
Exopolysaccharides from Microalgae and Cyanobacteria: Diversity of Strains, Production Strategies, and Applications

Abstract
:1. Introduction
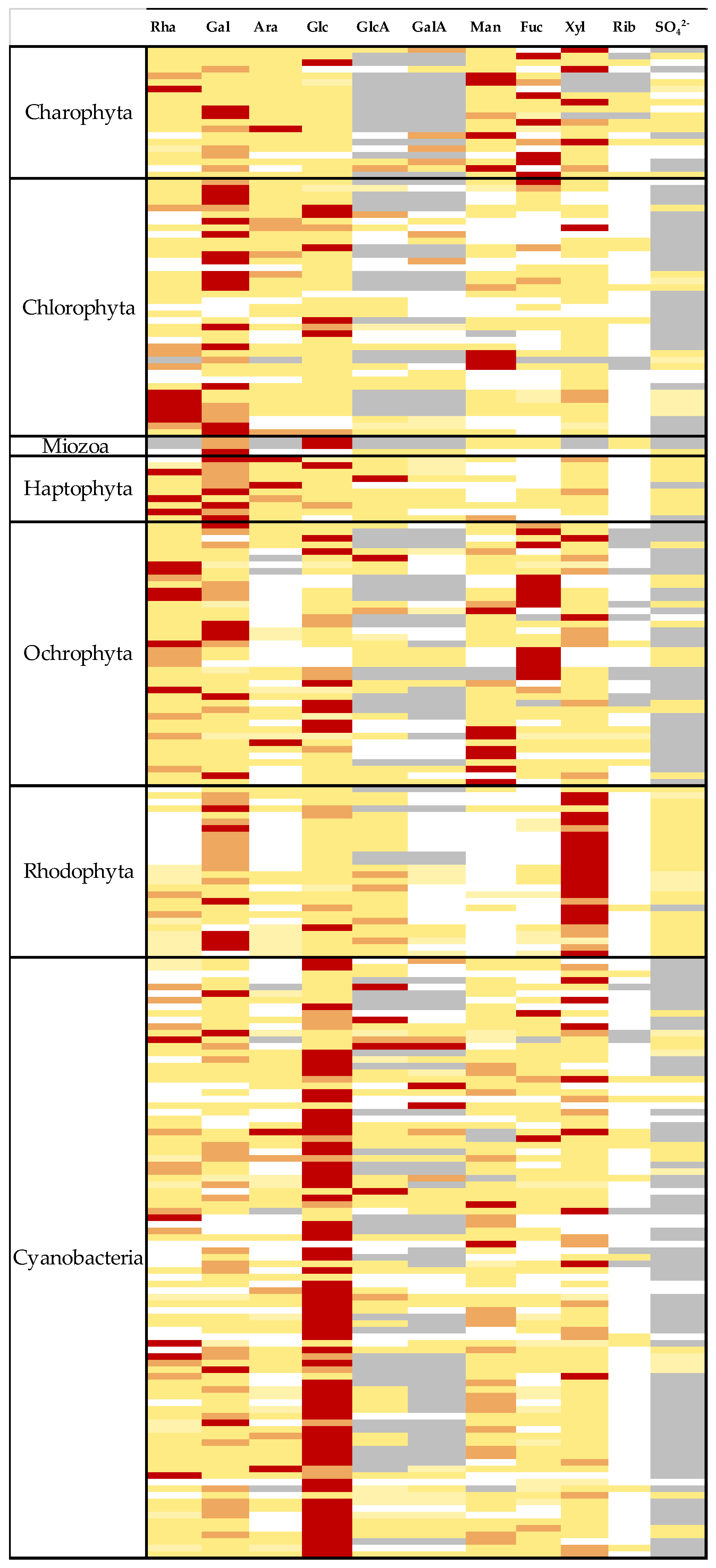
2. Diversity of Producing Strains and Structures

{kind=link}
{kind=link}
{kind=link}
| Genera | Species | Rha | Gal | Ara | Glc | GlcA | GalA | Man | Fuc | Xyl | Rib | SO42− | Proteins | Other Informations | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Closterium | sp. | [35] | |||||||||||||
| Cosmarium | sp. | Uronic acids | [33] | ||||||||||||
| Cosmarium | sp. | [33] | |||||||||||||
| Hyalotheca | dissiliens | [68] | |||||||||||||
| Klebsormidium | flaccidum (749B) | Uronic acids | [34] | ||||||||||||
| flaccidum (748A) | Uronic acids | ||||||||||||||
| flaccidum (446C) | Uronic acids | ||||||||||||||
| Micrasterias | denticulata | [69] | |||||||||||||
| Netrium | digitus | Uronic acids | [33] | ||||||||||||
| interruptum | Uronic acids | ||||||||||||||
| Interruptum UTEX 2509 | |||||||||||||||
| oblongum | Uronic acids | ||||||||||||||
| Penium | cylindrus | Uronic acids | [33] | ||||||||||||
| spirostriolatum | Uronic acids | ||||||||||||||
| margaritaceum | Methyl groups | [70] | |||||||||||||
| Pleurotaenium | trabecula | Uronic acids | [33] | ||||||||||||
| Spondylosium | panduriforme | [71] | |||||||||||||
| Staurastrum | iversenii | [72] | |||||||||||||
| orbiculare | [48] | ||||||||||||||
| Stauroneis | sp. | GlcN | [37] | ||||||||||||
| Tetmemorus | brebissonii | Uronic acids | [33] |

| Genera | Species | Rha | Gal | Ara | Glc | GlcA | GalA | Man | Fuc | Xyl | Rib | SO42− | Proteins | Other Informations | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ankistrodesmus | densus | [73] | |||||||||||||
| Botryococcus | braunii | [74] | |||||||||||||
| braunii | Methyl groups | [75] | |||||||||||||
| braunii | Uronic acids, methyl groups | [76] | |||||||||||||
| Bracteacoccus | sp. | Uronic acids | [34] | ||||||||||||
| Chlamydomonas | agustae | [77] | |||||||||||||
| corrosa | |||||||||||||||
| humicola | [78] | ||||||||||||||
| peterfii | |||||||||||||||
| reinhardtii | Pyruvate | [65] | |||||||||||||
| mexicana | Uronic acids | [79] | |||||||||||||
| sajao | Uronic acids | ||||||||||||||
| sajao | [78] | ||||||||||||||
| stigmatophora | [80] | ||||||||||||||
| sp. | [81] | ||||||||||||||
| Chlorella | autotrophica | [82] | |||||||||||||
| mirabilis (678F) | Uronic acids | [34] | |||||||||||||
| mirabilis (7410G) | Uronic acids | ||||||||||||||
| ellipsoidea | Uronic acids | ||||||||||||||
| pyrenoidosa | [83] | ||||||||||||||
| vulgaris | [81] | ||||||||||||||
| ellipsoidea | |||||||||||||||
| sp. | |||||||||||||||
| sp. | [25] | ||||||||||||||
| Chlorococcum | sp. | [84] | |||||||||||||
| Coccomyxa | sp. | [25] | |||||||||||||
| Desmococcus | olivaceus | Methyl groups | [31] | ||||||||||||
| Dunaliella | bardawil | [82] | |||||||||||||
| tertiolecta | [85] | ||||||||||||||
| salina | [86] | ||||||||||||||
| sp. | [25] | ||||||||||||||
| Geminella | terricola | Uronic acids | [34] | ||||||||||||
| Haematococcus | lacustris | [87] | |||||||||||||
| Heterosigma | akashiwo | ManA | [43] | ||||||||||||
| Oocystis | sp. | [81] | |||||||||||||
| Palmella | mucosa | ||||||||||||||
| Prasinococcus | sp. | [25] | |||||||||||||
| Scenedesmus | quadricauda | [88] | |||||||||||||
| Stichococcus | bacillaris (772B, 747C) | Uronic acids | [34] | ||||||||||||
| bacillaris (774E, 677A) | Uronic acids | ||||||||||||||
| Tetraselmis | chui | [25] | |||||||||||||
| globosa | |||||||||||||||
| rubens | |||||||||||||||
| sp. |
| Genera | Species | Rha | Gal | Ara | Glc | GlcA | GalA | Man | Fuc | Xyl | Rib | SO42− | Proteins | Other Informations | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Amphidinium | carterae | [25] | |||||||||||||
| Cochlodinium | polykrikoides | [89] | |||||||||||||
| Crypthecodinium | cohnii | [90] | |||||||||||||
| Gyrodinium | impudicum | [30] | |||||||||||||
| Bigelowiela | natans | [25] |
| Genera | Species | Rha | Gal | Ara | Glc | GlcA | GalA | Man | Fuc | Xyl | Rib | SO42− | Proteins | Other Informations | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Calyptrosphaera | sp. | [25] | |||||||||||||
| Chrysotila | dentata | [25] | |||||||||||||
| sp. | [25] | ||||||||||||||
| Diacronema | sp. | [25] | |||||||||||||
| Emiliania | huxleyi | [25] | |||||||||||||
| Exanthemachrysis | sp. | [25] | |||||||||||||
| sp. | [25] | ||||||||||||||
| Hymenomonas | coronata | [25] | |||||||||||||
| Isochrysis | braarudii | [25] | |||||||||||||
| Ochrosphaera | verrucosa | [25] | |||||||||||||
| sp. | [25] | ||||||||||||||
| Pavlova | enorae | [25] | |||||||||||||
| gyrans | [25] | ||||||||||||||
| sp. | [25] | ||||||||||||||
| sp. | [25] | ||||||||||||||
| sp. | [25] | ||||||||||||||
| sp. | [25] | ||||||||||||||
| Prymnesium | parvum | [25] | |||||||||||||
| Rebecca | sp. | [25] | |||||||||||||
| Ruttnera | lamellosa | [25] | |||||||||||||
| PLY 431 | [25] | ||||||||||||||
| RCC 3704 | [25] |
| Genera | Species | Rha | Gal | Ara | Glc | GlcA | GalA | Man | Fuc | Xyl | Rib | SO42− | Proteins | Other informations | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Achnanthes | longipes | [91] | |||||||||||||
| Amphora | coffeaeformis | ||||||||||||||
| holsatica | Uronic acids | [92] | |||||||||||||
| rostrata | Pyruvate | [66] | |||||||||||||
| salina | GlcNAc | [47] | |||||||||||||
