
Marine Migrastatics: A Comprehensive 2022 Update



Abstract
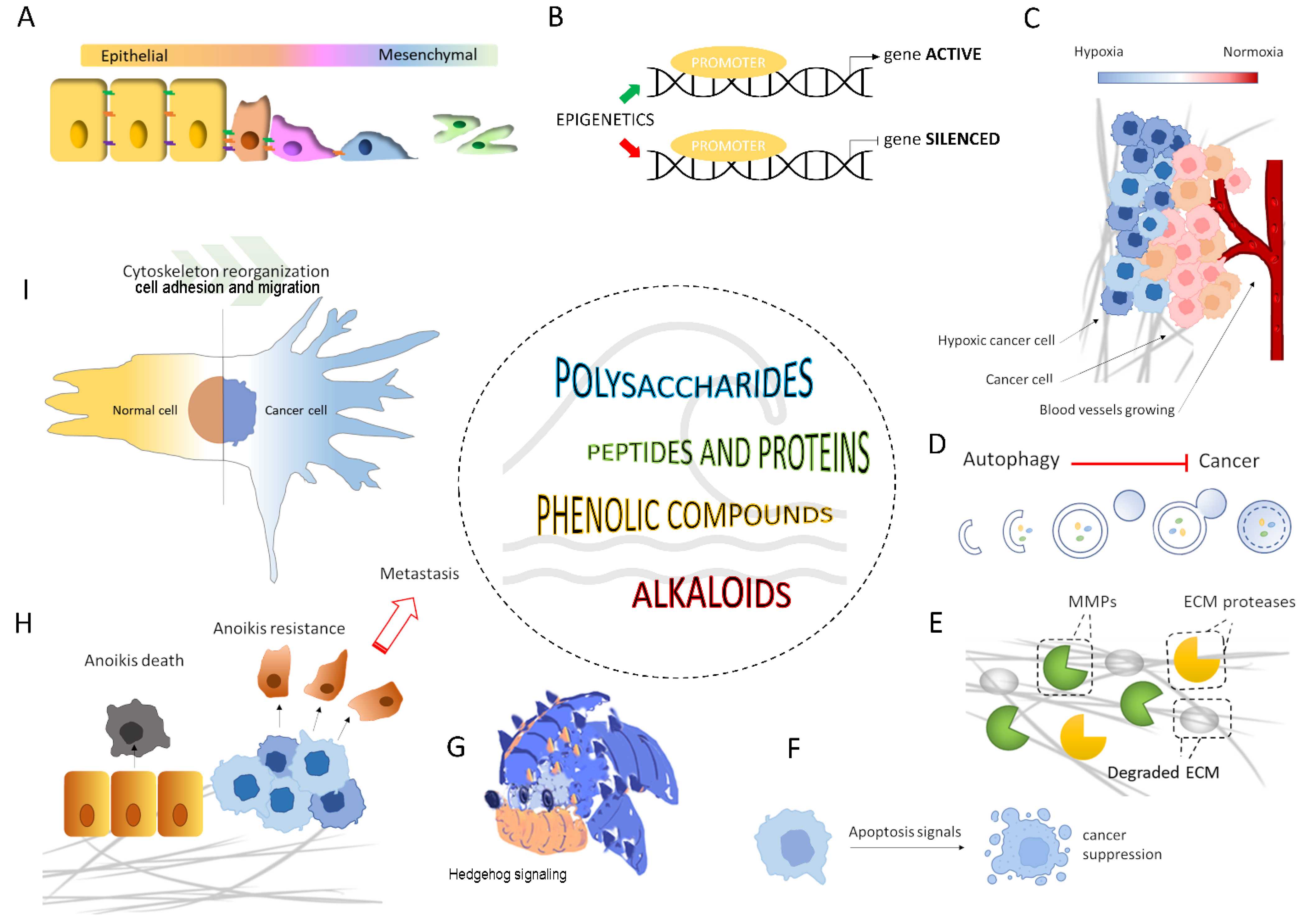
:1. Introduction
2. Biomechanisms Underlying Cancer Metastasis as Therapeutic Molecular Targets
2.1. Epithelial-Mesenchymal Transition (EMT) and EMT-Related Signaling Pathways
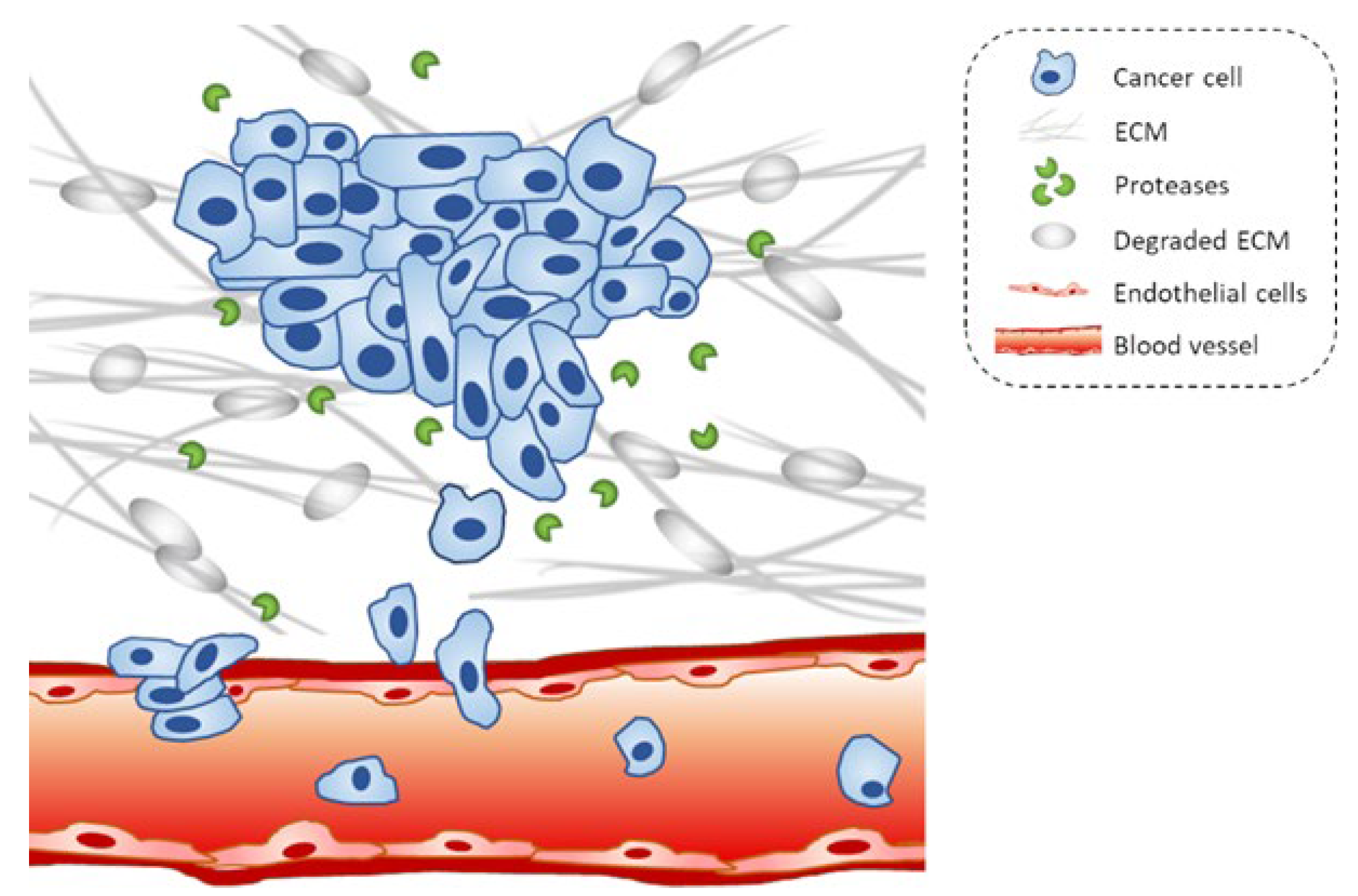
2.2. ECM Disassembly and the Role of Matrix Metalloproteinases (MMPs)
2.3. Anoikis Resistance
2.4. Cytoskeletal Remodeling and Cell Adhesion
2.5. Hypoxia and Neo-Angiogenesis
2.6. Apoptosis and Autophagy
2.7. Epigenetic Control
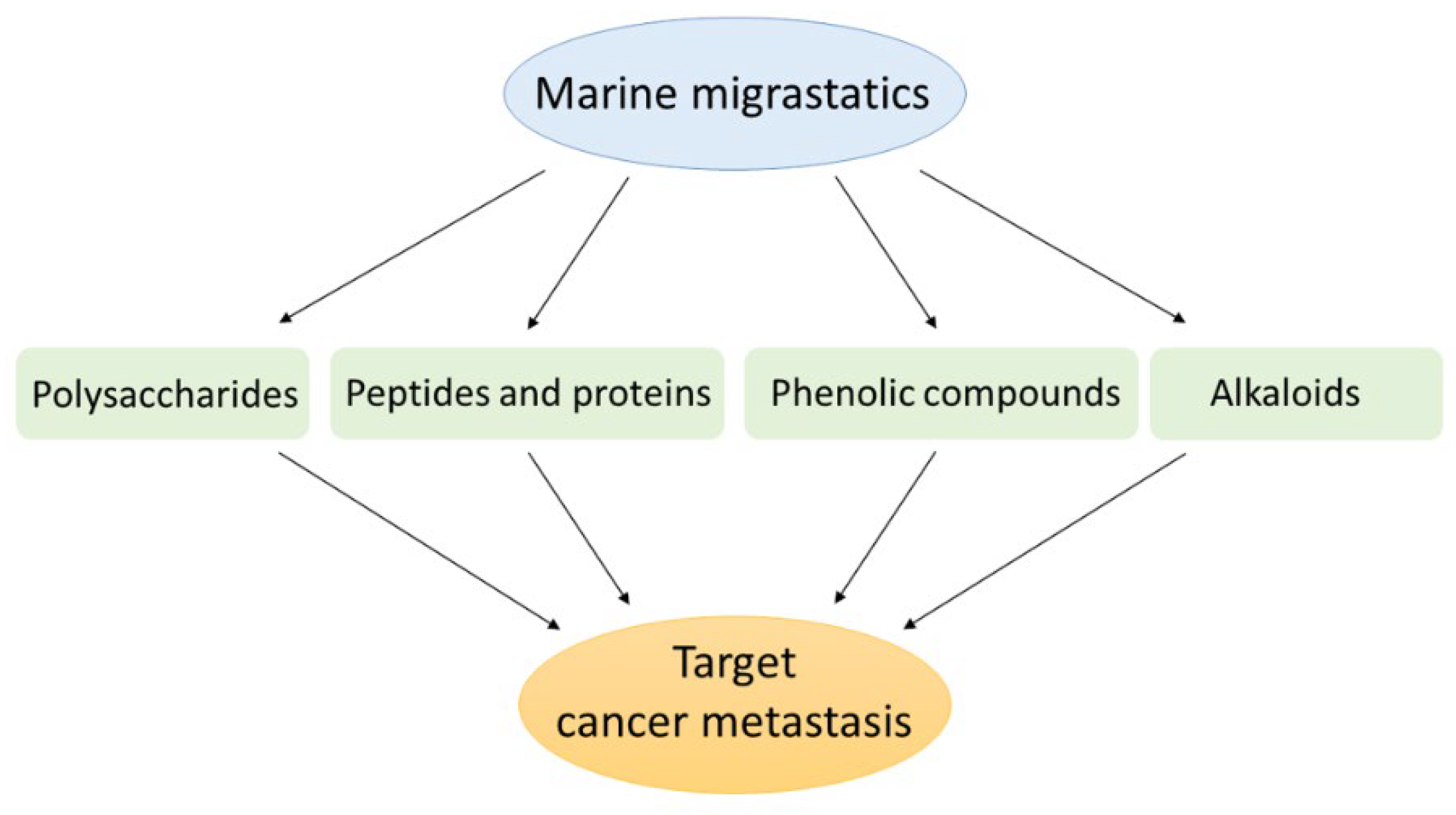
3. Marine Compounds as Migrastatics
3.1. Marine Polysaccharides
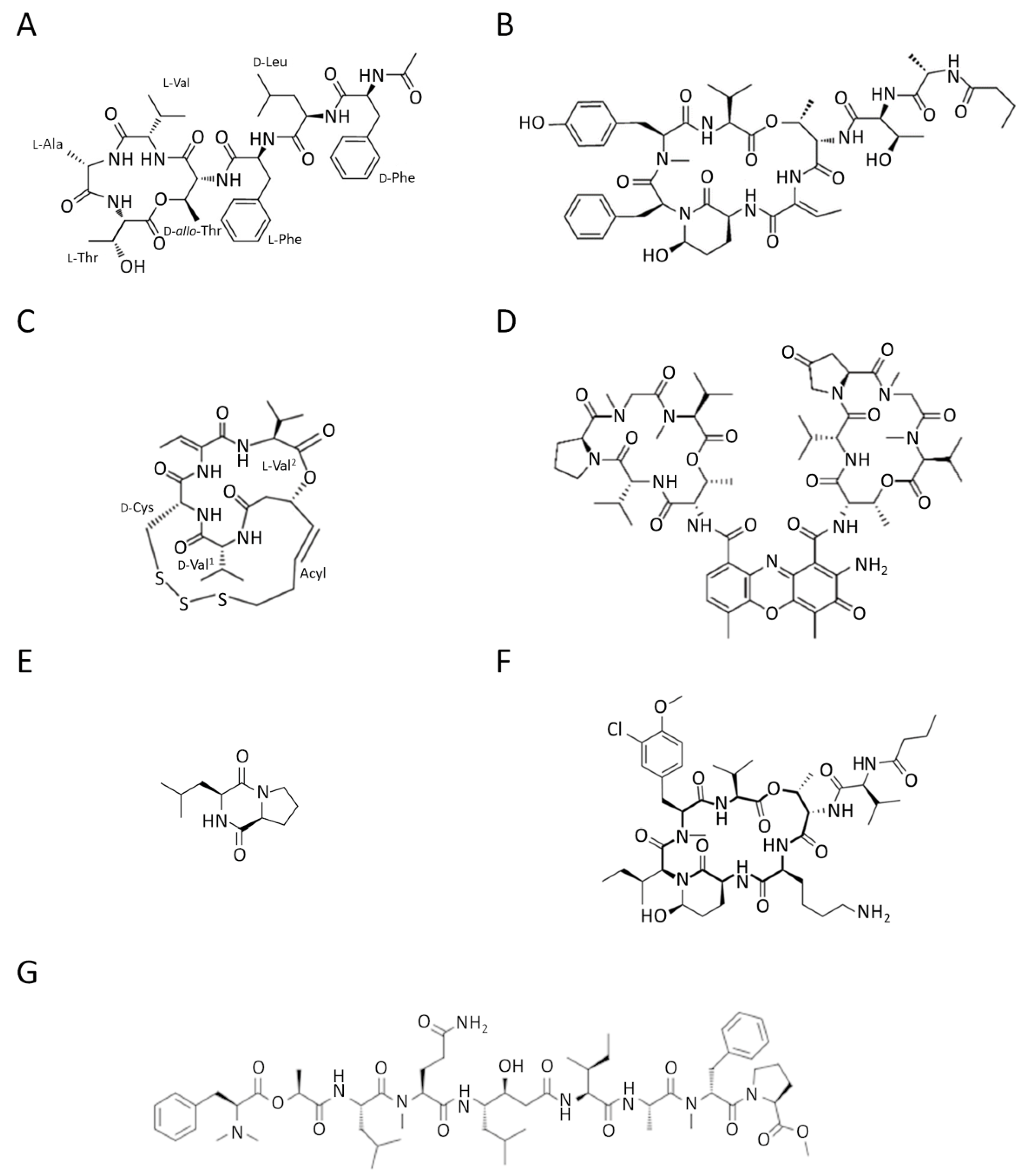
3.2. Marine Peptides and Proteins
3.3. Marine Phenolic Compounds
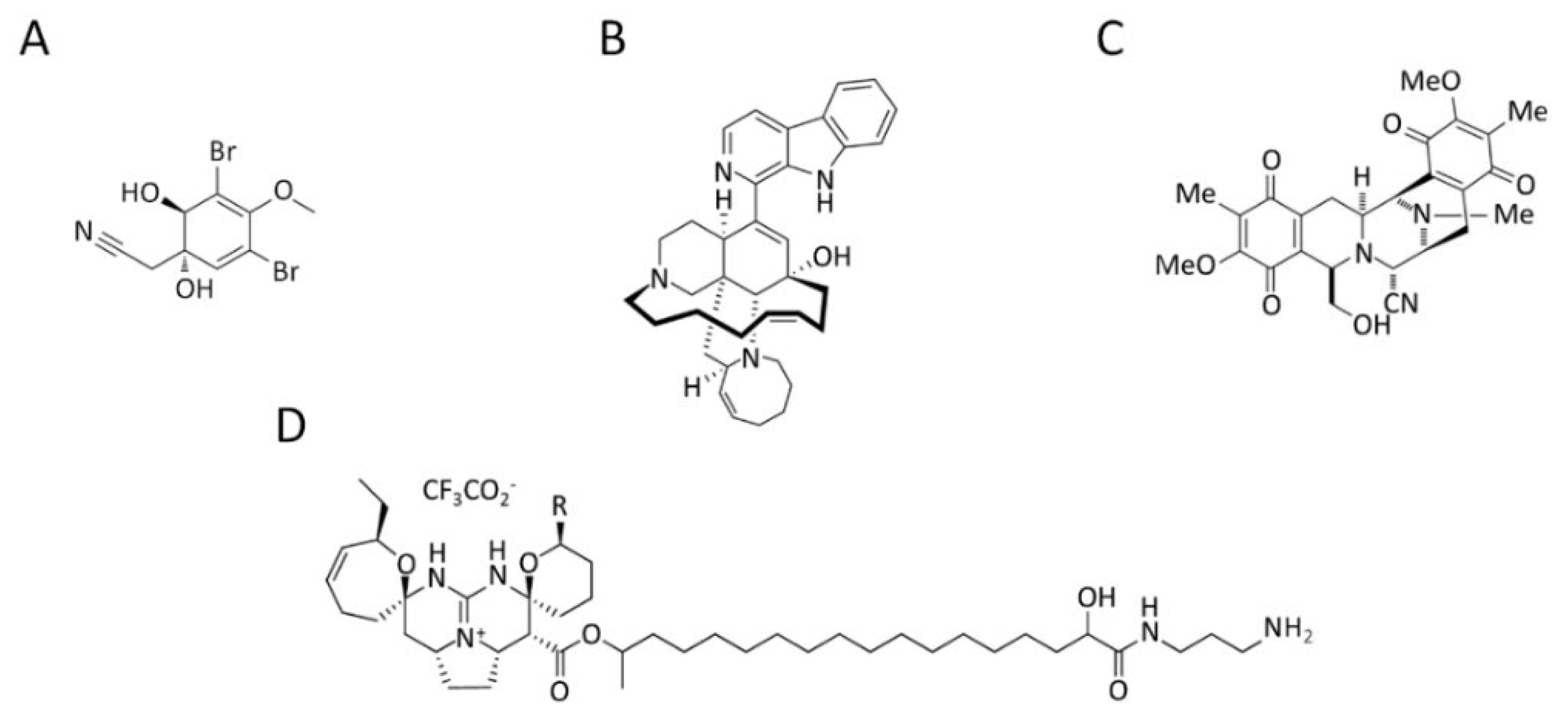
3.4. Marine Alkaloids
4. Research Methodology
