | Poisoning victims with symptoms consistent with CP, after ingestion of a predatory local fish; 12 intoxications were confirmed analytically for CTX. | 37 | [31] |
| CTXs | Portugal | 2012 | Amberjack and Barred Hogfish (Seriola sp. Bodianus scrofa) | Hospitalization of 12 poisoning victims, CTX NDA | 12 | [31] |
| CTXs | Spain | 2013 | Grouper (Epinephelus sp.) | Poisoning victims with symptoms consistent with CP, intoxications were confirmed analytically for CTX | 15 | [31] |
| CTXs | Spain | 2015 | Grouper (Epinephelus sp., Mycteroperca fusca) and Bluefish (Pomatomus saltatrix) | Poisoning victims with symptoms consistent with CP, 2 intoxications were confirmed analytically for CTX | 8 | [31] |
| CTXs | Portugal | 2015 | Grouper (Epinephelus marginatus) | Hospitalization of 4 out of 7 poisoning victims, CTX NDA | 7 | [31] |
| CTXs | Spain | 2016 | Grouper and Amberjack (Epinephelus sp. and Seriola sp.) | Poisoning victims with symptoms consistent with CP, intoxications were confirmed analytically for CTX | 5 | [31] |
| CTXs | Portugal | 2016 | Red Porgy (Pagrus pagrus) | Poisoning victims with symptoms consistent with CP, intoxications were confirmed analytically for CTX | 4 | [31] |
| CTXs | Spain | 2017 | Grouper (Epinephelus sp., Mycteroperca fusca) | Poisoning victims with symptoms consistent with CP, intoxications were confirmed analytically for CTX | 2 | [31] |
| CTXs | Spain | 2018 | Triggerfish (Canthidermis sufflame) | Hospitalization of 1 person. Poisoning victims with symptoms consistent with CP. CTX NDA | 4 | [31] |
| CTXs | Spain | 2019 | Amberkacl (Seriola spp.) | Poisoning victims with symptoms consistent with CP, intoxications were confirmed analytically for CTX | 6 | [31] |
| Tetrodotoxins | ||||||
| TTX and 5,6,11-trideoxyTTX | Spain | 2008 | Trumpet Shell (Charonia lampas) | Hospitalization of a person who ate a contaminated gastropod (315 mg TTX/kg) | 1 | [22] |
2.5. Brevetoxins
2.6. Tetrodotoxins
3. Challenges for the Detection of Emerging Toxin Detection
3.1. Cyclic Imines
3.2. Azaspiracids
3.3. Palytoxins
3.4. Ciguatoxins
3.5. Brevetoxins
3.6. Tetrodotoxins
| Equipment | Chromatographic Column | Mobile Phase | Toxins Tested | LOD/LOQ | Refs. |
|---|---|---|---|---|---|
| Imine Cyclic | |||||
| UPLC-MS/MS | Aquity UPLC BEH C18 (2.1 µM × 100 mm, 1.7 µm, Waters, Barcelona, Spain) | A = 100% water. B = acetonitrile:water (95:5), both containing 50 mM FA and 2 Mm AM. | PnTX-A,B,C,D,E,F,G GYM-A,B,C,D, 12-Me GYM-A. SPX-13, SPX-13,19 SPX-A,B,C,D,E,F,G,H,I. 20-MeSPX C. 27-OH-13,19-didesMe SPX-C. 27-OH-13-desMe SPX-C. | 0.1 µg/kg SPX-13, SPX-13,19, and PnTX-G. | [48] |
| LC-MS/MS | Agilent ZORBAX SB-octylsilyl (C8) (50 × 2.1 mm id, 1.8 μm). | A = 100% water B = acetonitrile:water (95:5), both with 2 mM AF and 50 mM FA. | PnTX-G. GYM-A. SPX-13 | LOD = 0.3 µg/kg and LOQ = 1 µg/kg. | [46] |
| Azaspiracids | |||||
| LC-MS/MS | Chromatographic column Acquity UPLC BEH C18 (2.1 × 100 mm, 1.7 µm, Waters). | A = 6.7 mM NH4OH (pH11). B = MeCN 90% with 6.7 mM NH4OH. | AZA-1,2,3,4,5,6; AZA-7,8,9,10,11,12; AZA-33,34,35,36,37,38; AZA-39,40,41,42,43; AZA-54,55,56,57,58; Me-AZA2; AZA2 phosphate; AZA11phosphate | LOQ = 42 mg AZA-2/kg meat. | [87] |
| Palytoxins | |||||
| UPLC-IT-TOF | HSS T3 column. Mobile phases. Temp: 35 °C | A = water. B = acetonitrile. Both acidified with 30 mM FA. | PLTX (m/z 906.81 and 1359.71) and 42-OH-PLTX (m/z 912.15 and 1367.72). | LOD = 190 ng/mL. LOQ = 650 ng/mL | [161] |
| LC-MS/LC-HRMS | Poroshell 120 EC-C18, 2.1 um × 100 mm. 25 ° C. | A = water. B = acetonitrile-water (95:5). Both containing 30 mM AA. | PLTX (m/z 906.8) and PLTX methyl ester (m/z 869.4). | LOD = 15 ng/mL | [190] |
| Ciguatoxins | |||||
| LC-MS/MS (detection and quantification) | Poroshell 120 EC-C18 column (3.0 × 50 mm, 2.7 µm, Agilent) | A = 0.1% FA and 5 mM AF. B = MeOH 0.1% FA and 5 mM AF | CTX-1B, C-CTX-1, 2,3-dihydroxiCTX-3C, 51-hydroxiCTX-3C, 52-epi-54-deoxyCTX-1B/54-deoxyCTX-1B, 49-epiCTX3C/CTX3C, CTX4A/CTX4B | 0.0045 μg/kg | [186] |
| LC-MS/MS (confirmatory for CTX-C) | Poroshell 120 EC-C18 column (3.0 × 50 mm, 2.7 µm, Agilent) | A = 0.1% FA and 5 mM AF. B = MeCN 0.1% FA and 5 mM AF | C-CTX-1 is based on three water losses and two confirmatory product ions m/z 191.1 108.9. | 0.0045 μg/kg | [186] |
| LRMS & HRMS (confirmatory) | C18 Kinetex column 50 × 2.1 mm, 1.7 μm, 100 Å (Phenomenex) | A = 2 mM AF B = 95% MeCN, 2 mM AF and 50 mM FA | P-CTX-3C and P-CTX-1B/Seafood and phytoplankton | P-CTX1B (0.075 μg/kg P-CTX3C (0.10 μg P-CTX1B eq./kg) | [185] |
| Brevetoxins | |||||
| LC-MS/MS | BDS Hypersil C8 (octylsilyl) HPLC column (3 µm, 50 × 2.1 mm, Thermo Scientific) | A = 50%/2.5% IA. B = 97.5% methanol/2.5% IA C = 30 mM AF and 470 mM FA D = 90% acetonitrile | BTX-B1, BTX-B2, S-deoxyBTX-B2, BTX-B5, BTX-2 and, BTX-3 | 0.025–0.048 mg/kg | [187] |
| LC-MS/MS | Kinetex XB-C18 (100 × 2.1 mm), 2.6 µm + pre-column Core-shell, 2.1 mm (Phenomenex) | A = 2 mM AF and 50 mM FA. B = MeOH/water (95:5, v/v), 2 mM AF and 50 mM FA | BTX-2, BTX-3 | 23 µg/kg | [132] |
| Tetrodotoxins | |||||
| HILIC-MS/MS | Waters Acquity UPLC Glycan BEH Amide HILIC Column, 130 Å 1.7 μm, 2.1 × 150 | A = 0.015% FA + 0.06% of 25% ammonia B = 70% MeCN + 0.01% FA. | TTX, 4-epi-TTX, 5,6,11-trideoxy TTX; 11-nor TTX-6-ol; 4,9-anhydro TTX; 5-deoxy TTX/11-deoxy TTX; 6,11-dideoxy-TTX | 0.31 ± 0.12 µg/kg | [189] |
| UPLC-MS/MS | ACQUITY UPLC BEH Amide (2.1 × 100 mm, 1.7 µm, Waters) | A = 0.1% FA and 10 mM AF. B = 95% MeCN 0.1% FA and 2% 100 mM AF | TTX, 4-epi-TTX; 5,6,11-trideoxy TTX; 11-nor TTX-6-ol; 5-deoxy TTX; and 4,9-anhydro TTX | 0.25 µg/kg | [188] |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Otero, P.; Alfonso, A.; Alfonso, C.; Vieytes, M.R.; Louzao, M.C.; Botana, A.M.; Botana, L.M. New protocol to obtain spirolides from Alexandrium ostenfeldii cultures with high recovery and purity. Biomed. Chromatogr. 2010, 24, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Griffith, A.W.; Gobler, C.J. Harmful algal blooms: A climate change co-stressor in marine and freshwater ecosystems. Harmful Algae 2020, 91, 101590. [Google Scholar] [CrossRef] [PubMed]
- Cembella, A.D. Chemical ecology of eukaryotic microalgae in marine ecosystems. Phycologia 2003, 42, 420–447. [Google Scholar] [CrossRef]
- Townhill, B.L.; Tinker, J.; Jones, M.; Pitois, S.; Creach, V.; Simpson, S.D.; Dye, S.; Bear, E.; Pinnegar, J.K. Harmful algal blooms and climate change: Exploring future distribution changes. ICES J. Mar. Sci. 2018, 75, 1882–1893. [Google Scholar] [CrossRef] [Green Version]
- Vilariño, N.; Louzao, M.C.; Cagide, E.; Carrera, C.; Abal, P.; Vieytes, M.R.; Botana, L.M. Human Poisoning from Marine Toxins: Unknowns for Optimal Consumer Protection. Toxins 2018, 10, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otero, P.; Miguens, N.; Rodriguez, I.; Botana, L.M. toxins Pinnatoxin-G and High Levels of Esterified OA Group. Toxins 2019, 11, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlson, B.; Andersen, P.; Arneborg, L.; Cembella, A.; Eikrem, W.; John, U.; Joy, J.; Klemm, K.; Kobos, J.; Lehtinen, S.; et al. Harmful algal blooms and their effects in coastal seas of Northern Europe. Harmful Algae 2021, 102, 101989. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Regulation (EU) No 853/2004 of the European Parliament and of the Council of 29 April 2004 Laying Down Specific Hygiene Rules for Food of Animal Origin; European Commission: Brussels, Belgium, 2004; pp. 55–205.