| sp. | GlcN, traces GlcNAc | [37] | |||||||||||||
| Asterionella | socialis | [93] | |||||||||||||
| Aulacoseira | granulata | GlcNAc | [38] | ||||||||||||
| Chaetoceros | affinis | [94] | |||||||||||||
| curvisetus | [95] | ||||||||||||||
| decipiens | [96] | ||||||||||||||
| decipiens | [97] | ||||||||||||||
| Coscinodiscus | nobilis | [98] | |||||||||||||
| radiatus | [47] | ||||||||||||||
| Cylindrotheca | closterium | Uronic acids | [99] | ||||||||||||
| fusiformis | Uronic acids | [28] | |||||||||||||
| Cymbella | cistula | [91] | |||||||||||||
| mexicana | |||||||||||||||
| Cyclotella | nana | [93] | |||||||||||||
| Glossomastix | sp. | [25] | |||||||||||||
| sp. | |||||||||||||||
| sp. | |||||||||||||||
| Melosira | nummuloides | Uronic acids | [92] | ||||||||||||
| Navicula | directa | Uronic acids | [92] | ||||||||||||
| curvilineata | [47] | ||||||||||||||
| incerta | [93] | ||||||||||||||
| phyllepta | [100] | ||||||||||||||
| salinarum | Uronic acids | [99] | |||||||||||||
| subinflata | Uronic acids, pyruvate, methyl groups | [67] | |||||||||||||
| Nitzschia | angularis | Other main sugar | [93] | ||||||||||||
| closterium | [85] | ||||||||||||||
| epithemoides | [100] | ||||||||||||||
| frustulum | [93] | ||||||||||||||
| Pelagococcus | sp. | [25] | |||||||||||||
| Phaeodactylum | tricornutum CCMP632 ovoid | Methyl groups | [101] | ||||||||||||
| tricornutum CCMP632 fusiform | Methyl groups | ||||||||||||||
| tricornutum | [85] | ||||||||||||||
| Phaeomonas | sp. | [25] | |||||||||||||
| Pinnularia | viridis | Uronic acids, methyl groups | [102] | ||||||||||||
| Thalassiosira | pseudonana | [103] | |||||||||||||
| sp. | GlcNAc, GalNAc, | [48] | |||||||||||||
| Tribonema | Sp. | [104] | |||||||||||||
| Triceratium | dubium | GlcNAc | [47] |
| Genera | Species | Rha | Gal | Ara | Glc | GlcA | GalA | Man | Fuc | Xyl | Rib | SO42− | Proteins | Other Informations | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dixionella | grisea | Uronic acids, methyl groups | [11] | ||||||||||||
| Flintiella | sanguinaria | Methyl and acetyl groups | [59] | ||||||||||||
| Porphyridium | aerugineum | Methyl groups | [58] | ||||||||||||
| cruentum | [105] | ||||||||||||||
| cruentum | Methyl groups | [58] | |||||||||||||
| marinum | [13] | ||||||||||||||
| purpureum | [106] | ||||||||||||||
| purpureum | [107] | ||||||||||||||
| sordidum | Methyl groups | [107] | |||||||||||||
| sordidum | [32] | ||||||||||||||
| sp. | Uronic acids, methyl groups | [62] | |||||||||||||
| sp. | [108] | ||||||||||||||
| Timspurckia | oligopyrenoides | [32] | |||||||||||||
| Erythrolobus | coxiae | [32] | |||||||||||||
| madagascarensis | [32] | ||||||||||||||
| Corynoplastis | japonica | [32] | |||||||||||||
| Rhodella | grisea | Methyl groups | [61] | ||||||||||||
| maculata | [106] | ||||||||||||||
| reticulata | [109] | ||||||||||||||
| violacea | [12] | ||||||||||||||
| Neorhodella | cyanea | [32] | |||||||||||||
| Chroodactylon | ornatum | [32] | |||||||||||||
| Chroothece | richteriana | [32] | |||||||||||||
| Bangiopsis | subsimplex | [32] | |||||||||||||
| Rhodaphanes | brevistipitata | [32] | |||||||||||||
| Rhodospora | sordida | [32] |
| Genera | Species | Rha | Gal | Ara | Glc | GlcA | GalA | Man | Fuc | Xyl | Rib | SO42− | Proteins | Other Informations | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Anabaena | augstmalis | GalN | [46] | ||||||||||||
| cylindrica | Other uronic acid | [44] | |||||||||||||
| flos-aquae | [81] | ||||||||||||||
| sp. | Uronic acids, pyruvate | [110] | |||||||||||||
| spiroides | GlcNAc, GalNAc | [38] | |||||||||||||
| sphaerica | [49] | ||||||||||||||
| torulosa | GlcN | ||||||||||||||
| Anacystis | nidulans | [111] | |||||||||||||
| Aphanocapsa | halophytia | [112] | |||||||||||||
| Aphanothece | halophytica | [39] | |||||||||||||
| Arthrospira | maxima | [113] | |||||||||||||
| platensis | [114] | ||||||||||||||
| platensis | [53] | ||||||||||||||
| platensis | Methyl groups | [40] | |||||||||||||
| Calothrix | pulvinata | Uronic acids | [34] | ||||||||||||
| sp. | GlcN | [46] | |||||||||||||
| Chlorogloeopsis | sp. | [49] | |||||||||||||
| Chroococcus | minutus | GlcN, methyl groups | [50] | ||||||||||||
| submarinus | GlcN | [45] | |||||||||||||
| Cyanospira | capsulata | [41] | |||||||||||||
| Cyanothece | sp. | [115] | |||||||||||||
| sp. | [116] | ||||||||||||||
| Fischerella | maior | [42] | |||||||||||||
| muscicola | [49] | ||||||||||||||
| Geitlerinema | sp. | [45] | |||||||||||||
| Gloeocapsa | gelatinosa | [117] | |||||||||||||
| kuetzingigiana | GalN, traces GlcN | [51] | |||||||||||||
| sp. | [118] | ||||||||||||||
| Gloeocapsosis | crepidinum | GlcN, GalN | [51] | ||||||||||||
| Gloeothece | PCC6909 | [119] | |||||||||||||
| sp. | Methyl groups | [56] | |||||||||||||
| Johannesbaptistia | pellucida | [45] | |||||||||||||
| Leptolyngbya | foveolarum | Uronic acids | [34] | ||||||||||||
| tenuis | Uronic acids | ||||||||||||||
| sp. | GalN, traces GlcN | [51] | |||||||||||||
| Lyngbya | conferviodes | [120] | |||||||||||||
| Mastidocladus | laminosus | [121] | |||||||||||||
| Microcoleus | vaginatus | [55] | |||||||||||||
| vaginatus | GlcNAc, methyl groups | [31] | |||||||||||||
| Microcystis | aeruginosa | GlcNAc, GalNAc | [38] | ||||||||||||
| aeruginosa | [122] | ||||||||||||||
| aeruginosa flos-aquae | |||||||||||||||