5. Summary and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Lambert, A.W.; Pattabiraman, D.R.; Weinberg, R.A. Emerging Biological Principles of Metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sayes, N.; Vito, A.; Mossman, K. Tumor Heterogeneity: A Great Barrier in the Age of Cancer Immunotherapy. Cancers 2021, 13, 806. [Google Scholar] [CrossRef] [PubMed]
- Gandalovičová, A.; Rosel, D.; Fernandes, M.; Veselý, P.; Heneberg, P.; Čermák, V.; Petruželka, L.; Kumar, S.; Sanz-Moreno, V.; Brábek, J. Migrastatics-Anti-metastatic and Anti-invasion Drugs: Promises and Challenges. Trends Cancer 2017, 3, 391–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayat Mokhtari, R.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination therapy in combating cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, J.; Raškova, M.; Rösel, D.; Brábek, J.; Gil-Henn, H. Are We Ready for Migrastatics? Cells 2021, 10, 1845. [Google Scholar] [CrossRef] [PubMed]
- Lindequist, U. Marine-Derived Pharmaceuticals–Challenges and Opportunities. Biomol. Ther. 2016, 24, 561–571. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.Y.; Li, H.J.; Li, Q.Y.; Wu, Y.C. Application of marine natural products in drug research. Bioorg. Med. Chem. 2021, 35, 116058. [Google Scholar] [CrossRef]
- Dayanidhi, D.L.; Thomas, B.C.; Osterberg, J.S.; Vuong, M.; Vargas, G.; Kwartler, S.K.; Schmaltz, E.; Dunphy-Daly, M.M.; Schultz, T.F.; Rittschof, D.; et al. Exploring the Diversity of the Marine Environment for New Anti-cancer Compounds. Front. Mar. Sci. 2021, 7, 614766. [Google Scholar] [CrossRef]
- Dyshlovoy, S.A.; Honecker, F. Marine Compounds and Cancer: Updates 2020. Mar. Drugs 2020, 18, 643. [Google Scholar] [CrossRef] [PubMed]
- Marine Pharmacology. Available online: https://www.marinepharmacology.org/approved (accessed on 3 March 2022).