- European Commission. Regulation (EU) No 786/2013 of 16 August 2013 Amending Annex III to Regulation (EC) No 853/2004 of the European Parliament and of the Council as Regards the Permitted Limits of Yessotoxins in Live Bivalve Molluscs; European Commission: Brussels, Belgium, 2013; p. 14.
- European Commission. Commision Delegated Regulation (EU) 2021/1374 of 12 April 2021 Amending Annex III to Regulation (EC) No 853/2004 of the European Parliament and of the Council on Specific Hygiene Requirements for food of Animal Origin; European Commission: Brussels, Belgium, 2021; pp. 1–15.
- Alexander, J.; Benford, D.; Cockburn, A.; Cravedi, J.P.; Dogliotti, E.; Di Domenico, A.; Fernández-Cruz, M.L.; Fink-Gremmels, J.; Fürst, P.; Galli, C.; et al. Marine biotoxins in shellfish—Azaspiracid group Scientific Opinion of the Panel on Contaminants in the Food chain. EFSA J. 2008, 723, 1–52. [Google Scholar]
- European Commission. Commission Implementing Regulation (EU) 2021/1709 of 23 September 2021 Amending Implementing Regulation (EU) 2019/627 as Regards Uniform Practical Arrangements for the Performance of Official Controls on Products of Animal Origin; European Commission: Brussels, Belgium, 2021; pp. 84–88.
- Boundy, M.J.; Harwood, D.T.; Tommasi, E.; Burger, E.; van Ginkel, R.; Waugh, C.; Selwood, A.I.; Finch, S. Acute toxicity of decarbamoyl gonyautoxin 1 & 4 to mice by various routes of administration. Toxicon 2021, 204, 56–63. [Google Scholar] [CrossRef]
- Alexander, J.; Benford, D.; Cockburn, A.; Cravedi, J.; Dogliotti, E.; di Domenico, A.; Fernández-Cruz, M.L.; Fink-Gremmels, J.; Fürst, P.; Galli, C.; et al. Marine biotoxins in shellfish—Saxitoxin group Scientific Opinion of the Panel on Contaminants in the Food Chain. EFSA J. 2009, 1181, 1019. [Google Scholar] [CrossRef]
- Alexander, J.; Benford, D.; Boobis, A.; Ceccatelli, S.; Cravedi, J.; di Domenico, A.; Doerge, D.; Dogliotti, E.; Edler, L.; Farmer, P.; et al. Marine biotoxins in shellfish—Domoic acid Scientific Opinion of the Panel on Contaminants in the Food Chain Adopted on 2 July 2009. EFSA J. 2009, 1181, 1–61. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EU) No 15/2011 of 10 January 2011 Amending Regulation (EC) No 2074/2005 as Regards Recognised Testing Methods for Detecting Marine Biotoxins in Live Bivalve Molluscs; European Commission: Brussels, Belgium, 2011; pp. 22–25.
- Lawrence, J.F.; Niedzwiadek, B. Quantitative determination of paralytic shellfish poisoning toxins in shellfish by using prechromatographic oxidation and liquid chromatography with fluorescence detection. J. AOAC Int. 2001, 84, 1099–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reference, E.U. Laboratory for Marine EU-Harmonised Standard Operating Procedure for Determination of Lipophilic Marine Biotoxins in Molluscs by LC-MS/MS. 2015. Available online: http://www.aesan.gob.es/AECOSAN/docs/documentos/laboratorios/LNRBM/ARCHIVO2EU-Harmonised-SOP-LIPO-LCMSMS_Version5.pdf (accessed on 15 January 2022).
- European Commission. Commission Decision of 12 August 2002 Implementing Council Directive 96/23/EC Concerning the Performance of Analytical Methods and the Interpretation of Results; European Commission: Brussels, Belgium, 2002; pp. 8–36.
- Estevez, P.; Castro, D.; Pequeño-valtierra, A.; Giraldez, J.; Gago-martinez, A. Emerging Marine Biotoxins in Seafood from European Coasts: Incidence and Analytical Challenges. Foods 2019, 8, 149. [Google Scholar] [CrossRef] [Green Version]
- Mudge, E.M.; Miles, C.O.; Hardstaff, W.R.; Mccarron, P. Toxicon: X Fatty acid esters of azaspiracids identified in mussels (Mytilus edulis) using liquid chromatography-high resolution mass spectrometry. Toxicon X 2020, 8, 100059. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, P.; Alfonso, A.; Vale, C.; Alfonso, C.; Vale, P.; Tellez, A.; Botana, L.M. First toxicity report of tetrodotoxin and 5,6,11-trideoxyTTX in the trumpet shell Charonia lampas lampas in Europe. Anal. Chem. 2008, 80, 5622–5629. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Azevedo, J.; Rodriguez, P.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. New gastropod vectors and tetrodotoxin potential expansion in temperate waters of the Atlantic Ocean. Mar. Drugs 2012, 10, 712–726. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.D.; Higgins, C.; Higman, W.; Hungerford, J. Potential threats posed by tetrodotoxins in UK waters: Examination of detection methodology used in their control. Mar. Drugs 2015, 13, 7357–7376. [Google Scholar] [CrossRef] [Green Version]
- Gerssen, A.; Bovee, T.H.F.; Klijnstra, M.D.; Poelman, M.; Portier, L.; Hoogenboom, R.L.A.P. First report on the occurrence of tetrodotoxins in bivalve molluscs in the Netherlands. Toxins 2018, 10, 450. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.; Rodríguez, I.; Barreiro, A.; Kaufmann, M.; Neto, A.I.; Hassouani, M.; Sabour, B.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. Tetrodotoxins occurrence in non-traditional vectors of the north atlantic waters (Portuguese maritime territory, and morocco coast). Toxins 2019, 11, 306. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.D.; Higgins, C.; Davidson, K.; Veszelovszki, A.; Payne, D.; Hungerford, J.; Higman, W. Potential threats posed by new or emerging marine biotoxins in UK waters and examination of detection methodology used in their control: Brevetoxins. Mar. Drugs 2015, 13, 1224–1254. [Google Scholar] [CrossRef] [Green Version]
- Murrell, R.N.; Gibson, J.E. Brevetoxin 2 alters expression of apoptotic, DNA damage, and cytokine genes in Jurkat cells. Hum. Exp. Toxicol. 2011, 30, 182–191. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel. Scientific Opinion on marine biotoxins in shellfish—Cyclic imines (spirolides, gymnodimines, pinnatoxins and pteriatoxins). EFSA J. 2010, 8, 1628. [Google Scholar] [CrossRef]
- Chain, E. Scientific Opinion on marine biotoxins in shellfish—Palytoxin group. EFSA J. 2009, 7, 1393. [Google Scholar] [CrossRef]
- Varela Martínez, C.; León Gómez, I.; Martínez Sánchez, E.V.; Carmona Alférez, R.; Nuñez Gallo, D.; Friedemann, M.; Oleastro, M.; Boziaris, I. Incidence and epidemiological characteristics of ciguatera cases in Europe. EFSA Support. Publ. 2021, 18, 6650E. [Google Scholar] [CrossRef]
- EFSA Panel. Scientific Opinion on marine biotoxins in shellfish—Emerging toxins: Brevetoxin group. EFSA J. 2010, 8, 1677. [Google Scholar] [CrossRef]
- EFSA Panel. Risks for public health related to the presence of tetrodotoxin (TTX) and TTX analogues in marine bivalves and gastropods. EFSA J. 2017, 15, 4752. [Google Scholar] [CrossRef] [Green Version]
- Katikou, P. Digital technologies and open data sources in marine biotoxins’ risk analysis: The case of ciguatera fish poisoning. Toxins 2021, 13, 692. [Google Scholar] [CrossRef]
- O’Neill, A.; Morrell, N.; Turner, A.D.; Maskrey, B.H. Method performance verification for the combined detection and quantitation of the marine neurotoxins cyclic imines and brevetoxin shellfish metabolites in mussels (Mytilus edulis) and oysters (Crassostrea gigas) by UHPLC-MS/MS. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2021, 1179, 122864. [Google Scholar] [CrossRef]
- Vieira, A.C.; Rubiolo, J.A.; Alfonso, A.; Otero, P.; Mercedes, R.; Botana, L.M. Oral toxicity of Okadaic acid in mice: Study of lethality, organ damage, distribution and effects on detoxifying gene expression. Toxins 2013, 5, 2093–2108. [Google Scholar] [CrossRef] [Green Version]