| viridis | |||||||||||||||
| Nostoc | calcicola | [78] | |||||||||||||
| carneum | [116] | ||||||||||||||
| commune | [123] | ||||||||||||||
| commune | [124] | ||||||||||||||
| flagelliforme | [123] | ||||||||||||||
| sp. | [54] | ||||||||||||||
| sp. | [81] | ||||||||||||||
| sp. | Methyl groups | [125] | |||||||||||||
| insulare | Methyl groups | [60] | |||||||||||||
| insulare | [50] | ||||||||||||||
| muscoru | [118] | ||||||||||||||
| entophytum | |||||||||||||||
| sp. | [46] | ||||||||||||||
| verrucosum | [126] | ||||||||||||||
| Oscillatoria | amphibia | [120] | |||||||||||||
| corallinae | [120] | ||||||||||||||
| planktothrix FP1 | [127] | ||||||||||||||
| sp. | [49] | ||||||||||||||
| sp. | [116] | ||||||||||||||
| Phormidium | autumnale | [46] | |||||||||||||
| battersii | [45] | ||||||||||||||
| ambiguum | Uronic acids | [34] | |||||||||||||
| corium (442D, 746B) | Uronic acids | ||||||||||||||
| corium (444A) | Uronic acids | ||||||||||||||
| corium (674A) | Uronic acids | ||||||||||||||
| corium (743D) | Uronic acids | ||||||||||||||
| ectocarpi (K5) | [120] | ||||||||||||||
| ectocarpi (ME3) | |||||||||||||||
| ectocarpi (N182, C86) | |||||||||||||||
| joveolarum (C52) | |||||||||||||||
| joveolarum (MEU) | |||||||||||||||
| minutum (D5) | |||||||||||||||
| minutum (NB5) | |||||||||||||||
| minutum (RT6) | |||||||||||||||
| sp. (CCAP1464) | |||||||||||||||
| sp. (PNG91, 90-14/1) | |||||||||||||||
| sp. | [49] | ||||||||||||||
| tenue | GlcNAc | [31] | |||||||||||||
| tenue | GlcNAc | [125] | |||||||||||||
| golenkinianum | [45] | ||||||||||||||
| sp. | GlcN, traces GalN | [51] | |||||||||||||
| Rhabdoderma | rubrum | GlcN | [45] | ||||||||||||
| javanicum | Methyl groups | [31] | |||||||||||||
| javanicum | [125] | ||||||||||||||
| Synechococcus | sp. | [128] | |||||||||||||
| sp. | [25] | ||||||||||||||
| Synechocystis | aquatilis | GlcN | [46] | ||||||||||||
| sp. | [118] | ||||||||||||||
| sp. (PCC6803) | GlcN, GalN | [52] | |||||||||||||
| sp. (PCC6714) | GlcN, GalN | ||||||||||||||
| Tolypothrix | tenuis | GlcN | [49] | ||||||||||||
| Trichormus | variabilis (VRUC162) | GalN | [46] | ||||||||||||
| variabilis (VRUC168) | GlcN |
3. Culture Strategies for EPS Production and Impact on Composition
3.1. Nutrient Limitations
3.2. Other Stresses
| Growth Phase | Starvation | Other Stresses | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Strain | Exponential | Stationary | N | P | Ca2+ or Mg2+ | High Salinity | High Light | References | |
| c | Anabaena cylindrica | + | ++ | − | [44] | ||||
| c | Anabaena flos-aquae | + | ++ | [81,161] | |||||
| c | Anabaena sp. | + | + | − | [147] | ||||
| c | Anabaena torulosa | − | [49] | ||||||
| c | Anacystis nidulans | + | ++ | + | [111] | ||||
| c | Aphanocapsa halophyta | − | [112] | ||||||
| c | Aphanocapsa halophytica | + | ++ | [112] | |||||
| c | Arthrospira platensis | + | − | − | − | + | [29,40] | ||
| c | Chroococcus minutus | + | ++ | [50] | |||||
| c | Cyanospira capsulata | + | ++ | + | [162] | ||||
| c | Cyanothece | + | + | + | − | − | [115,148,149,169] | ||
| c | Microcoleus vaginatus | + | + | [31,54,55,175] | |||||
| c | Nostoc calcicola | + | + | [78] | |||||
| c | Nostoc insulare | + | ++ | [60] | |||||
| c | Nostoc sp. | + | − | + | + | [31,54,55,167,173] | |||
| c | Phormidium | − | − | + | + | [31,49,151,152] | |||
| c | Scytonema javanicum | + | [31] | ||||||
| c | Spirulina sp. | + | + | + | + | [49] | |||
| c | Synechococcus sp. | + | ++ | [128] | |||||
| c | Synechocystis | + | ++ | − | +/− | [52,176] | |||
| d | Achnanthes brevipes | − | + | [166] | |||||
| d | Chaetoceros affinis | + | − | + | [94] | ||||
| d | Cylindrotheca closterium | + | + | + | [150,165] | ||||
| d | Cylindrotheca fusiformis | + | + | [28] | |||||
| d | Nitzschia frustulum | + | [93] | ||||||
| d | Thalassiosira Pseudonana | + | + | [103,165] | |||||
| m | Botryococcus braunii | + | ++ | + | [140,170] | ||||
| m | Chlamydomonas | + | + | [141] | |||||
| m | Chlorella sp. | + | + | + | [142,164] | ||||
| m | Desmococcus olivaceus | + | [31] | ||||||
| m | Dunaliella salina | + | [174] | ||||||
| m | Flintiella sanguinaria | + | + | + | [59] | ||||
| m | Heterosigma akashiwo | + | ++ | [43] | |||||
| m | Porphyridium aeruginum | + | + | [143] | |||||
| m | Porphyridium marinum | + | + | + | + | [13] | |||
| m | Rhodella grisea | + | + | [144] | |||||
| m | Rhodella reticulata | + | + | [145] | |||||
| m | Rhodella violacea | + | + | + | [12] | ||||
| m | Scenedesmus | − | + | [153] | |||||
| m | Tetraselmis gracilis | + | + | [146] | |||||
3.3. Culture Strategies
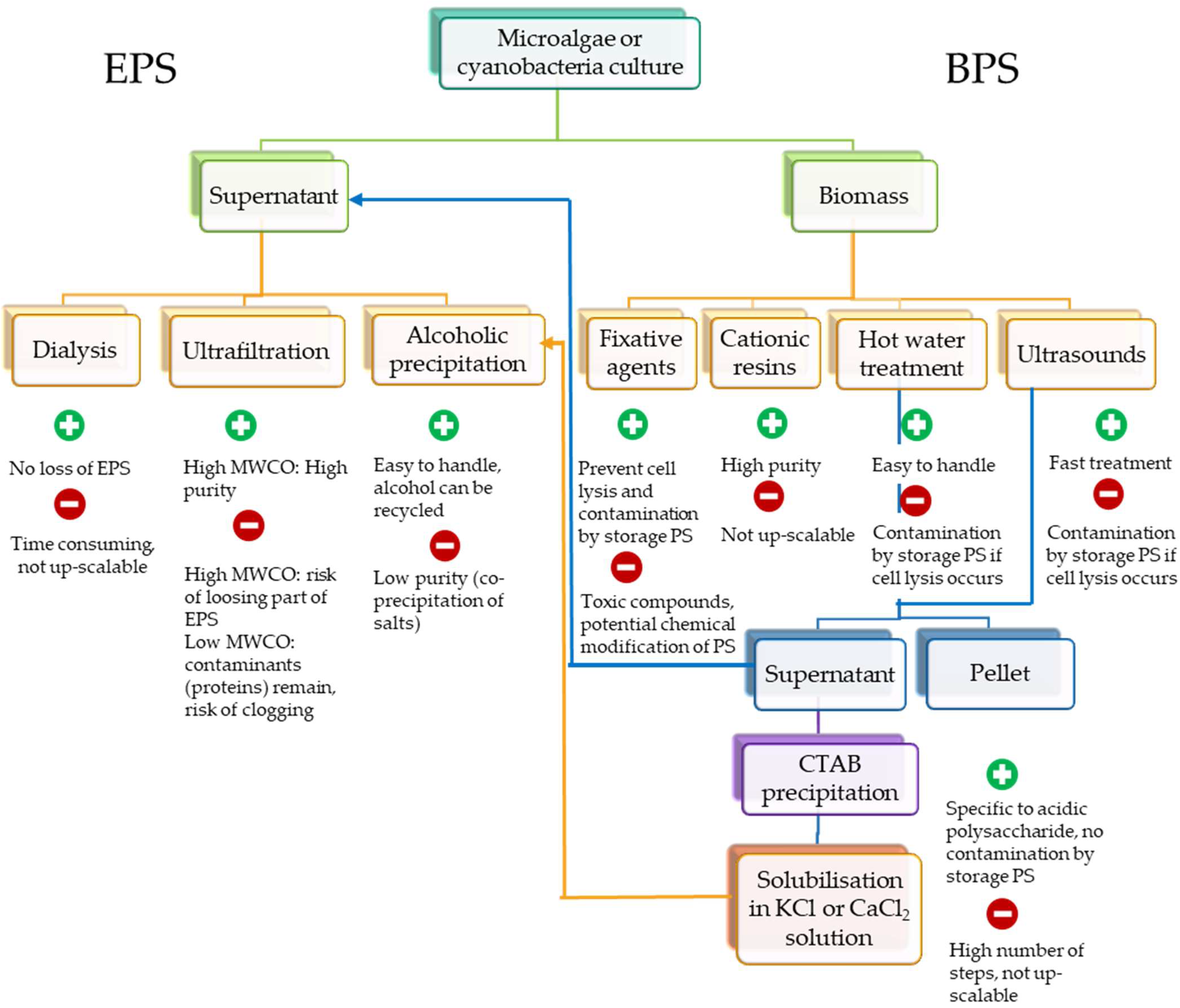
4. Processes for Extraction and Purification
4.1. Released EPSs
4.2. Cell-Bound PSs
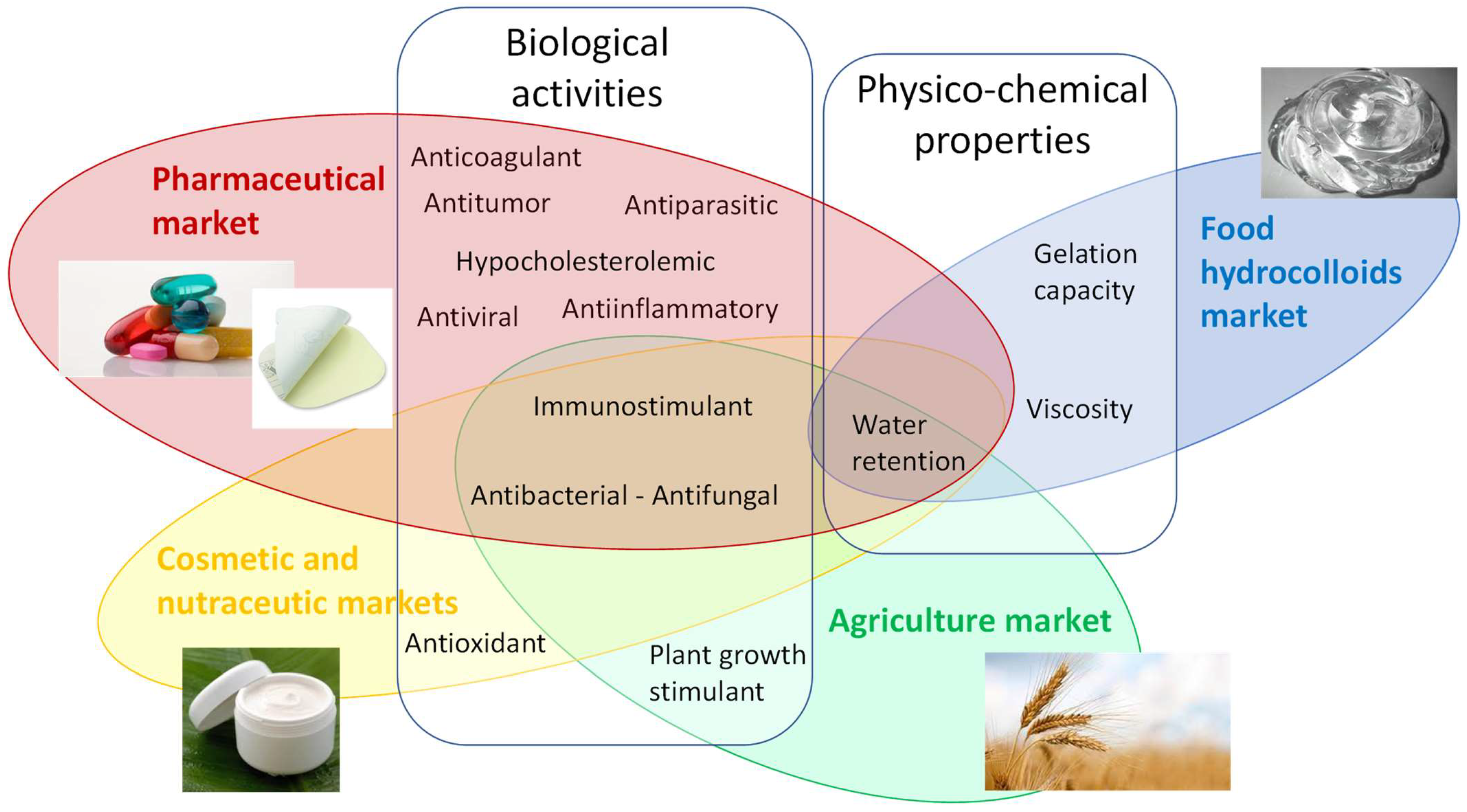
5. Applications
5.1. Biological Activities
5.2. Physico-Chemical Properties
5.3. Other Properties: Plant Stimulating Action
5.4. Potential Commercial Markets
5.4.1. Cosmetic and Nutraceutic Markets
5.4.2. Pharmaceutical and Medical Markets
5.4.3. Hydrocolloids
5.4.4. Agriculture Market
6. Conclusions
Funding
Conflicts of Interest
References
- Delattre, C.; Pierre, G.; Laroche, C.; Michaud, P. Production, extraction and characterization of microalgal and cyanobacterial exopolysaccharides. Biotechnol. Adv. 2016, 34, 1159–1179. [Google Scholar] [CrossRef]