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Sig. Transduct. Target Ther. 2020, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Tamma, R.; Annese, T. Epithelial-Mesenchymal Transition in Cancer: A Historical Overview. Transl. Oncol. 2020, 13, 100773. [Google Scholar] [CrossRef]
- Garg, M. Epithelial-mesenchymal transition-activating transcription factors-multifunctional regulators in cancer. World J. Stem Cells 2013, 5, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Ma, L.; Zhang, Z.; Liu, X.; Gao, H.; Zhuang, Y.; Yang, P.; Kornmann, M.; Tian, X.; Yang, Y. Hedgehog Signaling Regulates Epithelial-Mesenchymal Transition in Pancreatic Cancer Stem-Like Cells. J. Cancer 2016, 7, 408–417. [Google Scholar] [CrossRef] [Green Version]
- Islam, S.S.; Mokhtari, R.B.; Noman, A.S.; Uddin, M.; Rahman, M.Z.; Azadi, M.A.; Zlotta, A.; van der Kwast, T.; Yeger, H.; Farhat, W.A. Sonic hedgehog (Shh) signaling promotes tumorigenicity and stemness via activation of epithelial-to-mesenchymal transition (EMT) in bladder cancer. Mol. Carcinog. 2016, 55, 537–551. [Google Scholar] [CrossRef]
- Riaz, S.K.; Ke, Y.; Wang, F.; Kayani, M.A.; Malik, M.F.A. Influence of SHH/GLI1 axis on EMT mediated migration and invasion of breast cancer cells. Sci. Rep. 2019, 9, 6620. [Google Scholar] [CrossRef]
- Yue, D.; Li, H.; Che, J.; Zhang, Y.; Tseng, H.H.; Jin, J.Q.; Luh, T.M.; Giroux-Leprieur, E.; Mo, M.; Zheng, Q.; et al. Hedgehog/Gli promotes epithelial-mesenchymal transition in lung squamous cell carcinomas. J. Exp. Clin. Cancer Res. 2014, 33, 34. [Google Scholar] [CrossRef] [Green Version]
- Skoda, A.M.; Simovic, D.; Karin, V.; Kardum, V.; Vranic, S.; Serman, L. The role of the Hedgehog signaling pathway in cancer: A comprehensive review. Bosn. J. Basic Med. Sci. 2018, 18, 8–20. [Google Scholar] [CrossRef]
- Kleiner, D.E.; Stetler-Stevenson, W.G. Matrix metalloproteinases and metastasis. Cancer Chemother. Pharmacol. 1999, 43, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Bischoff, R. Physiology and pathophysiology of matrix metalloproteases. Amino Acids 2011, 41, 271–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egeblad, M.; Werb, Z. New functions for the matrix metalloproteinases in cancer progression. Nat. Rev. Cancer 2002, 2, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, A. Anoikis. Cell Death Differ. 2005, 12, 1473–1477. [Google Scholar] [CrossRef]
- Adeshakin, F.O.; Adeshakin, A.O.; Afolabi, L.O.; Yan, D.; Zhang, G.; Wan, X. Mechanisms for Modulating Anoikis Resistance in Cancer and the Relevance of Metabolic Reprogramming. Front. Oncol. 2021, 11, 626577. [Google Scholar] [CrossRef]
- Jacquemet, G.; Hamidi, H.; Ivaska, J. Filopodia in cell adhesion, 3D migration and cancer cell invasion. Curr. Opin. Cell Biol. 2015, 37, 119. [Google Scholar] [CrossRef] [Green Version]
- Arjonen, A.; Kaukonen, R.; Ivaska, J. Filopodia and adhesion in cancer cell motility. Cell Adh. Migr. 2011, 5, 421–430. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.M.; Rodriguez, Y.A.R.; Jeong, K.; Ahn, E.E.; Lim, S.S. Targeting focal adhesion kinase in cancer cells and the tumor microenvironment. Exp. Mol. Med. 2020, 52, 877–886. [Google Scholar] [CrossRef]
- Čermák, V.; Dostál, V.; Jelínek, M.; Libusová, L.; Kovář, J.; Rösel, D.; Brábek, J. Microtubule-targeting agents and their impact on cancer treatment. Eur. J. Cell. Biol. 2020, 99, 151075. [Google Scholar] [CrossRef]
- Rankin, E.B.; Giaccia, A.J. Hypoxic control of metastasis. Science 2016, 352, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Rankin, E.B.; Nam, J.M.; Giaccia, A.J. Hypoxia: Signaling the Metastatic Cascade. Trends Cancer 2016, 2, 295–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as anticancer mechanism: Function and dysfunction of its modulators and targeted therapeutic strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Z.; Yang, Z.; Xu, Y.; Chen, Y.; Yu, Q. Apoptosis, autophagy, necroptosis, and cancer metastasis. Mol. Cancer 2015, 14, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neophytou, C.M.; Trougakos, I.P.; Erin, N.; Papageorgis, P. Apoptosis Deregulation and the Development of Cancer Multi-Drug Resistance. Cancers 2021, 13, 4363. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.W.; Lee, S.H. The Roles of Autophagy in Cancer. Int. J. Mol. Sci. 2018, 19, 3466. [Google Scholar] [CrossRef] [Green Version]
- Fardi, M.; Solali, S.; Farshdousti Hagh, M. Epigenetic mechanisms as a new approach in cancer treatment: An updated review. Genes Dis. 2018, 5, 304–311. [Google Scholar] [CrossRef]
- Ahuja, N.; Sharma, A.R.; Baylin, S.B. Epigenetic Therapeutics: A New Weapon in the War Against Cancer. Annu. Rev. Med. 2016, 67, 73–89. [Google Scholar] [CrossRef] [Green Version]
- Karthikeyan, A.; Joseph, A.; Nair, B.G. Promising bioactive compounds from the marine environment and their potential effects on various diseases. J. Genet. Eng. Biotechnol. 2022, 20, 14. [Google Scholar] [CrossRef]
- Cao, R.; Jin, W.; Shan, Y.; Wang, J.; Liu, G.; Kuang, S.; Sun, C. Marine Bacterial Polysaccharide EPS11 Inhibits Cancer Cell Growth via Blocking Cell Adhesion and Stimulating Anoikis. Mar. Drugs 2018, 16, 85. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, G.; Ma, W.; Lu, Z.; Sun, C. Marine Bacterial Polysaccharide EPS11 Inhibits Cancer Cell Growth and Metastasis via Blocking Cell Adhesion and Attenuating Filiform Structure Formation. Mar. Drugs 2019, 17, 50. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Liu, R.; Shan, Y.; Sun, C. Marine bacterial exopolysaccharide EPS11 inhibits migration and invasion of liver cancer cells by directly targeting collagen I. J. Biol. Chem. 2021, 297, 101133. [Google Scholar] [CrossRef]
- Ou, M.; Sun, X.; Liang, J.; Liu, F.; Wang, L.; Wu, X.; Tu, J. A polysaccharide from Sargassum thunbergii inhibits angiogenesis via downregulating MMP-2 activity and VEGF/HIF-1α signaling. Int. J. Biol. Macromol. 2017, 94, 451–458. [Google Scholar] [CrossRef]