- Botana, A.M.; Otero, P.; Rodriguez, P.; Alfonso, A.; Botana, L.M. Current situation on analysis of marine toxins. Rev. Anal. Chem. 2013, 32, 15–34. [Google Scholar] [CrossRef]
- Otero, P.; Alfonso, A.; Alfonso, C.; Rodríguez, P.; Vieytes, M.R.; Botana, L.M. Chromatography À Tandem Mass Spectrometry Method Major Causes for Concern. Anal. Chem. 2011, 83, 5903–5911. [Google Scholar] [CrossRef] [PubMed]
- Otero, P.; Alfonso, A.; Rodríguez, P.; Rubiolo, J.A.; Manuel, J.; Bermúdez, R.; Vieytes, M.R.; Botana, L.M. Pharmacokinetic and toxicological data of spirolides after oral and intraperitoneal administration. FOOD Chem. Toxicol. 2012, 50, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Otero, P.; Alfonso, A.; Vieytes, M.R.; Cabado, A.; Vieites, J.M.; Botana, L.M. Effects of Environmental Regimens on the Toxin profile of Alexandrium ostenfeldii. Environ. Toxicol. Chem. 2010, 29, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Aasen, J.; MacKinnon, S.L.; LeBlanc, P.; Walter, J.A.; Hovgaard, P.; Aune, T.; Quilliam, M.A. Detection and identification of spirolides in Norwegian shellfish and plankton. Chem. Res. Toxicol. 2005, 18, 509–515. [Google Scholar] [CrossRef]
- Fonfría, E.S.; Vilariño, N.; Molgó, J.; Aráoz, R.; Otero, P.; Espiña, B.; Louzao, M.C.; Alvarez, M.; Botana, L.M. Detection of 13, 19-didesmethyl C spirolide by fluorescence polarization using Torpedo electrocyte membranes. Anal. Biochem. 2010, 403, 102–107. [Google Scholar] [CrossRef]
- Franco, M.; Bravo, I.; Salgado, P.; Riob, P. Toxicon Differences in the toxin pro fi les of Alexandrium ostenfeldii (Dinophyceae) strains isolated from different geographic origins: Evidence of paralytic toxin, spirolide, and gymnodimine. Toxicon 2015, 103, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Davidson, K.; Baker, C.; Higgins, C.; Higman, W.; Swan, S.; Veszelovszki, A.; Turner, A.D. Potential threats posed by new or emerging marine biotoxins in ukwaters and examination of detection methodologies used for their control: Cyclic imines. Mar. Drugs 2015, 13, 7087–7112. [Google Scholar] [CrossRef]
- Guinder, V.A.; Tillmann, U.; Krock, B.; Delgado, A.L.; Krohn, T.; Cardona, J.E.G.; Metfies, K.; Abbate, C.L.; Silva, R.; Lara, R. Plankton multiproxy analyses in the Northern Patagonian Shelf, Argentina: Community structure, phycotoxins, and characterization of toxic Alexandrium strains. Front. Mar. Sci. 2018, 5, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Moreiras, G.; Le, M. Analysis of Cyclic Imines in Mussels (Mytilus galloprovincialis) from Galicia (NW Spain) by LC-MS/MS. Int. J. Environ. Res. Public Health 2019, 17, 281. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.; Barreiro, A.; Rodriguez, P.; Otero, P.; Azevedo, J.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. New Invertebrate Vectors for PST, Spirolides and Okadaic Acid in the North Atlantic. Mar. Drugs 2013, 11, 1936–1960. [Google Scholar] [CrossRef] [Green Version]
- Otero, P.; Vale, C.; Boente-Juncal, A.; Costas, C.; Carmen Louzao, M.; Botana, L.M. Detection of cyclic imine toxins in dietary supplements of green lipped mussels (Perna canaliculus) and in shellfish mytilus chilensis. Toxins 2020, 12, 613. [Google Scholar] [CrossRef] [PubMed]
- Nieva, J.A.; Tebben, J.; Tillmann, U.; Wohlrab, S.; Krock, B. Mass Spectrometry-based characterization of new spirolides from Alexandrium ostenfeldii (Dinophyceae). Mar. Drugs 2020, 18, 505. [Google Scholar] [CrossRef] [PubMed]
- Otero, P.; Alfonso, A.; Alfonso, C.; Aráoz, R.; Molgó, J.; Vieytes, M.R.; Botana, L.M. Analytica Chimica Acta First direct fluorescence polarization assay for the detection and quantification of spirolides in mussel samples. Anal. Chim. Acta 2011, 701, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Nagatsu, A.; Fukuzawa, S.; Zheng, S.; Chen, H.; Data, N.M.R. Pinnatoxin A: A toxic amphoteric macrocycle from the Okinawan bivalve Pinna muricata. J. Am. Chem. Soc. 1995, 117, 1155–1156. [Google Scholar]
- Selwood, A.I.; Miles, C.O.; Wilkins, A.L.; Ginkel, R.V.; Munday, R.; Rise, F.; McNabb, P. Isolation, Structural Determination and Acute Toxicity of Pinnatoxins E, F and G. J. Agric. Food Chem. 2010, 58, 6532–6542. [Google Scholar] [CrossRef]
- Mcnabb, P.S.; Mccoubrey, D.J.; Rhodes, L.; Smith, K.; Selwood, A.I.; Van Ginkel, R.; Mackenzie, A.L.; Munday, R.; Holland, P.T. New perspectives on biotoxin detection in Rangaunu Harbour, New Zealand arising from the discovery of pinnatoxins. Harmful Algae 2012, 13, 34–39. [Google Scholar] [CrossRef]
- Aráoz, R.; Barnes, P.; Sechet, V.; Delepierre, M.; Zinn-justin, S.; Molgó, J.; Zakarian, A.; Hess, P.; Servent, D. Cyclic imine toxins survey in coastal European shellfish samples: Bioaccumulation and Mode of Action of 28-O-palmitoyl ester of pinnatoxin-G. First Report of portimine-A bioaccumulation. Harmful Algae 2020, 9898, 101887. [Google Scholar] [CrossRef]
- Rundberget, T.; Bunæs, J.A.; Selwood, A.I.; Miles, C.O. Toxicon Pinnatoxins and spirolides in Norwegian blue mussels and seawater. Toxicon 2011, 58, 700–711. [Google Scholar] [CrossRef]
- Guillotin, S.; Delcourt, N. Marine Neurotoxins’ Effects on Environmental and Human Health: An OMICS Overview. Mar. Drugs 2022, 20, 18. [Google Scholar] [CrossRef]
- Lamas, J.P.; Ar, F.; Moroño, Á.; Correa, J.; Muñíz, S.; Blanco, J. Detection and Spatio-Temporal Distribution of Pinnatoxins in Shellfish from the Atlantic and Cantabrian Coasts of Spain. Toxins 2019, 11, 340. [Google Scholar] [CrossRef] [Green Version]
- Moreira-González, A.R.; Comas-González, A.; Valle-Pombrol, A.; Seisdedo-Losa, M.; Hernández-Leyva, O.; Fernandes, L.F.; Chomérat, N.; Bilien, G.; Hervé, F.; Rovillon, G.A.; et al. Summer bloom of Vulcanodinium rugosum in Cienfuegos Bay (Cuba) associated to dermatitis in swimmers. Sci. Total Environ. 2021, 757, 143782. [Google Scholar] [CrossRef] [PubMed]
- Rambla-Alegre, M.; Miles, C.O.; De, P.; Fernandez-tejedor, M.; Jacobs, S.; Sioen, I.; Verbeke, W.; Samdal, I.A.; Sandvik, M.; Barbosa, V.; et al. Occurrence of cyclic imines in European commercial seafood and consumers risk assessment. Environ. Res. 2018, 161, 392–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hess, P.; Abadie, E.; Hervé, F.; Berteaux, T.; Séchet, V.; Aráoz, R.; Molgó, J.; Zakarian, A.; Sibat, M.; Rundberget, T.; et al. Pinnatoxin G is responsible for atypical toxicity in mussels (Mytilus galloprovincialis) and clams (Venerupis decussata) from Ingril, a French Mediterranean lagoon. Toxicon 2013, 75, 16–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnich, N.; Abadie, E.; Delcourt, N.; Fessard, V.; Fremy, J.M.; Hort, V.; Lagrange, E.; Maignien, T.; Molgó, J.; Peyrat, M.B.; et al. Health risk assessment related to pinnatoxins in French shellfish. Toxicon 2020, 180, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sosa, S.; Pelin, M.; Cavion, F.; Herv, F.; Hess, P.; Tubaro, A. Acute Oral Toxicity of Pinnatoxin G in Mice. Toxins 2020, 12, 87. [Google Scholar] [CrossRef] [Green Version]
- Farabegoli, F.; Blanco, L.; Rodríguez, L.P.; Vieites, J.M.; Cabado, A.G. Phycotoxins in Marine Shellfish: Origin, Occurrence and Effects on Humans. Mar. Drugs 2018, 16, 188. [Google Scholar] [CrossRef] [Green Version]
- Miles, C.O.; Wilkins, A.L.; Stirling, D.J.; Mackenzie, A.L. New Analogue of Gymnodimine from a Gymnodinium Species. J Agric Food Chem. 2000, 48, 1373–1376. [Google Scholar] [CrossRef]