- Dimitriu, S. (Ed.) Polysaccharides, Structural Diversity and Functional Versatility; Marcel Dekker: New York, NY, USA, 2004. [Google Scholar]
- Grima, E.M.; Belarbi, E.H.; Fernández, F.G.A.; Medina, A.R.; Chisti, Y. Recovery of microalgal biomass and metabolites: Process options and economics. Biotechnol. Adv. 2003, 20, 491–515. [Google Scholar] [CrossRef]
- Norsker, N.H.; Barbosa, M.J.; Vermuë, M.H.; Wijffels, R.H. Microalgal production—A close look at the economics. Biotechnol. Adv. 2011, 29, 24–27. [Google Scholar] [CrossRef]
- Pereira, S.; Zille, A.; Micheletti, E.; Moradas-Ferreira, P.; De Philippis, R.; Tamagnini, P. Complexity of cyanobacterial exopolysaccharides: Composition, structures, inducing factors and putative genes involved in their biosynthesis and assembly. FEMS Microbiol. Rev. 2009, 33, 917–941. [Google Scholar] [CrossRef]
- De Philippis, R.; Vincenzini, M. Exocellular polysaccharides from cyanobacteria and their possible applications. FEMS Microbiol. Rev. 1998, 22, 151–175. [Google Scholar] [CrossRef]
- Palmer, J.; Flint, S.; Brooks, J. Bacterial cell attachment, the beginning of a biofilm. J. Ind. Microbiol. Biotechnol. 2007, 34, 577–588. [Google Scholar] [CrossRef]
- Tamaru, Y.; Takani, Y.; Yoshida, T.; Sakamoto, T. Crucial role of extracellular polysaccharides in desiccation and freezing tolerance in the terrestrial cyanobacterium Nostoc. Commune. Appl. Env. Microbiol. 2005, 71, 7327–7333. [Google Scholar] [CrossRef] [Green Version]
- Shnyukova, E.I.; Zolotareva, E.K. Ecological role of exopolysaccharides of Bacillariophyta: A review. Algologia 2017, 27, 22–44. [Google Scholar] [CrossRef] [Green Version]
- Ramus, J. The capture and transduction of light energy. In The Biology of Seaweeds; Lobban, C.S., Wynne, M.J., Eds.; Blackwell Scientific Publications: Boston, MA, USA, 1981; pp. 458–492. [Google Scholar]
- Arad, S.M.; Levy-Ontman, O. Red microalgal cell-wall polysaccharides: Biotechnological aspects. Curr. Opin. Biotechnol. 2010, 21, 358–364. [Google Scholar] [CrossRef]
- Villay, A.; Laroche, C.; Roriz, D.; El Alaoui, H.; Delbac, F.; Michaud, P. Optimisation of culture parameters for exopolysaccharides production by the microalga Rhodella Violacea. Biores. Technol. 2013, 146, 732–735. [Google Scholar] [CrossRef]
- Soanen, N.; Da Silva, E.; Gardarin, C.; Michaud, P.; Laroche, C. Improvement of exopolysaccharide production by Porphyridium marinum. Biores. Technol. 2016, 213, 231–238. [Google Scholar] [CrossRef]
- Gaignard, C.; Gargouch, N.; Dubessay, P.; Delattre, C.; Pierre, G.; Laroche, C.; Fendri, I.; Abdelkafi, S.; Michaud, P. New horizons in culture and valorization of red microalgae. Biotechnol. Adv. 2019, 37, 193–222. [Google Scholar] [CrossRef]
- Belnap, J.; Lange, O.L. (Eds.) Biological Soil Crusts: Structure, Function and Management; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Chen, L.; Rossi, F.; Deng, S.; Liu, Y.; Wang, G.; Adessi, A.; De Philippis, R. Macromolecular and chemical features of the excreted extracellular polysaccharides in induced biological crusts of different ages. Soil Biol. Biochem. 2014, 78, 1–9. [Google Scholar] [CrossRef]
- Colica, G.; Li, H.; Rossi, F.; Li, D.; Liu, Y.; De Philippis, R. Microbial secreted exopolysaccharides affect the hydrological behavior of induced biological crusts in desert soils. Soil Biol. Biochem. 2014, 68, 62–72. [Google Scholar] [CrossRef]
- Mager, D.M.; Thomas, A.D. Extracellular polysaccharides from cyanobacterial soil crusts: A review of their role in dryland soil processes. J. Arid. Environ. 2011, 75, 91–97. [Google Scholar] [CrossRef]
- Underwood, G.J.C.; Paterson, D.M. The importance of extracellular carbohydrate production by marine epipelic diatoms. Adv. Bot. Res. 2003, 40, 184–240. [Google Scholar]
- Singh, S.; Arad, S.M.; Richmond, A. Extracellular polysaccharide production in outdoor mass cultures of Porphyridium sp. in flat plate glass reactors. J. Appl. Phycol. 2000, 12, 269–275. [Google Scholar] [CrossRef]
- Mugnai, B.; Rossi, F.; Felde, V.J.M.N.L.; Colesie, C.; Budel, B.; Peth, S.; Kaplan, A.; De Philippis, R. The potential of the cyanobacterium Leptolyngbya ohadii as inoculum for stabilizing bare sandy substrates. Soil Biol. Biochem. 2018, 127, 318–328. [Google Scholar] [CrossRef]
- Bernaerts, T.M.M.; Kyomugasho, C.; Van Looveren, N.; Gheysen, L.; Foubert, I.; Hendrickx, M.E.; Van Loey, A.M. Molecular and rheological characterization of different cell wall fractions of Porphyridium cruentum. Carbohydr. Polym. 2018, 195, 542–550. [Google Scholar] [CrossRef]
- Borowitzka, M.A. High-value products from microalgae—Their development and commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Tischer, R.G.; Moore, B.G. An extracellular polysaccharide produced by Palmella mucosa Kütz. Arch. Mikrobiol. 1964, 49, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Gaignard, C.; Laroche, C.; Pierre, G.; Dubessay, P.; Delattre, C.; Gardarin, C.; Gourvil, P.; Probert, I.; Dubuffet, A.; Michaud, P. Screening of microalgae: Investigation of new exopolysaccharide producers. Algal Res. 2019, 44, 101711. [Google Scholar] [CrossRef]
- Rossi, F.; De Philippis, R. Exocellular polysaccharides in microalgae and cyanobacteria: Chemical features, role and enzymes and genes involved in their biosynthesis. In Developments in Applied Phycology 6—The Physiology of Microalgae; Borowitzka, M.A., Beardall, J., Raven, J.A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 565–590. [Google Scholar]