- Malyarenko, O.S.; Usoltseva, R.V.; Shevchenko, N.M.; Isakov, V.V.; Zvyagintseva, T.N.; Ermakova, S.P. In vitro anticancer activity of the laminarans from Far Eastern brown seaweeds and their sulfated derivatives. J. Appl. Phycol. 2017, 29, 543–553. [Google Scholar] [CrossRef]
- Malyarenko, O.S.; Usoltseva, R.V.; Zvyagintseva, T.N.; Ermakova, S.P. Laminaran from brown alga Dictyota dichotoma and its sulfated derivative as radioprotectors and radiosensitizers in melanoma therapy. Carbohydr. Polym. 2019, 206, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, H.; Wang, X.; Zhang, X.; Liu, W.; Wang, Y.; Zhang, Y.; Pan, H.; Wang, Q.; Han, Y. Effect of polysaccharide from Undaria pinnatifida on proliferation, migration and apoptosis of breast cancer cell MCF7. Int. J. Biol. Macromol. 2019, 121, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.; Qin, Z.; Yongping, X.; Gang, C.; Yukuan, Q. Sulfated Polysaccharide From Undaria pinnatifida Induces Apoptosis and Inhibits Proliferation, Migration, and Invasion in Ovarian Cancer via Suppressing the Hedgehog Signaling Pathway. Front. Mater. 2021, 8, 795061. [Google Scholar] [CrossRef]
- Wu, J.; Chen, X.; Qiao, K.; Su, Y.; Liu, Z. Purification, structural elucidation, and in vitro antitumor effects of novel polysaccharides from Bangia fuscopurpurea. Food Sci. Hum. Wellness. 2021, 10, 63–71. [Google Scholar] [CrossRef]
- Wu, J.; Lin, C.; Chen, X.; Pan, N.; Liu, Z. Polysaccharides isolated from Bangia fuscopurpurea induce apoptosis and autophagy in human ovarian cancer A2780 cells. Food Sci. Nutr. 2021, 9, 6707–6719. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, L.; Liu, Y.; Chen, M.; Zhang, S.; Kong, D. Sea cucumber Cucumaria frondosa fucoidan inhibits osteosarcoma adhesion and migration by regulating cytoskeleton remodeling. Oncol. Rep. 2020, 44, 469–476. [Google Scholar] [CrossRef]
- Jiang, W.; Tian, W.; Ijaz, M.; Wang, F. Inhibition of EGF-induced migration and invasion by sulfated polysaccharide of Sepiella maindroni ink via the suppression of EGFR/Akt/p38 MAPK/MMP-2 signaling pathway in KB cells. Biomed. Pharmacother. 2017, 95, 95–102. [Google Scholar] [CrossRef]
- Jiang, W.; Cheng, Y.; Zhao, N.; Li, L.; Shi, Y.; Zong, A.; Wang, F. Sulfated polysaccharide of Sepiella Maindroni ink inhibits the migration, invasion and matrix metalloproteinase-2 expression through suppressing EGFR-mediated p38/MAPK and PI3K/Akt/mTOR signaling pathways in SKOV-3 cells. Int. J. Biol. Macromol. 2018, 107, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Wongprasert, K.; Rudtanatip, T.; Praiboon, J. Immunostimulatory activity of sulfated galactans isolated from the red seaweed Gracilaria fisheri and development of resistance against white spot syndrome virus (WSSV) in shrimp. Fish Shellfish. Immunol. 2014, 36, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Boonsri, B.; Choowongkomon, K.; Kuaprasert, B.; Thitiphatphuvanon, T.; Supradit, K.; Sayinta, A.; Duangdara, J.; Rudtanatip, T.; Wongprasert, K. Probing the Anti-Cancer Potency of Sulfated Galactans on Cholangiocarcinoma Cells Using Synchrotron FTIR Microspectroscopy, Molecular Docking, and In Vitro Studies. Mar. Drugs 2021, 19, 258. [Google Scholar] [CrossRef] [PubMed]
- Eke, I.; Cordes, N. Focal adhesion signaling and therapy resistance in cancer. Semin. Cancer Biol. 2015, 31, 65–75. [Google Scholar] [CrossRef]
- da Silva Barbosa, J.; Palhares, L.C.G.F.; Silva, C.H.F.; Sabry, D.A.; Chavante, S.F.; Rocha, H.A.O. In Vitro Antitumor Potential of Sulfated Polysaccharides from Seaweed Caulerpa cupressoides var. flabellata. Mar. Biotechnol. 2021, 23, 77–89. [Google Scholar] [CrossRef]
- Jose, G.M.; Kurup, G.M. Sulfated polysaccharides from Padina tetrastromatica arrest cell cycle, prevent metastasis and downregulate angiogenic mediators in HeLa cells. Bioact. Carbohydr. Diet. Fibre. 2017, 12, 7–13. [Google Scholar] [CrossRef]
- Shakeel, E.; Arora, D.; Jamal, Q.M.S.; Akhtar, S.; Khan, M.K.A.; Kamal, M.A.; Siddiqui, M.H.; Lohani, M.; Arif, J.M. Marine Drugs: A Hidden Wealth and a New Epoch for Cancer Management. Curr. Drug Metab. 2018, 19, 523–543. [Google Scholar] [CrossRef]
- Dyshlovoy, S.A.; Honecker, F. Marine Compounds and Cancer: The First Two Decades of XXI Century. Mar. Drugs. 2019, 18, 20. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.C.; Harris, J.L.; Khanna, K.K.; Hong, J.H. A Comprehensive Review on Current Advances in Peptide Drug Development and Design. Int. J. Mol. Sci. 2019, 20, 2383. [Google Scholar] [CrossRef] [Green Version]
- Rangel, M.; de Santana, C.J.; Pinheiro, A.; Dos Anjos, L.; Barth, T.; Pires, O.R.; Fontes, W.; Castro, M.S. Marine Depsipeptides as Promising Pharmacotherapeutic Agents. Curr. Protein Pept. Sci. 2017, 18, 72–91. [Google Scholar] [CrossRef]
- Le, T.C.; Pulat, S.; Lee, J.; Kim, G.J.; Kim, H.; Lee, E.Y.; Hillman, P.F.; Choi, H.; Yang, I.; Oh, D.C.; et al. Marine Depsipeptide nobilamide I Inhibits Cancer Cell Motility and Tumorigenicity via Suppressing Epithelial-Mesenchymal Transition and MMP2/9 Expression. ACS Omega 2022, 7, 1722–1732. [Google Scholar] [CrossRef] [PubMed]
- Al-Awadhi, F.H.; Paul, V.J.; Luesch, H. Structural Diversity and Anticancer Activity of Marine-Derived Elastase Inhibitors: Key Features and Mechanisms Mediating the Antimetastatic Effects in Invasive Breast Cancer. Chembiochem 2018, 19, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Wang, X.; Zhang, H.; Sun, J.; Zheng, L.; Liu, H.; Wang, J.; Shen, A.; Geng, M.; Guo, Y. Chromopeptide A, a highly cytotoxic depsipeptide from the marine sediment-derived bacterium Chromobacterium sp. HS-13-94. Acta Pharm. Sin. B 2015, 5, 62–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asfaha, Y.; Schrenk, C.; Alves Avelar, L.A.; Hamacher, A.; Pflieger, M.; Kassack, M.U.; Kurz, T. Recent advances in class IIa histone deacetylases research. Bioorg. Med. Chem. 2019, 27, 115087. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.Y.; Wang, J.D.; Wang, X.; Liu, H.C.; Zhang, M.M.; Liu, Y.C.; Zhang, C.H.; Su, Y.; Shen, Y.Y.; Guo, Y.W.; et al. Marine-derived chromopeptide A, a novel class I HDAC inhibitor, suppresses human prostate cancer cell proliferation and migration. Acta Pharmacol. Sin. 2017, 38, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Seto, E. HDACs and HDAC Inhibitors in Cancer Development and Therapy. Cold Spring Harb. Perspect. Med. 2016, 6, a026831. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Zhang, C.; Liu, F.; Ma, J.; Jia, F.; Han, Z.; Xie, W.; Li, X. Actinomycin V Inhibits Migration and Invasion via Suppressing Snail/Slug-Mediated Epithelial-Mesenchymal Transition Progression in Human Breast Cancer MDA-MB-231 Cells In Vitro. Mar. Drugs 2019, 17, 305. [Google Scholar] [CrossRef] [Green Version]