- Miles, C.O.; Wilkins, A.L.; Stirling, D.J.; MacKenzie, A.L. Gymnodimine C, an Isomer of Gymnodimine B, from Karenia selliformis. J. Agric. Food Chem. 2003, 51, 4838–4840. [Google Scholar] [CrossRef]
- Bacchiocchi, S.; Siracusa, M.; Campacci, D.; Ciriaci, M.; Dubbini, A.; Tavoloni, T.; Stramenga, A.; Gorbi, S.; Piersanti, A. Cyclic Imines (CIs) in Mussels from North-Central Adriatic Sea: First Evidence of Gymnodimine A in Italy. Toxins 2020, 12, 370. [Google Scholar] [CrossRef]
- Van de Waal, D.B.; Tillmann, U.; Martens, H.; Krock, B.; van Scheppingen, Y.; John, U. Characterization of multiple isolates from an Alexandrium ostenfeldii bloom in The Netherlands. Harmful Algae 2015, 49, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Krock, B.; Pitcher, G.C.; Ntuli, J.; Cembella, A.D. Confirmed identification of gymnodimine in oysters from the west coast of South Africa by liquid chromatography–tandem mass spectrometry. Afr. J. Mar. Sci. 2009, 31, 113–118. [Google Scholar] [CrossRef]
- Correa, J.; Rossignoli, A.E.; Blanco, J. Gymnodimine A in molluscs from the north Atlantic Coast of Spain: Prevalence, concentration, and relationship with spirolides. Environ. Pollut. 2021, 279, 116919. [Google Scholar] [CrossRef]
- Giuliani, M.E.; Accoroni, S.; Mezzelani, M.; Lugarini, F.; Bacchiocchi, S.; Siracusa, M.; Tavoloni, T.; Piersanti, A.; Totti, C.; Regoli, F.; et al. Biological E ff ects of the Azaspiracid-Producing Dinoflagellate Azadinium dexteroporum in Mytilus galloprovincialis from the Mediterranean Sea. Mar. Drugs 2019, 17, 595. [Google Scholar] [CrossRef] [Green Version]
- Ofuji, K.; Satake, M.; McMahon, T.; Silke, J.; James, K.J.; Naoki, H.; Oshima, Y.; Yasumoto, T. Two analogs of azaspiracid isolated from mussels, Mytilus edulis, involved in human intoxication in Ireland. Nat. Toxins 1999, 7, 99–102. [Google Scholar] [CrossRef]
- Ito, E.; Satake, M.; Ofuji, K.; Kurita, N.; Mcmahon, T.; James, K.; Yasumoto, T. Multiple organ damage caused by a new toxin azaspiracid, isolated from mussels produced in Ireland. Toxicon 2000, 38, 917–930. [Google Scholar] [CrossRef]
- Leyva, V.; Maruenda, H. Assessment of the Presence of Lipophilic Phycotoxins in Scallops (Argopecten purpuratus) Farmed along Peruvian Coastal Waters. J. Food Prot. 2021, 84, 204–212. [Google Scholar] [CrossRef]
- Rossi, R.; Aversano, C.D.; Krock, B.; Ciminiello, P.; Percopo, I.; Tillmann, U.; Soprano, V.; Zingone, A. Mediterranean Azadinium dexteroporum (Dinophyceae) produces six novel azaspiracids and azaspiracid-35: A structural study by a multi-platform mass spectrometry approach. Anal. Bioanal. Chem. 2017, 1121–1134. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Qiu, J.; Xie, T.; Mccarron, P.; Li, A. Toxicon Accumulation and transformation of azaspiracids in scallops (Chlamys farreri) and mussels (Mytilus galloprovincialis) fed with Azadinium poporum, and response of antioxidant enzymes. Toxicon 2018, 143, 20–28. [Google Scholar] [CrossRef]
- Aasen, J.A.B.; Espenes, A.; Hess, P.; Aune, T. Toxicon Sub-lethal dosing of azaspiracid-1 in female NMRI mice. Toxicon 2010, 56, 1419–1425. [Google Scholar] [CrossRef]
- Ferreiro, S.F.; Vilariño, N.; Carrera, C.; Carmen Louzao, M.; Cantalapiedra, A.G.; Santamarina, G.; Manuel Cifuentes, J.; Vieira, A.C.; Botana, L.M. Subacute cardiovascular toxicity of the marine phycotoxin azaspiracid-1 in rats. Toxicol. Sci. 2016, 151, 104–114. [Google Scholar] [CrossRef]
- Pelin, M.; Kilcoyne, J.; Nulty, C.; Hess, P.; Tubaro, A.; Sosa, S. Toxic equivalency factors (TEFs) after acute oral exposure of azaspiracid 1, -2 and -3 in mice. Toxicol. Lett. 2018, 282, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Twiner, M. Azaspiracid Shellfish Poisoning: A Review on the Chemistry, Ecology, and Toxicology with an Emphasis on Human Health Impacts. Mar. Drugs 2008, 6, 39–72. [Google Scholar] [CrossRef] [PubMed]
- Aune, T.; Espenes, A.; Aasen, J.A.B.; Quilliam, M.A.; Hess, P.; Larsen, S. Study of possible combined toxic effects of azaspiracid-1 and okadaic acid in mice via the oral route. Toxicon 2012, 60, 895–906. [Google Scholar] [CrossRef]
- Tillmann, U.; Potvin, E.; Jeong, H.J.; Kilcoyne, J. Structure Elucidation and in Vitro Toxicity of New Azaspiracids Isolated from the Marine Dinoflagellate Azadinium poporum. Mar. Drugs 2015, 13, 6687–6702. [Google Scholar] [CrossRef] [Green Version]
- Kilcoyne, J.; McCarron, P.; Twiner, M.J.; Rise, F.; Hess, P.; Wilkins, A.L.; Miles, C.O. Identification of 21,22-Dehydroazaspiracids in Mussels (Mytilus edulis) and in Vitro Toxicity of Azaspiracid-26. J. Nat. Prod. 2018, 81, 885–8893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elgarch, A.; Vale, P.; Rifai, S.; Fassouane, A. Detection of Diarrheic Shellfish Poisoning and Azaspiracid Toxins in Moroccan Mussels: Comparison of the LC-MS Method with the Commercial Immunoassay Kit. Mar. Drugs 2008, 587–594. [Google Scholar] [CrossRef]
- Callaghan, K.O.; Moriarty, M.; Driscoll, D.O.; Hamilton, B.; Lehane, M.; Lo, A.; James, K.J.; Furey, A. First evidence of azaspiracids (AZAs): A family of lipophilic polyether marine toxins in scallops (Argopecten purpuratus) and mussels (Mytilus chilensis) collected in two regions of Chile. Toxicon 2010, 55, 692–701. [Google Scholar] [CrossRef]
- Bacchiocchi, S.; Siracusa, M.; Ruzzi, A.; Gorbi, S.; Ercolessi, M.; Anna, M.; Ammazzalorso, P.; Orletti, R. Toxicon Two-year study of lipophilic marine toxin profile in mussels of the North-central Adriatic Sea: First report of azaspiracids in Mediterranean seafood. Toxicon 2015, 108, 115–125. [Google Scholar] [CrossRef]
- Dhanji-rapkova, M.; Neill, A.O.; Maskrey, B.H.; Coates, L.; Swan, S.C.; Teixeira, M.; Kelly, R.J.; Hat, R.G.; Rowland-pilgrim, S.J.; Lewis, A.M.; et al. Variability and profiles of lipophilic toxins in bivalves from Great Britain during five and a half years of monitoring: Azaspiracids and yessotoxins. Harmful Algae. 2019, 87, 101629. [Google Scholar] [CrossRef]
- Correa, J.; Mu, S.; Martín, H.; Mari, C. Toxicon Presence of azaspiracids in bivalve molluscs from Northern Spain. Toxicon 2017, 137, 135–143. [Google Scholar] [CrossRef]
- Vale, P.; Bire, R.; Hess, P. Confirmation by LC-MS/MS of azaspiracids in shellfish from the Portuguese north-western coast. Toxicon 2008, 51, 1449–1456. [Google Scholar] [CrossRef] [PubMed]
- Alfonso, C.; Rehmann, N.; Hess, P.; Alfonso, A.; Wandscheer, C.B.; Vale, C.; Otero, P.; Vieytes, M.R.; Botana, L.M. Evaluation of Various pH and Temperature Conditions on the Stability of Azaspiracids and Their Importance in Preparative Isolation and Toxicological Studies. Anal. Chem. 2008, 80, 9672–9680. [Google Scholar] [CrossRef] [PubMed]
- Aasen, J.A.B.; Espenes, A.; Miles, C.O.; Samdal, I.A.; Hess, P.; Aune, T. Combined oral toxicity of azaspiracid-1 and yessotoxin in female NMRI Mice. Toxicon 2011, 57, 909–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyva-Valencia, I.; Hernández-Castro, J.E.; Band-Schmidt, C.J.; Turner, A.D.; O’Neill, A.; Núñez-Vázquez, E.J.; López-Cortés, D.J.; Bustillos-Guzmán, J.J.; Hernández-Sandoval, F.E. Lipophilic Toxins in Wild Bivalves from the Southern Gulf of California, Mexico. Mar. Drugs 2021, 19, 99. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Rodríguez, I.; Barreiro, A.; Kaufmann, M.; Isabel, A.; Hassouani, M.; Sabour, B.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. Lipophilic toxins occurrence in non-traditional invertebrate vectors from North Atlantic Waters (Azores, Madeira, and Morocco): Update on geographical tendencies and new challenges for monitoring routines. Mar. Pollut. Bull. 2020, 161, 111725. [Google Scholar] [CrossRef]