- Cheng, Y.S.; Labavitch, J.M.; Vabdergheynst, J.S. Elevated CO2 concentration impacts cell wall polysaccharide composition of green microalgae of the genus Chlorella. Lett. Appl. Microbiol. 2015, 60, 1–7. [Google Scholar] [CrossRef]
- Magaletti, E.; Urbani, R.; Sist, P.; Ferrari, C.R.; Cicero, A.M. Abundance and chemical characterization of extracellular carbohydrates released by the marine diatom Cylindrotheca fusiformis under N- and P-limitation. Eur. J. Phycol. 2004, 39, 133–142. [Google Scholar] [CrossRef]
- Phelippé, M.; Goncalves, O.; Thouand, G.; Cogne, G.; Laroche, C. Characterization of the polysaccharides chemical diversity of the cyanobacteria Arthrospira platensis. Algal Res. 2019, 38, 101426. [Google Scholar] [CrossRef]
- Yim, J.H.; Kim, S.J.; Ahn, S.H.; Lee, H.K. Optimal conditions for the production of sulfated polysaccharide by marine microalga Gyrodinium impudicum strain KG03. Biomol. Eng. 2003, 20, 273–280. [Google Scholar] [CrossRef]
- Hu, C.; Liu, Y.; Paulsen, B.S.; Petersen, D.; Klaveness, D. Extracellular carbohydrate polymers from five desert soil algae with different cohesion in the stabilization of fine sand grain. Carbohydr. Polym. 2003, 54, 33–42. [Google Scholar] [CrossRef]
- Esqueda, A.B.; Gardarin, C.; Laroche, C. Exploring the diversity of red microalgae for exopolysaccharide production. Mar. Drugs 2022, 20, 246. [Google Scholar] [CrossRef]
- Kiemle, S.N.; Domozych, D.S.; Gretz, M.R. The extracellular polymeric substances of desmids (Conjugatophyceae, Streptophyta): Chemistry, structural analyses and implications in wetland biofilms. Phycologia 2007, 46, 617–627. [Google Scholar] [CrossRef]
- Barberousse, H.; Ruiz, G.; Gloaguen, V.; Lombardo, R.J.; Djediat, C.; Mascarell, G.; Castaing, J.C. Capsular polysaccharides secreted by building façade colonisers: Characterisation and adsorption to surfaces. Biofouling 2006, 22, 361–370. [Google Scholar] [CrossRef]
- Domozych, C.R.; Plante, K.; Blais, P.; Paliulis, L.; Domozych, D.S. Mucilage processing and secretion in the green alga Closterium. I. Cytology and Biochemistry. J. Phycol. 1993, 29, 650–659. [Google Scholar] [CrossRef]
- Giroldo, D.; Vieira, A.A.H.; Paulsen, B.S. Microbial degradation of extracellular polysaccharides released by a tropical strain of Staurastrum orbiculare (Zygnematophyceae). Phycologia 2005, 44, 671–677. [Google Scholar] [CrossRef]
- Jin, C.; Yu, Z.; Peng, S.; Feng, K.; Zhang, L.; Zhou, X. The characterization and comparison of exopolysaccharides from two benthic diatoms with different biofilm formation abilities. Acad. Bras. Ciênc. 2018, 90, 1503–1519. [Google Scholar] [CrossRef] [PubMed]
- Gouvêa, S.P.; Vieira, A.A.H.; Lombardi, A.T. Copper and cadmium complexation by high molecular weight materials of dominant microalgae and of water from a eutrophic reservoir. Chemosphere 2005, 60, 1332–1339. [Google Scholar] [CrossRef]
- Li, P.; Liu, Z.; Xu, R. Chemical characterisation of the released polysaccharide from the cyanobacterium Aphanothece halophytica GR02. J. Appl. Phycol. 2001, 13, 71–77. [Google Scholar] [CrossRef]
- Filali-Mouhim, R.; Cornet, J.-F.; Fontane, T.; Fournet, B.; Dubertret, G. Production, isolation and preliminary characterization of the exopolysaccharide of the cyanobacterium Spirulina platensis. Biotechnol. Lett. 1993, 15, 567–572. [Google Scholar] [CrossRef]
- Vincenzini, M.; De Philippis, R.; Sili, C.; Materassi, R. Stability of molecular and rheological properties of the exopolysaccharide produced by Cyanospira capsulata cultivated under different growth conditions. J. Appl. Phycol. 1993, 5, 539–541. [Google Scholar] [CrossRef]
- Bellezza, S.; Albertano, P.; De Philippis, R.; Paradossi, G. Exopolysaccharides of two cyanobacterial strains from Roman hypogea. Geomicrobiol. J. 2006, 23, 301–310. [Google Scholar] [CrossRef]
- Lopes, D.C.; Maurer, J.B.B.; Stevan-Hancke, F.R.; de Oliveira Proença, L.A.; Zawadzki-Baggio, S.F. Chemical analysis of exopolysaccharide fractions and lipid compounds of the microalga Heterosigma akashiwo grown in vitro. Bot. Mar. 2012, 55, 565–580. [Google Scholar] [CrossRef]
- Lama, L.; Nicolaus, B.; Calandrelli, V.; Manca, M.C.; Romano, I.; Gambacorta, A. Effect of growth conditions on endo- and exopolymer biosynthesis in Anabaena cylindrica 10C. Phytochem 1996, 42, 655–659. [Google Scholar] [CrossRef]
- Richert, L.; Golubic, S.; Guédès, R.L.; Ratiskol, J.; Payri, C.; Guezennec, J. Characterization of exopolysaccharides produced by cyanobacteria isolated from Polynesian microbial mats. Curr. Microbiol. 2005, 51, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Di Pippo, F.; Ellwood, N.T.W.; Gismondi, A.; Bruno, L.; Rossi, F.; Magni, P.; De Philippis, R. Characterization of exopolysaccharides produced by seven biofilm-forming cyanobacterial strains for biotechnological applications. J. Appl. Phycol. 2013, 25, 1697–1708. [Google Scholar] [CrossRef] [Green Version]
- Tesson, B.; Hildebrand, M. Characterization and location of insoluble organic matrices associated with diatom cell walls: Insight into their roles during cell wall formation. PLoS ONE 2013, 8, e61675. [Google Scholar] [CrossRef] [Green Version]
- Giroldo, D.; Vieira, A.A.H.; Paulsen, B.S. Relative increase of deoxy sugars during microbial degradation of an extracellular polysaccharide released by a tropical freshwater Thalassiosira sp. (bacillariophyceae). J. Phycol. 2003, 39, 1109–1115. [Google Scholar] [CrossRef]