- Kgk, D.; Kumari, S.; Shailender, G.; Malla, R.R. Marine natural compound cyclo(L-leucyl-L-prolyl) peptide inhibits migration of triple negative breast cancer cells by disrupting interaction of CD151 and EGFR signaling. Chem Biol. Interact. 2020, 315, 108872. [Google Scholar] [CrossRef]
- Al-Awadhi, F.H.; Salvador, L.A.; Law, B.K.; Paul, V.J.; Luesch, H. Kempopeptin C, a Novel Marine-Derived Serine Protease Inhibitor Targeting Invasive Breast Cancer. Mar. Drugs 2017, 15, 290. [Google Scholar] [CrossRef] [Green Version]
- Al-Awadhi, F.H.; Law, B.K.; Paul, V.J.; Luesch, H. Grassystatins D-F, Potent Aspartic Protease Inhibitors from Marine Cyanobacteria as Potential Antimetastatic Agents Targeting Invasive Breast Cancer. J. Nat. Prod. 2017, 80, 2969–2986. [Google Scholar] [CrossRef]
- Suleria, H.A.; Masci, P.P.; Gobe, G.C.; Osborne, S.A. Therapeutic potential of abalone and status of bioactive molecules: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2017, 57, 1742–1748. [Google Scholar] [CrossRef] [PubMed]
- Gong, F.; Chen, M.F.; Chen, J.; Li, C.; Zhou, C.; Hong, P.; Sun, S.; Qian, Z.J. Boiled Abalone Byproduct Peptide Exhibits Anti-Tumor Activity in HT1080 Cells and HUVECs by Suppressing the Metastasis and Angiogenesis In Vitro. J. Agric. Food Chem. 2019, 67, 8855–8867. [Google Scholar] [CrossRef] [PubMed]
- Gong, F.; Chen, M.-F.; Zhang, Y.-Y.; Li, C.-Y.; Zhou, C.-X.; Hong, P.-Z.; Sun, S.-L.; Qian, Z.-J. A Novel Peptide from Abalone (Haliotis discus hannai) to Suppress Metastasis and Vasculogenic Mimicry of Tumor Cells and Enhance Anti-Tumor Effect In Vitro. Mar. Drugs 2019, 17, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Zhangsun, D.; Harvey, P.J.; Kaas, Q.; Wu, Y.; Zhu, X.; Hu, Y.; Li, X.; Tsetlin, V.I.; Christensen, S.; et al. Cloning, synthesis, and characterization of αO-conotoxin GeXIVA, a potent α9α10 nicotinic acetylcholine receptor antagonist. Proc. Natl. Acad. Sci USA 2015, 112, E4026–E4035. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Bao, J.; Zhangsun, M.; Dong, S.; Zhangsun, D.; Luo, S. αO-Conotoxin GeXIVA Inhibits the Growth of Breast Cancer Cells via Interaction with α9 Nicotine Acetylcholine Receptors. Mar. Drugs 2020, 18, 195. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Cui, J.; Zhang, R.; Wang, Y.; Hong, M. A novel fibrinolytic serine protease from the polychaete Nereis (Neanthes) virens (Sars): Purification and characterization. Biochimie 2007, 89, 93–103. [Google Scholar] [CrossRef]
- Chen, Y.; Tang, Y.; Tang, Y.; Yang, Z.; Ding, G. Serine Protease from Nereis virens Inhibits H1299 Lung Cancer Cell Proliferation via the PI3K/AKT/mTOR Pathway. Mar. Drugs 2019, 17, 366. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Zhu, X.; Yang, K.; Zhu, M.; Farooqi, A.A.; Kang, D.; Sun, M.; Xu, Y.; Lin, X.; Feng, Y.; et al. PBN11-8, a Cytotoxic Polypeptide Purified from Marine bacillus, Suppresses Invasion and Migration of Human Hepatocellular Carcinoma Cells by Targeting Focal Adhesion Kinase Pathways. Polymers 2018, 10, 1043. [Google Scholar] [CrossRef] [Green Version]
- Ru, R.; Guo, Y.; Mao, J.; Yu, Z.; Huang, W.; Cao, X.; Hu, H.; Meng, M.; Yuan, L. Cancer Cell Inhibiting Sea Cucumber (Holothuria leucospilota) Protein as a Novel Anti-Cancer Drug. Nutrients 2022, 14, 786. [Google Scholar] [CrossRef]
- Hussain, G.; Huang, J.; Rasul, A.; Anwar, H.; Imran, A.; Maqbool, J.; Razzaq, A.; Aziz, N.; Makhdoom, E.u.H.; Konuk, M.; et al. Putative Roles of Plant-Derived Tannins in Neurodegenerative and Neuropsychiatry Disorders: An Updated Review. Molecules 2019, 24, 2213. [Google Scholar] [CrossRef] [Green Version]
- Briguglio, G.; Costa, C.; Pollicino, M.; Giambò, F.; Catania, S.; Fenga, C. Polyphenols in cancer prevention: New insights (Review). Int. J. Funct. Nutr. 2020, 1, 9. [Google Scholar] [CrossRef]
- Montero, L.; Del Pilar Sánchez-Camargo, A.; Ibáñez, E.; Gilbert-López, B. Phenolic Compounds from Edible Algae: Bioactivity and Health Benefits. Curr. Med. Chem. 2018, 25, 4808–4826. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Liu, Y.; Xiao, Z.; Tang, Y.; Hong, P.; Sun, S.; Zhou, C.; Qian, Z.J. Inhibition effects of 7-phloro-eckol from Ecklonia cava on metastasis and angiogenesis induced by hypoxia through regulation of AKT/mTOR and ERK signaling pathways. Arab. J. Chem. 2021, 14, 103187. [Google Scholar] [CrossRef]
- Lee, Y.J.; Park, J.H.; Park, S.A.; Joo, N.R.; Lee, B.H.; Lee, K.B.; Oh, S.M. Dieckol or Phlorofucofuroeckol extracted from Ecklonia cava suppresses lipopolysaccharide-mediated human breast cancer cell migration and invasion. J. Appl. Phycol. 2020, 32, 631–640. [Google Scholar] [CrossRef]
- Vitale, G.A.; Coppola, D.; Palma Esposito, F.; Buonocore, C.; Ausuri, J.; Tortorella, E.; de Pascale, D. Antioxidant Molecules from Marine Fungi: Methodologies and Perspectives. Antioxidants 2020, 9, 1183. [Google Scholar] [CrossRef] [PubMed]
- Vasarri, M.; Vitale, G.A.; Varese, G.C.; Barletta, E.; D’Auria, M.V.; de Pascale, D.; Degl’Innocenti, D. Dihydroauroglaucin Isolated from the Mediterranean Sponge Grantia compressa Endophyte Marine Fungus Eurotium chevalieri Inhibits Migration of Human Neuroblastoma Cells. Pharmaceutics 2022, 14, 616. [Google Scholar] [CrossRef]