- Patocka, J.; Gupta, R.C.; Wu, Q.H.; Kuca, K. Toxic potential of palytoxin. J. Huazhong Univ. Sci. Technol. Med. Sci. 2015, 35, 773–780. [Google Scholar] [CrossRef]
- Tichadou, L.; Glaizal, M.; Armengaud, A.; Grossel, H.; Lemée, R.; Kantin, R.; Lasalle, J.L.; Drouet, G.; Rambaud, L.; Malfait, P.; et al. Health impact of unicellular algae of the Ostreopsis genus blooms in the Mediterranean Sea: Experience of the French Mediterranean coast surveillance network from 2006 to 2009. Clin. Toxicol. 2010, 48, 839–844. [Google Scholar] [CrossRef] [Green Version]
- Ajani, P.; Harwood, D.T.; Murray, A. Recent trends in marine phycotoxins from Australian coastal waters. Mar. Drugs 2017, 15, 33. [Google Scholar] [CrossRef] [Green Version]
- Patocka, J.; Wu, O.; Kuca, K. Palytoxin. In Handbook of Foodborne Diseases; Liu, D., Ed.; CRC Press: Boca Raton, CA, USA, 2018; pp. 1035–1040. [Google Scholar]
- Ramos, V.; Vasconcelos, V. Palytoxin and analogs: Biological and ecological effects. Mar. Drugs 2010, 8, 2021–2037. [Google Scholar] [CrossRef] [Green Version]
- Tibiriçá, C.E.J.A.; Leite, I.P.; Batista, T.V.V.; Fernandes, L.F.; Chomérat, N.; Herve, F.; Hess, P.; Mafra, L.L. Ostreopsis cf. Ovata bloom in currais, Brazil: Phylogeny, toxin profile and contamination of mussels and marine plastic litter. Toxins 2019, 11, 446. [Google Scholar] [CrossRef] [Green Version]
- Pfannkuchen, M.; Godrijan, J.; Marić Pfannkuchen, D.; Iveša, L.; Kružić, P.; Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; et al. Toxin-producing ostreopsis cf. ovata are likely to bloom undetected along Coastal Areas. Environ. Sci. Technol. 2012, 46, 5574–5582. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Grillo, C.; Melchiorre, N. Putative Palytoxin and Its New Analogue, Ovatoxin-a, in Ostreopsis ovata Collected Along the Ligurian Coasts During the 2006 Toxic Outbreak. J. Am. Soc. Mass Spectrom. 2008, 19, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barroso García, P.; de la Puerta, P.R.; Parrón Carreño, T.; Marín Martínez, P.; Guillén Enríquez, J. Brote con síntomas respiratorios en la provincia de Almería por una posible exposición a microalgas tóxicas. Gac. Sanit. 2008, 22, 578–584. [Google Scholar] [CrossRef] [Green Version]
- Hall, C.; Levy, D.; Sattler, S. A Case of Palytoxin Poisoning in a Home Aquarium Enthusiast and His Family. Case Rep. Emerg. Med. 2015, 2015, 621815. [Google Scholar] [CrossRef]
- Biré, R.; Trotereau, S.; Lemée, R.; Delpont, C.; Chabot, B.; Aumond, Y.; Krys, S. Occurrence of palytoxins in marine organisms from different trophic levels of the French Mediterranean coast harvested in 2009. Harmful Algae 2013, 28, 10–22. [Google Scholar] [CrossRef]
- Reverté, L.; Soliño, L.; Carnicer, O.; Diogène, J.; Campàs, M. Alternative methods for the detection of emerging marine toxins: Biosensors, biochemical assays and cell-based assays. Mar. Drugs 2014, 12, 5719–5763. [Google Scholar] [CrossRef] [Green Version]
- Del Favero, G.; Beltramo, D.; Sciancalepore, M.; Lorenzon, P.; Coslovich, T.; Poli, M.; Testai, E.; Sosa, S.; Tubaro, A. Toxicity of palytoxin after repeated oral exposure in mice and invitro effects on cardiomyocytes. Toxicon 2013, 75, 3–15. [Google Scholar] [CrossRef]
- Boente-Juncal, A.; Raposo-García, S.; Vale, C.; Louzao, M.C.; Otero, P.; Botana, L.M. In vivo evaluation of the chronic oral toxicity of the marine toxin palytoxin. Toxins 2020, 12, 489. [Google Scholar] [CrossRef]
- Bidard, J.N.; Vijverberg, H.P.M.; Frelin, C. Ciguatoxin is a novel of type of Na+ channel toxin. J. Biol. Chem. 1984, 259, 8353–8357. [Google Scholar] [CrossRef]
- Lombet, A.; Bidard, J.N.; Lazdunski, M. Ciguatoxin and brevetoxins share a common receptor site on the neuronal voltage-dependent Na+ channel. FEBS Lett. 1987, 219, 355–359. [Google Scholar] [CrossRef] [Green Version]
- Pottier, I.; Vernoux, J.P.; Jones, A.; Lewis, R.J. Characterisation of multiple Caribbean ciguatoxins and congeners in individual specimens of horse-eye jack (Caranx latus) by high-performance liquid chromatography/mass spectrometry. Toxicon 2002, 40, 929–939. [Google Scholar] [CrossRef]
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P.; et al. An updated review of ciguatera fish poisoning: Clinical, epidemiological, environmental, and public health management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Otero, P.; Pe, S.; Alfonso, A.; Vale, C.; Rodrı, P.; Gouveia, N.N.; Gouveia, N.; Vale, P.; Hirama, M.; Ishihara, Y.; et al. First Toxin Profile of Ciguateric Fish in Madeira Arquipelago (Europe). Anal. Chem. 2010, 6032–6039. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Benford, D.; Boobis, A.; Ceccatelli, S.; Cravedi, J.; di Domenico, A.; Doerge, D.; Dogliotti, E.; Edler, L.; Farmer, P.; et al. Scientific Opinion on marine biotoxins in shellfish—Emerging toxins: Ciguatoxin group. EFSA J. 2010, 8, 1–38. [Google Scholar] [CrossRef]
- Celis, J.S.; Mancera, J.E. Ciguatera En Las Islas Del Caribe Durante 31. Boletín Investig. Mar. Costeras 2015, 44, 7–32. [Google Scholar]
- Skinner, M.P.; Brewer, T.D.; Johnstone, R.; Fleming, L.E.; Lewis, R.J. Ciguatera fish poisoning in the pacific islands (1998 to 2008). PLoS Negl. Trop. Dis. 2011, 5, e1416. [Google Scholar] [CrossRef]
- Kibler, S.R.; Tester, P.A.; Kunkel, K.E.; Moore, S.K.; Litaker, R.W. Effects of ocean warming on growth and distribution of dinoflagellates associated with ciguatera fish poisoning in the Caribbean. Ecol. Modell. 2015, 316, 194–210. [Google Scholar] [CrossRef] [Green Version]
- Fraga, S.; Rodríguez, F.; Caillaud, A.; Diogène, J.; Raho, N.; Zapata, M. Gambierdiscus excentricus sp. nov. (Dinophyceae), a benthic toxic dinoflagellate from the Canary Islands (NE Atlantic Ocean). Harmful Algae 2011, 11, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, B.; Hurbungs, M.; Jones, A.; Lewis, R.J. Multiple ciguatoxins present in Indian Ocean reef fish. Toxicon 2002, 40, 1347–1353. [Google Scholar] [CrossRef]
- Mak, Y.L.; Wai, T.C.; Murphy, M.B.; Chan, W.H.; Wu, J.J.; Lam, J.C.W.; Chan, L.L.; Lam, P.K.S. Pacific ciguatoxins in food web components of coral reef systems in the Republic of Kiribati. Environ. Sci. Technol. 2013, 47, 14070–14079. [Google Scholar] [CrossRef]
- Food and Drug Administration Handbook Fish and Fishery Products Hazard and Control Guidance. 2021; pp. 1–401. Available online: https://www.fda.gov/media/80637/download (accessed on 21 January 2022).
- Silva, M.; Rodriguez, I.; Barreiro, A.; Kaufmann, M.; Neto, A.I.; Hassouani, M.; Sabour, B.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. First report of ciguatoxins in two starfish species: Ophidiaster ophidianus and Marthasterias glacialis. Toxins 2015, 7, 3740–3757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Arellano, J.L.; Luzardo, O.P.; Brito, A.P.; Cabrera, M.H.; Zumbado, M.; Carranza, C.; Angel-Moreno, A.; Dickey, R.W.; Boada, L.D. Ciguatera fish poisoning, Canary Islands. Emerg. Infect. Dis. 2005, 11, 1981–1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA. Fish and Fishery Products Hazards and Controls Guidance. Fourth Edition. 2011; pp. 1–401. Available online: https://www.federalregister.gov/documents/2011/04/28/2011-10234/guidance-for-industry-on-fish-and-fishery-products-hazards-and-controls-fourth-edition-availability (accessed on 21 January 2022).