- Nicolaus, B.; Panico, A.; Lama, L.; Romano, I.; Manca, M.C.; De Giulio, A.; Gambacorta, A. Chemical composition and production of exopolysaccharides from representative members of heterocystous and non-heterocystous cyanobacteria. Phytochem 1999, 52, 639–647. [Google Scholar] [CrossRef]
- Fischer, D.; Schlösser, U.G.; Pohl, P. Exopolysaccharide production by cyanobacteria grown in closed photobioreactors and immobilized using white cotton towelling. J. Appl. Phycol. 1997, 9, 205–213. [Google Scholar] [CrossRef]
- Rossi, F.; Micheletti, E.; Bruno, L.; Adhikary, S.P.; Albertano, P.; De Philippis, R. Characteristics and role of the exocellular polysaccharides produced by five cyanobacteria isolated from phototrophic biofilms growing on stone monuments. Biofouling 2012, 28, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Panoff, J.M.; Priem, B.; Morvan, H.; Joset, F. Sulphated exopolysaccharides produced by two unicellular strains of Cyanobacteria, Synechocystis PCC 6803 and 6714. Arch. Microbiol. 1988, 150, 558–563. [Google Scholar] [CrossRef]
- Majdoub, H.; Mansour, M.B.; Chaubet, F.; Roudesli, M.S.; Maaroufi, R.M. Anticoagulant activity of a sulfated polysaccharide from the green alga Arthrospira platensis. Biochim. Biophys. Acta Gen. Subj. 2009, 1790, 1377–1381. [Google Scholar] [CrossRef]
- Ge, H.; Xia, L.; Zhou, X.; Zhang, D.; Hu, C. Effects of light intensity on components and topographical structures of extracellular polysaccharides from the cyanobacteria Nostoc sp. J. Microbiol. 2014, 52, 179–183. [Google Scholar] [CrossRef]
- Ge, H.; Zhang, J.; Zhou, X.; Xia, L.; Hu, C. Effects of light intensity on components and topographical structures of extracellular polymeric substances from Microcoleus vaginatus (Cyanophyceae). Phycologia 2014, 53, 167–173. [Google Scholar] [CrossRef]
- Tease, B.; Jürgens, U.J.; Golecki, J.R.; Heinrich, U.R.; Rippka, R.; Weckesser, J. Fine-structural and chemical analyses on inner and outer sheath of the cyanobacterium Gloeothece sp. PCC 6909. Antonie Leeuwenhoek 1991, 59, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xu, C.; Santschi, P.H. Chemical composition and 234Th (IV) binding of extracellular polymeric substances (EPS) produced by the marine diatom Amphora sp. Mar. Chem. 2008, 112, 81–92. [Google Scholar] [CrossRef]
- Percival, E.; Foyle, R.A.J. The extracellular polysaccharides of Porphyridium cruentum and Porphyridium aerugineum. Carbohydr. Res. 1979, 72, 165–176. [Google Scholar] [CrossRef]
- Gaignard, C.; Macao, V.; Gardarin, C.; Rihouey, C.; Picton, L.; Michaud, P.; Laroche, C. The red microalga Flintiella sanguinaria as a new exopolysaccharide producer. J. Appl. Phycol. 2018, 30, 2803–2814. [Google Scholar] [CrossRef]
- Volk, R.B.; Venzke, K.; Blaschek, W. Structural investigation of a polysaccharide released by the cyanobacterium Nostoc insulare. J. Appl. Phycol. 2006, 19, 255–262. [Google Scholar] [CrossRef]
- Capek, P.; Matulová, M.; Combourieu, B. The extracellular proteoglycan produced by Rhodella grisea. Int. J. Biol. Macromol. 2008, 43, 390–393. [Google Scholar] [CrossRef] [Green Version]
- Geresh, S.; Lupescu, N.; Arad, S.M. Fractionation and partial characterization of the sulphated polysaccharide of Porphyridium. Phytochemistry 1992, 31, 4181–4186. [Google Scholar] [CrossRef]
- Levy-Ontman, O.; Arad, S.M.; Harvey, D.J.; Parsons, T.B.; Fairbanks, A.; Tekoah, Y. Unique N-glycan moieties of the 66-kDa cell wall glycoprotein from the red microalga Porphyridium sp. J. Biol. Chem. 2011, 286, 21340–21352. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-B.; Hayashi, T.; Hayashi, K.; Sankawa, U.; Maeda, M.; Nemoto, T.; Nakanishi, H. Further purification and structural analysis of Calcium spirulan from Spirulina platensis. J. Nat. Prod. 1998, 61, 1101–1104. [Google Scholar] [CrossRef]
- Bafana, A. Characterization and optimization of production of exopolysaccharide from Chlamydomonas reinhardtii. Carbohydr. Polym. 2013, 95, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Khandeparker, R.D.; Bhosle, N.B. Extracellular polymeric substances of the marine fouling diatom Amphora rostrata Wm. Sm. Biofouling 2001, 17, 117–127. [Google Scholar] [CrossRef]
- Bhosle, N.B.; Sawant, S.S.; Garg, A.; Wagh, A.B. Isolation and partial chemical analysis of exopolysaccharides from the marine fouling diatom Navicula subinflata. Bot. Mar. 1995, 38, 103–110. [Google Scholar] [CrossRef]
- Vieira, A.A.H.; Paulsen, B.S. Studies on the composition of extracellular polysaccharide produced by Hyalotheca dissiliens (Desmidiaceae). Biol. Bratis. 1994, 49, 535–539. [Google Scholar]
- Oertel, A.; Aichinger, N.; Hochreiter, R.; Thalhamer, J.; Lütz-Meindl, U. Analysis of mucilage secretion and excretion in Micrasterias (Chlorophyta) by means of immunoelectron microscopy and digital time lapse video microscopy. J. Phycol. 2004, 40, 711–720. [Google Scholar] [CrossRef]
- Domozych, D.S.; Kort, S.; Benton, S.; Yu, T. The extracellular polymeric substance of the green alga Penium margaritaceum and its role in biofilm formation. Biofilms 2005, 2, 129–144. [Google Scholar] [CrossRef]
- Paulsen, B.S.; Vieira, A.A.H. Structure of the capsular and extracellular polysaccharides produced by the desmid Spondylosium panduriforme (Chlorophyta). J. Phycol. 1994, 30, 638–641. [Google Scholar] [CrossRef]