- Liu, J.; Li, F.M.; La Kim, E.; Hong, J.K.; Jung, J.H. Viriditoxin, from a Jellyfish-derived Fungus, is Antibiotic to Fish Pathogens. Nat. Prod. Sci. 2013, 19, 61–65. [Google Scholar]
- Su, M.; Zhao, C.; Li, D.; Cao, J.; Ju, Z.; Kim, E.L.; Jung, Y.-S.; Jung, J.H. Viriditoxin Stabilizes Microtubule Polymers in SK-OV-3 Cells and Exhibits Antimitotic and Antimetastatic Potential. Mar. Drugs 2020, 18, 445. [Google Scholar] [CrossRef]
- Chojnacka, K.; Lewandowska, U. Chemopreventive effects of polyphenol-rich extracts against cancer invasiveness and metastasis by inhibition of type IV collagenases expression and activity. J. Funct. Foods 2018, 46, 295–311. [Google Scholar] [CrossRef]
- Rajamani, K.; Thirugnanasambandan, S.S. Polyphenols from brown alga, Padina boergesenii (Allendar & Kraft) decelerates renal cancer growth involving cell cycle arrest and induction of apoptosis in renal carcinoma cells. Environ. Toxicol. 2018, 33, 1135–1142. [Google Scholar] [CrossRef]
- Begum, S.M.F.M.; Chitra, K.; Joseph, B.; Sundararajan, R.; Hemalatha, S. Gelidiella acerosa inhibits lung cancer proliferation. BMC Complement Altern. Med. 2018, 18, 104. [Google Scholar] [CrossRef] [Green Version]
- Leri, M.; Ramazzotti, M.; Vasarri, M.; Peri, S.; Barletta, E.; Pretti, C.; Degl’Innocenti, D. Bioactive Compounds from Posidonia oceanica (L.) Delile Impair Malignant Cell Migration through Autophagy Modulation. Mar. Drugs 2018, 16, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasarri, M.; Leri, M.; Barletta, E.; Pretti, C.; Degl’Innocenti, D. Posidonia oceanica (L.) Delile Dampens Cell Migration of Human Neuroblastoma Cells. Mar. Drugs 2021, 19, 579. [Google Scholar] [CrossRef] [PubMed]
- Piazzini, V.; Vasarri, M.; Degl’Innocenti, D.; Guastini, A.; Barletta, E.; Salvatici, M.C.; Bergonzi, M.C. Comparison of Chitosan Nanoparticles and Soluplus Micelles to Optimize the Bioactivity of Posidonia oceanica Extract on Human Neuroblastoma Cell Migration. Pharmaceutics 2019, 11, 655. [Google Scholar] [CrossRef] [Green Version]
- Tohme, R.; Darwiche, N.; Gali-Muhtasib, H. A journey under the sea: The quest for marine anti-cancer alkaloids. Molecules 2011, 16, 9665–9696. [Google Scholar] [CrossRef]
- Bechmann, N.; Ehrlich, H.; Eisenhofer, G.; Ehrlich, A.; Meschke, S.; Ziegler, C.G.; Bornstein, S.R. Anti-Tumorigenic and Anti-Metastatic Activity of the Sponge-Derived Marine Drugs Aeroplysinin-1 and Isofistularin-3 against Pheochromocytoma In Vitro. Mar. Drugs 2018, 16, 172. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.-C.; Kuo, T.-T.; Chang, H.-Y.; Liu, W.-S.; Hsia, S.-M.; Huang, T.-C. Manzamine A Exerts Anticancer Activity against Human Colorectal Cancer Cells. Mar. Drugs 2018, 16, 252. [Google Scholar] [CrossRef] [Green Version]
- Charupant, K.; Suwanborirux, K.; Amnuoypol, S.; Saito, E.; Kubo, A.; Saito, N. Jorunnamycins A-C, new stabilized renieramycin-type bistetrahydroisoquinolines isolated from the Thai nudibranch Jorunna funebris. Chem. Pharm. Bull. 2007, 55, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Ecoy, G.A.U.; Chamni, S.; Suwanborirux, K.; Chanvorachote, P.; Chaotham, C. Jorunnamycin A from Xestospongia sp. Suppresses Epithelial to Mesenchymal Transition and Sensitizes Anoikis in Human Lung Cancer Cells. J. Nat. Prod. 2019, 82, 1861–1873. [Google Scholar] [CrossRef]
- Tabakmakher, K.M.; Makarieva, T.N.; Denisenko, V.A.; Popov, R.S.; Kuzmich, A.S.; Shubina, L.K.; Lee, H.-S.; Lee, Y.-J.; Fedorov, S.N. Normonanchocidins G and H, New Pentacyclic Guanidine Alkaloids from the Far-Eastern Marine Sponge Monanchora pulchra. Nat. Prod. Commun. 2017, 12, 7. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polysaccharides | Marine Source | Composition | In Vivo Animal Model | Target |
|---|---|---|---|---|
| Exopolysaccharide 11 | Bacillus sp. 11 | Mannose, glucosamine, galacturonic acid, glucose and xylose, in a ratio of 1:2.58:0.68:0.13:3.09:1.41 | Melanoma (mice model) | Filiform structures; cytoskeleton; CD99 [41] |
| Polysaccharides | Marine source | Composition | In vitro cell model | Target |
| Exopolysaccharide 11 | Bacillus sp. 11 | Mannose, glucosamine, galacturonic acid, glucose and xylose, in a ratio of 1:2.58:0.68:0.13:3.09:1.41 | Human lung cancer (A549 cells); human hepatoma (Huh7.5, HepG2, Bel-7402 cells) | Filiform structures; cytoskeleton; CD99 [40,41,42] |
| Fucoidan-like STPC2 | S. thunbergii | Fucose, xylose, galactose and glucuronic acid, in a ratio of 8.1:3.8:2.1:1.0 | Human lung cancer (A549 cells) | MMP-2 [43] |
| Laminaran sulfate | F. evanescens | β-(1→3)-linked D-glucopyranose and single β-(1→6)-linked D-glucose residues. Branches at C6 can be glucose or gentiobiose. | Human colorectal adenocarcinoma (HCT116 cells); human malignant melanoma (SK-MEL-5 cells); human breast adenocarcinoma (MDA-MB-231 cells) | MMP-2/9 [44] |
| Laminaran sulfate | D. dichotoma | (1→3)-linked glucopyranose residues with the branches of single glucose residues at C6 (ratio of bonds 1.3:1.6 = 3:1) | Human melanoma (SK-MEL-28 cells) | MMP-2/9 [45] |
| SPUP | U. Pinnatifida | Fucose, glucose, and galactose, in a ratio of 27.22:19.32:53.46 | Human breast cancer (MCF-7 cells); human ovarian cancer (SKOV-3 and A2780 cells) | Apoptosis [46]; Hedgehog signaling pathway [47] |
| BFP-3 | B. fuscopurpurea | Rhamnose, arabinose, mannose, glucose, and galactose | Human ovarian cancer (A2780 cells) | Apoptosis, autophagy [49] |
| Fucoidan | C. frondosa | L-fucose and a sulfate ester group. Total carbohydrate content 72.5%; sulfate content 26.1%. (98% purity). | Human osteosarcoma (U2OS cells) | Cytoskeleton; FAK [50] |
| SIP-SII | S. maindroni | Fucose, N-acetylgalactosamine and mannose, in a molar ratio of 2:2:1, with a single branch of glucuronic acid at the C-3 position of mannose. | Human epidermoid carcinoma (KB cells); human ovarian carcinoma (SKOV-3 cells) | EGF-mediated signaling pathway; MMP-2 [51,52] |