- Casteleyn, C.; Van Den Broeck, W.; Simoens, P. Regulation (EC) No 854/2004 laying down specific rules for the organisation of official controls on products of animal origin intended for human consumption is not in compliance with official anatomical nomenclature. Vlaams Diergeneeskd. Tijdschr. 2007, 76, 10–13. [Google Scholar]
- Vale, P. Biotoxinas emergentes em águas europeias e novos riscos para a saúde pública. Rev. Port. Saude Publica 2011, 29, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Abraham, A.; Wang, Y.; El Said, K.R.; Plakas, S.M. Characterization of brevetoxin metabolism in Karenia brevis bloom-exposed clams (Mercenaria sp.) by LC-MS/MS. Toxicon 2012, 60, 1030–1040. [Google Scholar] [CrossRef]
- Anses Opinion, the French Agency for Environmental and Occupational Health Safety. Public Health 2010, 33, 1–12.
- Morris, P.D.; Campbell, D.S.; Taylor, T.J.; Freeman, J.I. Clinical and epidemiological features of neurotoxic shellfish poisoning in North Carolina. Am. J. Public Health 1991, 81, 471–474. [Google Scholar] [CrossRef] [Green Version]
- Pierce, R.H.; Henry, M.S.; Blum, P.C.; Hamel, S.L.; Kirkpatrick, B.; Cheng, Y.S.; Zhou, Y.; Irvin, C.M.; Naar, J.; Weidner, A.; et al. Brevetoxin composition in water and marine aerosol along a Florida beach: Assessing potential human exposure to marine biotoxins. Harmful Algae 2005, 4, 965–972. [Google Scholar] [CrossRef]
- Abraham, A.; Flewelling, L.J.; El Said, K.R.; Odom, W.; Geiger, S.P.; Granholm, A.A.; Jackson, J.T.; Bodager, D. An occurrence of neurotoxic shellfish poisoning by consumption of gastropods contaminated with brevetoxins. Toxicon 2021, 191, 9–17. [Google Scholar] [CrossRef]
- Plakas, S.M.; Dickey, R.W. Advances in monitoring and toxicity assessment of brevetoxins in molluscan shellfish. Toxicon 2010, 56, 137–149. [Google Scholar] [CrossRef]
- Konoki, K.; Baden, D.G.; Scheuer, T.; Catterall, W.A. Molecular determinants of brevetoxin binding to voltage-gated sodium channels. Toxins 2019, 11, 513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amzil, Z.; Derrien, A.; Terrillon, A.T.; Duval, A.; Connes, C.; Marco-Miralles, F.; Nézan, E.; Mertens, K.N. Monitoring the emergence of algal toxins in shellfish: First report on detection of brevetoxins in French mediterranean mussels. Mar. Drugs 2021, 19, 393. [Google Scholar] [CrossRef] [PubMed]
- Band-Schmidt, C.J.; Martínez-López, A.; Bustillos-Guzmán, J.J.; Carreón-Palau, L.; Morquecho, L.; Olguín-Monroy, N.O.; Zenteno-Savín, T.; Mendoza-Flores, A.; González-Acosta, B.; Hernández-Sandoval, F.H.; et al. Morphology, biochemistry, and growth of raphidophyte strains from the Gulf of California. Hydrobiologia 2012, 693, 81–97. [Google Scholar] [CrossRef]
- Arnich, N.; Abadie, E.; Amzil, Z.; Bottein, M.Y.D.; Comte, K.; Chaix, E.; Delcourt, N.; Hort, V.; Mattei, C.; Molgó, J.; et al. Guidance level for brevetoxins in french shellfish. Mar. Drugs 2021, 19, 520. [Google Scholar] [CrossRef]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef] [Green Version]
- Pires, O.R.; Sebben, A.; Schwartz, E.F.; Bloch, C.; Morales, R.A.V.; Schwartz, C.A. The occurrence of 11-oxotetrodotoxin, a rare tetrodotoxin analogue, in the brachycephalidae frog Brachycephalus ephippium. Toxicon 2003, 42, 563–566. [Google Scholar] [CrossRef]
- Miyazawa, K.; Noguchi, T. Distribution and origin of tetrodotoxin. J. Toxicol. Toxin Rev. 2001, 20, 11–33. [Google Scholar] [CrossRef]
- Noguchi, T.; Arakawa, O.; Takatani, T. TTX accumulation in pufferfish. Comp. Biochem. Physiol. Part D Genom. Proteom. 2006, 1, 145–152. [Google Scholar] [CrossRef]
- Wu, Z.; Xie, L.; Xia, G.; Zhang, J.; Nie, Y.; Hu, J.; Wang, S.; Zhang, R. A new tetrodotoxin-producing actinomycete, Nocardiopsis dassonvillei, isolated from the ovaries of puffer fish Fugu rubripes. Toxicon 2005, 45, 851–859. [Google Scholar] [CrossRef]
- Magarlamov, T.Y.; Melnikova, D.I.; Chernyshev, A.V. Tetrodotoxin-producing bacteria: Detection, distribution and migration of the toxin in aquatic systems. Toxins 2017, 9, 166. [Google Scholar] [CrossRef]
- Scheib, H.; McLay, I.; Guex, N.; Clare, J.J.; Blaney, F.E.; Dale, T.J.; Tate, S.N.; Robertson, G.M. Modeling the pore structure of voltage-gated sodium channels in closed, open, and fast-inactivated conformation reveals details of site 1 toxin and local anesthetic binding. J. Mol. Model. 2006, 12, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Cestèle, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef]
- Bentur, Y.; Ashkar, J.; Lurie, Y.; Levy, Y.; Azzam, Z.S.; Litmanovich, M.; Golik, M.; Gurevych, B.; Golani, D.; Eisenman, A. Lessepsian migration and tetrodotoxin poisoning due to Lagocephalus sceleratus in the eastern Mediterranean. Toxicon 2008, 52, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Ebesu, J.S.M. Puffer poisoning: Epidemiology and treatment. J. Toxicol. Toxin Rev. 2001, 20, 1–10. [Google Scholar] [CrossRef]
- Saoudi, M.; Rabeh, F.B.; Jammoussi, K.; Abdelmouleh, A.; Belbahri, L.; El Feki, A. Biochemical and physiological responses in Wistar rat after administration of puffer fish (Lagocephalus lagocephalus) flesh. J. Food Agric. Environ. 2007, 5, 107–111. [Google Scholar]
- Katikou, P.; Gokbulut, C.; Kosker, A.R.; Campàs, M.; Ozogul, F. An Updated Review of Tetrodotoxin and Its Peculiarities. Mar Drugs. 2022, 20, 47. [Google Scholar] [CrossRef] [PubMed]
- Leão, J.M.; Lozano-Leon, A.; Giráldez, J.; Vilariño, Ó.; Gago-Martínez, A. Preliminary results on the evaluation of the occurrence of tetrodotoxin associated to marine vibrio spp. in bivalves from the galician rias (Northwest of Spain). Mar. Drugs 2018, 16, 81. [Google Scholar] [CrossRef] [Green Version]
- Bordin, P.; Dall’Ara, S.; Tartaglione, L.; Antonelli, P.; Calfapietra, A.; Varriale, F.; Guiatti, D.; Milandri, A.; Dell’Aversano, C.; Arcangeli, G.; et al. First occurrence of tetrodotoxins in bivalve molluscs from Northern Adriatic Sea (Italy). Food Control 2021, 120, 107510. [Google Scholar] [CrossRef]
- Vlamis, A.; Katikou, P.; Rodriguez, I.; Rey, V.; Alfonso, A.; Papazachariou, A.; Zacharaki, T.; Botana, A.M.; Botana, L.M. First detection of tetrodotoxin in greek shellfish by UPLC-MS/MS potentially linked to the presence of the dinoflagellate prorocentrum minimum. Toxins 2015, 7, 1779–1807. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.D.; Dhanji-Rapkova, M.; Coates, L.; Bickerstaff, L.; Milligan, S.; O’Neill, A.; Faulkner, D.; McEneny, H.; Baker-Austin, C.; Lees, D.N.; et al. Detection of Tetrodotoxin Shellfish Poisoning (TSP) toxins and causative factors in bivalve molluscs from the UK. Mar. Drugs 2017, 15, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katikou, P.; Georgantelis, D.; Sinouris, N.; Petsi, A.; Fotaras, T. First report on toxicity assessment of the Lessepsian migrant pufferfish Lagocephalus sceleratus (Gmelin, 1789) from European waters (Aegean Sea, Greece). Toxicon 2009, 54, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Boente-Juncal, A.; Vale, C.; Cifuentes, M.; Otero, P.; Camiña, M.; Rodriguez-Vieytes, M.; Botana, L.M. Chronic in vivo effects of repeated exposure to low oral doses of tetrodotoxin: Preliminary evidence of nephrotoxicity and cardiotoxicity. Toxins 2019, 11, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, L.P.; Vilariño, N.; Molgó, J.; Aráoz, R.; Botana, L.M. High-throughput receptor-based assay for the detection of spirolides by chemiluminescence. Toxicon 2013, 75, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, L.P.; Vilariño, N.; Molgó, J.; Aráoz, R.; Louzao, M.C.; Taylor, P.; Talley, T.; Botana, L.M. Development of a Solid-Phase Receptor-Based Assay for the Detection of Cyclic Imines Using a Microsphere-Flow Cytometry System. Anal. Chem. 2013, 85, 2340–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aráoz, R.; Nnghiem, H.-O.; Molgó, J.; Botana, L.M.; Vilariño, N. Method for Manufacturing an Analysis Substrate, and Use Thereof for Detecting Toxins; WIPO: Geneva, Switzerland, 2012. [Google Scholar]