- Freire-Nordi, C.S.; Vieira, A.A.H.; Nakaie, C.R.; Nascimento, O.R. Effect of polysaccharide capsule of the microalgae Staurastrum iversenii var. americanum on diffusion of charged and uncharged molecules, using EPR technique. Braz. J. Phys. 2006, 36, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, B.S.; Aslaksen, T.; Freire-Nordi, C.S.; Vieira, A.A.H. Extracellular polysaccharides from Ankistrodesmus Densus (Chlorophyceae). J. Phycol. 1998, 34, 638–641. [Google Scholar] [CrossRef]
- Allard, B.; Casadevall, E. Carbohydrate composition and characterization of sugars from the green microalga Botryococcus braunii. Phytochemistry 1990, 29, 1875–1878. [Google Scholar] [CrossRef]
- Casadevall, E.; Dif, D.; Largeau, C.; Gudin, C.; Chaumont, D.; Desanti, O. Studies on batch and continuous cultures of Botryococcus braunii: Hydrocarbon production in relation to physiological state, cell ultrastructure, and phosphate nutrition. Biotech. Bioeng. 1985, 27, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Sharma, R.; Chisti, Y.; Banerjee, U.C. Botryococcus braunii: A renewable source of hydrocarbons and other chemicals. Crit. Rev. Biotechnol. 2002, 22, 245–279. [Google Scholar] [CrossRef] [PubMed]
- Allard, B.; Tazi, A. Influence of growth status on composition of extracellular polysaccharides from two Chlamydomonas species. Phytochemistry 1992, 32, 41–47. [Google Scholar] [CrossRef]
- Flaibani, A.; Olsen, Y.; Painter, T.J. Polysaccharides in desert reclamation: Compositions of exocellular proteoglycan complexes produced by filamentous blue-green and unicellular green edaphic algae. Carbohydr. Res. 1989, 190, 235–248. [Google Scholar] [CrossRef]
- Barclay, W.R.; Lewin, R.A. Microalgal polysaccharide production for the conditioning of agricultural soils. Plant Soil 1985, 88, 159–169. [Google Scholar] [CrossRef]
- Kaplan, D.; Christiaen, D.; Arad, S.M. Chelating properties of extracellular polysaccharides from Chlorella spp. Appl. Environ. Microbiol. 1987, 53, 2953–2956. [Google Scholar] [CrossRef] [Green Version]
- Moore, B.G.; Tischer, R.G. Extracellular polysaccharides of algae: Effects on life-support systems. Science 1964, 145, 586–587. [Google Scholar] [CrossRef]
- Fábregas, J.; García, D.; Fernandez-Alonso, M.; Rocha, A.I.; Gómez-Puertas, P.; Escribano, J.M.; Otero, A.; Coll, J.M. In vitro inhibition of the replication of haemorrhagic septicaemia virus (VHSV) and African swine fever virus (ASFV) by extracts from marine microalgae. Antivir. Res. 1999, 44, 67–73. [Google Scholar] [CrossRef]
- Maksimova, I.V.; Bratkovskaya, L.B.; Plekhanov, S.E. Extracellular carbohydrates and polysaccharides of the alga Chlorella pyrenoidosa Chick S-39. Biol. Bulletin 2004, 31, 175–181. [Google Scholar] [CrossRef]
- Harun, R.; Danquah, M.K. Influence of acid pre-treatment on microalgal biomass for bioethanol production. Process. Biochem. 2011, 46, 304–309. [Google Scholar] [CrossRef]
- Brown, M.R. The amino acid and sugar composition of 16 species of microalgae used in mariculture. J. Exp. Mar. Biol. Ecol. 1991, 145, 79–99. [Google Scholar] [CrossRef]
- You, T.; Barnett, S.M. Effect of light quality on production of extracellular polysaccharides and growth rate of Porphyridium cruentum. Biochem. Eng. J. 2004, 19, 251–258. [Google Scholar] [CrossRef]
- Park, J.K.; Kim, Z.-H.; Lee, C.G.; Synytsya, A.; Jo, H.S.; Kim, S.O.; Park, J.W.; Park, Y.I. Characterization and Immunostimulating Activity of a Water-soluble Polysaccharide Isolated from Haematococcus lacustris. Biotechnol. Bioprocess Eng. 2011, 16, 1090–1098. [Google Scholar] [CrossRef]
- Mohamed, Z.A. Polysaccharides as a protective response against microcystin-induced oxidative stress in Chlorella vulgaris and Scenedesmus quadricauda and their possible significance in the aquatic ecosystem. Ecotoxicology 2008, 17, 504–516. [Google Scholar] [CrossRef]
- Hasui, M.; Matsuda, M.; Okutani, K.; Shigeta, S. In vitro antiviral activities of sulfated polysaccharides from a marine microalga (Cochlodinium polykrikoides) against human immunodeficiency virus and other enveloped viruses. Int. J. Biol. Macromol. 1995, 17, 293–297. [Google Scholar] [CrossRef]
- Liu, B.; Sun, Z.; Ma, X.; Yang, B.; Jiang, Y.; Wei, D.; Chen, F. Mutation breeding of extracellular polysaccharide-producing microalga Crypthecodinium cohnii by a novel mutagenesis with atmospheric and room temperature plasma. Int. J. Mol. Sci. 2015, 16, 8201–8212. [Google Scholar] [CrossRef] [Green Version]
- Wustman, B.A.; Gretz, M.R.; Hoagland, K.D. Extracellular matrix assembly in diatoms (Bacillariophyceae) (I. A model of adhesives based on chemical characterization and localization of polysaccharides from the marine diatom Achnanthes longipes and other diatoms). Plant Physiol. 1997, 113, 1059–1069. [Google Scholar] [CrossRef] [Green Version]
- Leandro, S.M.; Gil, M.C.; Delgadillo, I. Partial characterisation of exopolysaccharides exudated by planktonic diatoms maintained in batch cultures. Acta Oecologica 2003, 24, S49–S55. [Google Scholar] [CrossRef]
- Allan, G.G.; Lewin, J.; Johnson, P.G. Marine polymers IV Diatom polysaccharides. Bot. Mar. 1972, 15, 102–108. [Google Scholar] [CrossRef]
- Myklestad, S.; Haug, A.; Larsen, B. Production of carbohydrates by the marine diatom Chaetoceros affinis var. willei (Gran) Hustedt II. Preliminary investigation of the extracellular polysaccharide. J. Exp. Mar. Biol. Ecol. 1972, 9, 137–144. [Google Scholar]