| Sulfated galactans | G. fisheri | 3-linked β-d-galactopyranose and 4-linked 3,6-anhydro-α-l-galactopyranose or α-l-galactose-6-sulfate with partial methylation at 2-O-methylated-3,6-andydro-α-l-galactopyranose, 6-O-methylated-β-d-galactopyranose and 4-O-methyl-β-l-galactopyranose attached to C-6 of 3-linked-β-d-galactopyranose units, together with sulfation on C-4 and C-6 of d-galactopyranose units. | Human intrahepatic cholangiocarcinoma (HuCCA-1 cells) | EGFR-ERK signaling pathway; FAK; E-cadherin [54,55] |
| CCB-F0.5 | C. cupressoides var. flabellata | Galactose and mannose in a molar ratio of 1.0:0.1 and traces of xylose. Low protein content 0.17% | Murine melanoma (B16-F10 cells) | Not described [56] |
| CCB-F1.0 | C. cupressoides var. flabellata | Galactose, mannose, and xylose in a molar ratio of 1.0:0.1:0.6 and traces of glucose and rhamnose. Low protein content 0.19% | Murine melanoma (B16-F10 cells) | Not described [56] |
| Crude sulfate polysaccharide extract | P. tetrastromatica | - | Human cervical adenocarcinoma (HeLa cells) | MMP-2/9; angiogenesis [57] |
| Depsipeptides | Marine Source | In Vitro Cell Model | Target |
|---|---|---|---|
| Nobilamide I | Saccharomonospora sp., strain CNQ-490 | Human lung cancer (A549 cells); human gastric cancer (AGS cells); human colorectal cancer (Caco2 cells) | MMP-2/9 [62] |
| Molassamide | Cyanobacterium DRTO-73, Leptolyngbya sp. | Human breast adenocarcinoma (MDA-MB-231 cells) | ICAM-1 [63] |
| Chromopeptide A | Chromobacterium sp., strain HS-13-94 | Human prostate cancer (PC3 cells); mouse PC3 xenograft model | HDACs [66] |
| Actinomycin V | Streptomyces sp., strain N1510.2 | Human breast adenocarcinoma (MDA-MB-231 cells) | EMT [68] |
| CLP | Marine microorganisms | Human breast adenocarcinoma (MDA-MB-231 and MDA-MB-468 cells) | CD151/EGFR signaling [69] |
| Kempopeptin C | Lyngbya sp. | Human breast adenocarcinoma (MDA-MB-231) | Not described [70] |
| Grassystatin F | Cyanobacterium VPG 14-61 | Human breast adenocarcinoma (MDA-MB-231) | Protease inhibitor [71] |
| Peptides and Proteins | Marine Source | Sequence | Molecular Mass | In Vitro Cell Model | Target |
|---|---|---|---|---|---|
| BABP | H. discus hannai | EMDEAQDPSEW | 1234.41 Da | Human fibrosarcoma (HT1080 cells); human umbilical vein endothelial (HUVEC cells) | MMPs; VEGF [73] |
| AATP | H. discus hannai | KVEPQDPSEW | 1214.30 Da | Fibrosarcoma (HT1080 cells); human umbilical vein endothelial (HUVEC cells) | MMPs; VEGF [74] |
| αO-conotoxin GeXIVA | C. generalis | TCRSSGRYCRSPYDRRRRYCRRITDACV 1 | 3452 Da | Human breast cancer (MDA-MB-157 cells) | EMT [76] |
| N-V protease (or NAP) | N. virens | QAPNYSTASYNVVAVKINLFLSTNNKLYIHDTGVRAVYLAGMKVYLAANPTASSQTFNSDTLVYILDTGINEPNYYINLY 2 | 8888 Da | Human lung cancer (H1299 cells) | apoptosis [78] |
| PBN11-8 | B. pumilus | ASTGSQKVTVYAVAD 3 | 19,000 Da | Human hepatocellular carcinoma (BEL-7402 cells) | FAK; MMP-2/9 [79] |
| Phenolic Compound | Marine Source | Chemical Structure | In Vitro Cell Model | Target |
|---|---|---|---|---|
| 7-phloroeckol | E. cava |  | Human hepatoma cancer (HepG2 cells); human umbilical vein endothelial (HUVEC cells) | HIF-1α; angiogenesis [84] |
| Dieckol | E. cava |  | Human breast cancer (MCF-7, MDA-MB-231 cells) | MMP-2/9; TLR-4; NF-κB [85] |
| Phlorofucofuroeckol | E. cava |  | Human breast cancer (MCF-7, MDA-MB-231 cells) | MMP-2/9; TLR-4; NF-κB [85] |
| Dihydroauroglaucin | E. chevalieri |  | Human neuroblastoma (SH-SY5Y cells) | Autophagy [87] |
| Viriditoxin | P. variotii |  | Human ovarian cancer (SKOV-3 cells) | Cytoskeleton [89] |
| Gallic acid | P. boergeseni, P. oceanica |  | Human renal cancer (A498, ACHN cells); human fibrosarcoma (HT1080 cells); human neuroblastoma (SH-SY5Y cells) | Apoptosis [91]; autophagy [93]; autophagy, MMP-2 [94] |
| Caffeic acid | P. boergeseni |  | Human renal cancer (A498, ACHN cells) | Apoptosis [91] |
| Rutin | P. boergeseni |  | Human renal cancer (A498, ACHN cells) | Apoptosis [91] |
| Quercetin | P. boergeseni |  | Human renal cancer (A498, ACHN cells) | Apoptosis [91] |
| Ferulic acid | P. boergeseni, P. oceanica |  | Human renal cancer (A498, ACHN cells); human fibrosarcoma (HT1080 cells); human neuroblastoma (SH-SY5Y cells) | Apoptosis [91]; autophagy [93]; autophagy, MMP-2 [94] |
| Catechin | P. oceanica |  | Human fibrosarcoma (HT1080 cells); human neuroblastoma (SH-SY5Y cells) | Autophagy [93]; autophagy, MMP-2 [94] |
| Epicatechin | P. oceanica |  | Human fibrosarcoma (HT1080 cells); human neuroblastoma (SH-SY5Y cells) | Autophagy [93]; autophagy, MMP-2 [94] |
| Chlorogenic acid | P. oceanica |  | Human fibrosarcoma (HT1080 cells); human neuroblastoma (SH-SY5Y cells) | Autophagy [93]; autophagy, MMP-2 [94] |
| Alkaloids | Marine Source | In Vitro Cell Model | Target |
|---|---|---|---|
| Aeroplysinin-1 | A. aerophoba | Mouse pheocromocytoma cells (MTT cells) | Integrin β1 [97] |
| Manzamine A | Haliclona sp., Xestospongia sp. and Pellina sp. | Human colorectal carcinoma (HCT116 cells) | EMT [98] |
| Jorunnamycin A | Xestospongia sp. | Human lung cancer (H460 cells) | EMT; anoikis [100] |
| Normonanchocidines G and H | M. pulchra | Human cervical carcinoma (HeLa cells) | Not described [101] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasarri, M.; Barletta, E.; Degl’Innocenti, D. Marine Migrastatics: A Comprehensive 2022 Update. Mar. Drugs 2022, 20, 273. https://doi.org/10.3390/md20050273
Vasarri M, Barletta E, Degl’Innocenti D. Marine Migrastatics: A Comprehensive 2022 Update. Marine Drugs. 2022; 20(5):273. https://doi.org/10.3390/md20050273
Chicago/Turabian StyleVasarri, Marzia, Emanuela Barletta, and Donatella Degl’Innocenti. 2022. "Marine Migrastatics: A Comprehensive 2022 Update" Marine Drugs 20, no. 5: 273. https://doi.org/10.3390/md20050273
APA StyleVasarri, M., Barletta, E., & Degl’Innocenti, D. (2022). Marine Migrastatics: A Comprehensive 2022 Update. Marine Drugs, 20(5), 273. https://doi.org/10.3390/md20050273