- Noirmain, F.; Dano, J.; Volland, H.; Simon, S.; Servent, D.; Aráoz, R. Neurotorp: Fast & Early Warning Device for the Detection of Aquatic Neurotoxins Principle LFT-TR. In Proceedings of the 18th International Conference on Harmful Algae from Ecosystems to Socio-Ecosystems (Icha 2018), Nantes, France, 21–26 October 2018. [Google Scholar]
- Samdal, I.A.; Løvberg, K.E.; Briggs, L.R.; Kilcoyne, J.; Xu, J.; Forsyth, C.J.; Miles, C.O. Development of an ELISA for the Detection of Azaspiracids. J. Agric. Food Chem. 2015, 63, 7855–7861. [Google Scholar] [CrossRef] [PubMed]
- Leonardo, S.; Rambla-Alegre, M.; Samdal, I.A.; Miles, C.O.; Kilcoyne, J.; Diogène, J.; O’Sullivan, C.K.; Campàs, M. Immunorecognition magnetic supports for the development of an electrochemical immunoassay for azaspiracid detection in mussels. Biosens. Bioelectron. 2017, 92, 200–206. [Google Scholar] [CrossRef]
- Samdal, I.A.; Løvberg, K.E.; Kristoffersen, A.B.; Briggs, L.R.; Kilcoyne, J.; Forsyth, C.J.; Miles, C.O. A Practical ELISA for Azaspiracids in Shellfish via Development of a New Plate-Coating Antigen. J. Agric. Food Chem. 2019, 67, 2369–2376. [Google Scholar] [CrossRef]
- Alfonso, A.; Fernández-Araujo, A.; Alfonso, C.; Caramés, B.; Tobio, A.; Louzao, M.C.; Vieytes, M.R.; Botana, L.M. Palytoxin detection and quantification using the fluorescence polarization technique. Anal. Biochem. 2012, 424, 64–70. [Google Scholar] [CrossRef]
- Fraga, M.; Vilariño, N.; Louzao, M.C.; Fernández, D.A.; Poli, M.; Botana, L.M. Detection of palytoxin-like compounds by a flow cytometry-based immunoassay supported by functional and analytical methods. Anal. Chim. Acta 2016, 903, 1–12. [Google Scholar] [CrossRef]
- Zamolo, V.A.; Valenti, G.; Venturelli, E.; Chaloin, O.; Marcaccio, M.; Boscolo, S.; Castagnola, V.; Sosa, S.; Berti, F.; Fontanive, G.; et al. Highly sensitive electrochemiluminescent nanobiosensor for the detection of palytoxin. ACS Nano 2012, 6, 7989–7997. [Google Scholar] [CrossRef]
- Loeffler, C.R.; Bodi, D.; Tartaglione, L.; Dell’Aversano, C.; Preiss-Weigert, A. Improving in vitro ciguatoxin and brevetoxin detection: Selecting neuroblastoma (Neuro-2a) cells with lower sensitivity to ouabain and veratridine (OV-LS). Harmful Algae 2021, 103, 101994. [Google Scholar] [CrossRef] [PubMed]
- Soliño, L.; Widgy, S.; Pautonnier, A.; Turquet, J.; Loeffler, C.R.; Flores Quintana, H.A.; Diogène, J. Prevalence of ciguatoxins in lionfish (Pterois spp.) from Guadeloupe, Saint Martin, and Saint Barthélmy Islands (Caribbean). Toxicon 2015, 102, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Roué, M.; Smith, K.F.; Sibat, M.; Viallon, J.; Henry, K.; Ung, A.; Biessy, L.; Hess, P.; Darius, H.T.; Chinain, M. Assessment of ciguatera and other phycotoxin-related risks in anaho bay (Nuku Hiva Island, French Polynesia): Molecular, toxicological, and chemical analyses of passive samplers. Toxins 2020, 12, 321. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Asencio, L.; Clausing, R.J.; Rañada, M.L.; Alonso-Hernández, C.M.; Dechraoui Bottein, M.Y. A radioligand receptor binding assay for ciguatoxin monitoring in environmental samples: Method development and determination of quality control criteria. J. Environ. Radioact. 2018, 192, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Flewelling, L.J.; Corcoran, A.A.; Granholm, A.A.; Takeuchi, N.Y.; Van Hoeck, R.V.; Zahara, M.L. Validation and Assessment of an Enzyme-Linked Immunosorbent Assay (Elisa) for Use in Monitoring and Managing Neurotoxic Shellfish Poisoning. J. Shellfish Res. 2020, 39, 491–500. [Google Scholar] [CrossRef]
- McCall, J.R.; Jacocks, H.M.; Niven, S.C.; Poli, M.A.; Baden, D.G.; Bourdelais, A.J. Development and utilization of a fluorescence-based receptor-binding assay for the site 5 voltage-sensitive sodium channel ligands brevetoxin and ciguatoxin. J. AOAC Int. 2014, 97, 307–315. [Google Scholar] [CrossRef]
- Reverté, L.; De La Iglesia, P.; Del Río, V.; Campbell, K.; Elliott, C.T.; Kawatsu, K.; Katikou, P.; Diogène, J.; Campàs, M. Detection of Tetrodotoxins in Puffer Fish by a Self-Assembled Monolayer-Based Immunoassay and Comparison with Surface Plasmon Resonance, LC-MS/MS, and Mouse Bioassay. Anal. Chem. 2015, 87, 10839–10847. [Google Scholar] [CrossRef]
- Reverté, L.; Rambla-Alegre, M.; Leonardo, S.; Bellés, C.; Campbell, K.; Elliott, C.T.; Gerssen, A.; Klijnstra, M.D.; Diogène, J.; Campàs, M. Development and validation of a maleimide-based enzyme-linked immunosorbent assay for the detection of tetrodotoxin in oysters and mussels. Talanta 2018, 176, 659–666. [Google Scholar] [CrossRef] [Green Version]
- Reverté, L.; Campàs, M.; Yakes, B.J.; Deeds, J.R.; Katikou, P.; Kawatsu, K.; Lochhead, M.; Elliott, C.T.; Campbell, K. Tetrodotoxin detection in puffer fish by a sensitive planar waveguide immunosensor. Sensors Actuators B Chem. 2017, 253, 967–976. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.A.; Kværnø, L.; Dunn, T.B.; Beauchemin, A.; Raymer, B.; Mulder, J.A.; Olhava, E.J.; Juhl, M.; Kagechika, K.; Favor, D.A. Total Synthesis of (+)-Azaspiracid-1. An Exhibition of the Intricacies of Complex Molecule Synthesis. J. Am. Chem. Soc. 2008, 130, 16295–16309. [Google Scholar] [CrossRef] [Green Version]
- Okumu, A.A.; Forsyth, C.J. Synthesis of the C1–C19 Domain of Azaspiracid-34. Org. Lett. 2019, 21, 356–359. [Google Scholar] [CrossRef] [PubMed]
- Kilcoyne, J.; McCoy, A.; Burrell, S.; Krock, B.; Tillmann, U. Effects of Temperature, Growth Media, and Photoperiod on Growth and Toxin Production of Azadinium spinosum. Mar Drugs 2019, 17, 489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfonso, C.; Alfonso, A.; Otero, P.; Rodríguez, P.; Vieytes, M.R.; Elliot, C.; Higgins, C.; Botana, L.M. Purification of five azaspiracids from mussel samples contaminated with DSP toxins and azaspiracids. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2008, 865, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, C.J.; Xu, J.; Nguyen, S.T.; Samdal, I.A.; Briggs, L.R.; Rundberget, T.; Sandvik, M.; Miles, C.O. Antibodies with broad specificity to azaspiracids by use of synthetic haptens. J. Am. Chem. Soc. 2006, 128, 15114–15116. [Google Scholar] [CrossRef] [PubMed]
- Bellocci, M.; Sala, G.L.; Callegari, F.; Rossini, G.P. Azaspiracid-1 Inhibits Endocytosis of Plasma Membrane Proteins in Epithelial Cells. Toxicol. Sci. 2010, 117, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Leonardo, S.; Kilcoyne, J.; Samdal, I.A.; Miles, C.O.; O’Sullivan, C.K.; Diogène, J.; Campàs, M. Detection of azaspiracids in mussels using electrochemical immunosensors for fast screening in monitoring programs. Sens. Actuators B Chem. 2018, 262, 818–827. [Google Scholar] [CrossRef] [Green Version]
- Frederick, M.O.; De Lamo, S.; Janda, K.D.; Nicolaou, K.C.; Tobin, J.D. Monoclonal Antibodies with Orthogonal Azaspiracid Epitopes. ChemBioChem 2009, 10, 1625–1629. [Google Scholar] [CrossRef]
- Dillon, M.; Zaczek-Moczydlowska, M.A.; Edwards, C.; Turner, A.D.; Miller, P.I.; Moore, H.; McKinney, A.; Lawton, L.; Campbell, K. Current trends and challenges for rapid smart diagnostics at point-of-site testing for marine toxins. Sensors 2021, 21, 2499. [Google Scholar] [CrossRef]
- Brissard, C.; Herrenknecht, C.; Séchet, V.; Hervé, F.; Pisapia, F.; Harcouet, J.; Lémée, R.; Chomérat, N.; Hess, P.; Amzil, Z. Complex toxin profile of French Mediterranean Ostreopsis cf. ovata strains, seafood accumulation and ovatoxins prepurification. Mar. Drugs 2014, 12, 2851–2876. [Google Scholar] [CrossRef] [Green Version]
- Chinain, M.; Gatti, C.M.I.; Darius, H.T.; Quod, J.P.; Tester, P.A. Ciguatera poisonings: A global review of occurrences and trends. Harmful Algae 2021, 102, 101873. [Google Scholar] [CrossRef]
- Soliño, L.; Costa, P.R. Differential toxin profiles of ciguatoxins in marine organisms: Chemistry, fate and global distribution. Toxicon 2018, 150, 124–143. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.R.; Estévez, P.; Soliño, L.; Castro, D.; Rodrigues, S.M.; Timoteo, V.; Leao-Martins, J.M.; Santos, C.; Gouveia, N.; Diogène, J.; et al. An update on ciguatoxins and ctx-like toxicity in fish from different trophic levels of the selvagens islands (Ne atlantic, madeira, portugal). Toxins 2021, 13, 580. [Google Scholar] [CrossRef] [PubMed]
- Sibat, M.; Herrenknecht, C.; Darius, H.T.; Roué, M.; Chinain, M.; Hess, P. Detection of pacific ciguatoxins using liquid chromatography coupled to either low or high resolution mass spectrometry (LC-MS/MS). J. Chromatogr. A 2018, 1571, 16–28. [Google Scholar] [CrossRef]
- Gago-Martinez, A.; Leão, J.M.; Estevez, P.; Castro, D.; Barrios, C.; Hess, P.; Sibat, M. Characterisation of ciguatoxins. EFSA Support. Publ. 2021, 18, 6649E. [Google Scholar] [CrossRef]
- McNabb, P.S.; Selwood, A.I.; Van Ginkel, R.; Boundy, M.; Holland, P.T. Determination of brevetoxins in shellfish by LC/MS/MS: Single-laboratory validation. J. AOAC Int. 2012, 95, 1097–1105. [Google Scholar] [CrossRef]
- Rodríguez, I.; Alfonso, A.; González-Jartín, J.M.; Vieytes, M.R.; Botana, L.M. A single run UPLC-MS/MS method for detection of all EU-regulated marine toxins. Talanta 2018, 189, 622–628. [Google Scholar] [CrossRef]
- EURLMB. Determination of Tetrodotoxin by HILIC-MS/MS; European Union Reference Laboratory for Marine Biotoxins: Vigo, Spain, 2017; pp. 1–24. [Google Scholar]
- Mazzeo, A.; Varra, M.; Tartaglione, L.; Ciminiello, P.; Zendong, Z.; Hess, P.; Dell’aversano, C. Toward isolation of palytoxins: Liquid chromatography coupled to low-or high-resolution mass spectrometry for the study on the impact of drying techniques, solvents and materials. Toxins 2021, 13, 650. [Google Scholar] [CrossRef]


| Method | Procedure | Toxins/Matrix Tested | Range or LOQ | Refs. |
|---|---|---|---|---|
| Cyclic Imines | ||||
| Receptor-based method (FP) | A direct assay based on binding SPXs to nAChRs from T. marmorata membranes. | SPX-13/shellfish | 50–350 µg SPX-13/kg meat | [50] |
| Receptor-based method (FP) | Competition between SPXs and a-bungarotoxin for binding to nAChRs. | SPX-13/shellfish | 40–200 µg SPX-13/kg meat | [42] |
| Receptor-based method (Chemiluminescence) | Competition between SPXs and biotin-α-bungarotoxin immobilized on a streptavidin-coated surface, for binding to nAChRs. | SPX-13/shellfish | 50 μg SPX-13/kg meat. | [153] |
| Solid-Phase Receptor-Based Assay (microsphere-flow cytometry system). | Immobilization of nAChR or Ls-AChBP on the surface of carboxylated microspheres and the competition of CIs with biotin-α-BTX for binding to these proteins. | SPX-13/shellfish | 10–6000 μg SPX-13/kg of meat and a LOD of 3 μg SPX/kg. | [154] |
| Non-radioactive Microplate-Receptor Binding Assay (ABRAXIS) | Neurotoxins competitively inhibit biotinylated-α-BTX binding to nAChR in a concentration-dependent manner. | CIs, ATXs | nM range | [155] |
| Toxin-receptor lateral flow test “NeuroTorp” | Based on the immobilization of nAChR on high porosity borosilicate membrane filter support, and the use of a biotinylated α-BTX as toxin-tracer. | ATX-a and CIs (PnTXs, SPXs, GYM) | nM range | [156] |
| Emerging Azaspiracids | ||||
| Immunoassay | ELISA. Ovine polyclonal antibodies | AZA1-3 and Emerging AZAs including AZA-4−10, -33, and -34 and 37-epi-AZA-1. AZA-17 and AZA-19. | 57 μg/kg shellfish | [157] |
| Immunoassay | ELISA. Antibody immobilization supports MBs. Tracer: AZA-HRP | 63 μg AZA-1 eq./kg) | [158] | |
| Immunoassay | ELISA. Plate-coater: OVA−cdiAZA1. | AZA reference materials as well as the precursors to AZA-3 and AZA-6, | 37 μg/kg for AZA-1 in shellfish. | [159] |
| Palytoxins | ||||
| Receptor based method (FP) | Based on the interaction between the Na, K-ATPase, and PLTX. | PLTX/mussels, and ostreopsis | LOQ = 10 Nm LOD = 2 Nm | [160] |
| Immunodetection method (microspheres coupled to flow-cytometry detection). | Based on the competition between free PLTXs in solution and PLTX immobilized on the surface of microspheres for binding to a specific monoclonal anti-PLTX antibody. | PLTXs/musels | Dynamic range: 0.47–6.54 Nm and LOQ: 374–4430 μg/kg. | [161] |
| Electrochemiluminescence method | Electrochemiluminescence is directly proportional to PTX | PLTXs/mussel, algal samples | LOD = 220 ng/mL | [162] |
| Ciguatoxins | ||||
| Cell-based assay | Sensitivity to neuroblastoma N2a cell line | CTX-3C and CTX-1B/fish flesh | 1.35 pg CTX-3C/mL and 2.06 pg CTX-1B/mL | [163] |
| Cell-based assay | Sensitivity to neuroblastoma N2a cell line | P-CTX-1 eqs/lionfish | 0.0039 ppb–0.0096 ppb P-CTX-1 eq. | [164] |
| Cell-based assay | Sensitivity to neuroblastoma N2a cell line | P-CTX-1/SPATT | 0.02 ng P-CTX3C eq./g | [165] |
| Immunoassay | Radioligand receptor binding assay | P-CTX-3C/fish flesh | 0.75 ng P-CTX-3C eq./g | [166] |
| Brevetoxins | ||||
| Cell-based assay | Sensitivity to neuroblastoma N2a cell line | BTX-3/fish flesh | 3.04 ng BTX-3/mL | [163] |
| Immunoassay | ELISA | BTX-3/clam and oyster | 0.04 µg BTX-3 eq./g shellfish | [167] |
| Immunoassay | Radioligand receptor binding assay | BTX-1, BTX-3, BTX-9/K. brevis | 1 Pm to 1 µM BTX-2 | [168] |
| Tetrodotoxins | ||||
| Cell-based assay | Sensitivity to neuroblastoma N2a cell line | TTX/shellfish | 20 µg TTX/kg | [169] |
| Immunoassay | Competitive inhibition enzymatic immunoassay (Melisa) | TTX/mussels and oysters | 20 µg TTX/kg and 30 µg TTX/kg | [170] |
| SPR | Nanoarray planar waveguide biosensor | TTX/puffer fish | 0.4 to 3.29 mg/kg | [171] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otero, P.; Silva, M. Emerging Marine Biotoxins in European Waters: Potential Risks and Analytical Challenges. Mar. Drugs 2022, 20, 199. https://doi.org/10.3390/md20030199
Otero P, Silva M. Emerging Marine Biotoxins in European Waters: Potential Risks and Analytical Challenges. Marine Drugs. 2022; 20(3):199. https://doi.org/10.3390/md20030199
Chicago/Turabian StyleOtero, Paz, and Marisa Silva. 2022. "Emerging Marine Biotoxins in European Waters: Potential Risks and Analytical Challenges" Marine Drugs 20, no. 3: 199. https://doi.org/10.3390/md20030199
APA StyleOtero, P., & Silva, M. (2022). Emerging Marine Biotoxins in European Waters: Potential Risks and Analytical Challenges. Marine Drugs, 20(3), 199. https://doi.org/10.3390/md20030199