- Paulsen, B.S.; Haug, A.; Myklestad, S.M. Structural studies of the extracellular polysaccharide produced by the diatom Chaetoceros curvisetus Cleve. Acta Chem. Scand Ser. B Org. Chem. Biochem. 1975, 29, 337–340. [Google Scholar]
- Urbani, R.; Sist, P.; Pletikapić, G.; Radić, T.M.; Svetličić, V.; Žutic, V. Diatom polysaccharides: Extracellular production, isolation and molecular characterization. In The Complex World of Polysaccharides; Karunaratne, D.N., Ed.; IntechOpen: London, UK, 2012; pp. 345–370. [Google Scholar]
- Haug, A.; Myklestad, S. Polysaccharides of marine diatoms with special reference to Chaetoceros species. Mar. Biol. 1976, 34, 217–222. [Google Scholar] [CrossRef]
- Percival, E.; Rahman, M.A.; Weigel, H. Chemistry of the polysaccharides of the diatom Coscinodiscus nobilis. Phytochemistry 1980, 19, 809–811. [Google Scholar] [CrossRef]
- Staats, N.; De Winder, B.; Stal, L.; Mur, L. Isolation and characterization of extracellular polysaccharides from the epipelic diatoms Cylindrotheca closterium and Navicula salinarum. Eur. J. Phycol. 1999, 34, 161–169. [Google Scholar] [CrossRef]
- Bellinger, B.J.; Abdullahi, A.S.; Gretz, M.R.; Underwood, G.J.C. Biofilm polymers: Relationship between carbohydrate polymers from estuarine mudflats and unialgal cultures of benthic diatoms. Aquat. Microb. Ecol. 2005, 38, 169–180. [Google Scholar] [CrossRef]
- Willis, A.; Chiovitti, A.; Dugdale, T.M.; Wetherbee, R. Characterization of the extracellular matrix of Phaeodactylum tricornutum (Bacillariophyceae): Structure, composition, and adhesive characteristics. J. Phycol. 2013, 49, 937–949. [Google Scholar] [CrossRef]
- Chiovitti, A.; Higgins, M.J.; Harper, R.E.; Wetherbee, R.; Bacic, A. The complex polysaccharides of the raphid diatom Pinnularia viridis (Bacillariophyceae). J. Phycol. 2003, 39, 543–554. [Google Scholar] [CrossRef]
- Ai, X.X.; Liang, J.R.; Gao, Y.H.; Lo, S.C.L.; Lee, F.W.F.; Chen, C.P.; Luo, C.S.; Du, C. MALDI-TOF MS analysis of the extracellular polysaccharides released by the diatom Thalassiosira pseudonana under various nutrient conditions. J. Appl. Phycol. 2015, 27, 673–684. [Google Scholar] [CrossRef]
- Chen, X.; Song, L.; Wang, H.; Liu, S.; Yu, H.; Wang, X.; Li, R.; Liu, T.; Li, P. Partial characterization, the immune modulation and anticancer activities of sulfated polysaccharides from filamentous microalgae Tribonema sp. Molecules 2019, 24, 322. [Google Scholar] [CrossRef] [Green Version]
- De Jesus Raposo, M.F.; de Morais, A.M.M.B.; de Morais, R.M.S.C. Influence of sulphate on the composition and antibacterial and antiviral properties of the exopolysaccharide from Porphyridium cruentum. Life Sci. 2014, 101, 56–63. [Google Scholar] [CrossRef]
- Roussel, M.; Villay, A.; Delbac, F.; Michaud, P.; Laroche, C.; Roriz, D.; El Alaoui, H.; Diogon, M. Antimicrosporidian activity of sulphated polysaccharides from algae and their potential to control honeybee nosemosis. Carbohydr. Polym. 2015, 133, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Medina-Cabrera, E.; Rühmann, B.; Schmid, J.; Sieber, V. Characterization and comparison of Porphyridium sordidum and Porphyridium purpureum concerning growth characterisitics and polysaccharide production. Algal Res. 2020, 49, 101931. [Google Scholar] [CrossRef]
- Geresh, S.; Arad, S.M.; Levy-Ontman, O.; Zhang, W.; Tekoah, Y.; Glaser, R. Isolation and characterization of poly- and oligosaccharides from the red microalga Porphyridium sp. Carbohydr. Res. 2009, 344, 343–349. [Google Scholar] [CrossRef]
- Cohen, A.K.; Arad, S.M. Biosynthesis of the cell wall polysaccharide in the red microalga Rhodella reticulata. Isr. J. Plant Sci. 1998, 46, 147–153. [Google Scholar] [CrossRef]
- Moreno, J.; Vargas, M.A.; Madiedo, J.M.; Muñoz, J.; Rivas, J.; Guerrero, M.G. Chemical and rheological properties of an extracellular polysaccharide produced by the cyanobacterium Anabaena sp. ATCC 33047. Biotechnol. Bioeng. 2000, 67, 283–290. [Google Scholar] [CrossRef]
- Sangar, V.K.; Dugan, P.R. Polysaccharide produced by Anacystis nidulans: Its ecological implication. Appl. Microbiol. 1972, 24, 732–734. [Google Scholar] [CrossRef]
- Sudo, H.; Burgess, J.G.; Takemasa, H.; Nakamura, N.; Matsunaga, T. Sulfated exopolysaccharide production by the halophilic cyanobacterium Aphanocapsa halophytia. Curr. Microbiol. 1995, 30, 219–222. [Google Scholar] [CrossRef]
- Nie, Z.Y.; Xia, J.L.; Levert, J.M. Fractionation and characterization of polysaccharides from cyanobacterium Spirulina (Arthrospira) maxima in nitrogen-limited batch culture. J. Cent. South Univ. Technol. 2002, 9, 81–86. [Google Scholar] [CrossRef]
- Trabelsi, L.; M’sakni, N.H.; Ouada, H.B.; Bacha, H.; Roudesli, S. Partial characterization of extracellular polysaccharides produced by cyanobacterium Arthrospira platensis. Biotechnol. Bioprocess. Eng. 2009, 14, 27–31. [Google Scholar] [CrossRef]
- De Philippis, R.; Margheri, M.C.; Pelosi, E.; Ventura, S. Exopolysaccharide production by a unicellular cyanobacterium isolated from a hypersaline habitat. J. Appl. Phycol. 1993, 5, 387–394. [Google Scholar] [CrossRef]
- Parikh, A.; Madamwar, D. Partial characterization of extracellular polysaccharides from Cyanobacteria. Bioresour. Technol. 2006, 97, 1822–1827. [Google Scholar] [CrossRef] [PubMed]
- Raungsomboon, S.; Chidthaisong, A.; Bunnag, B.; Inthorn, D.; Harvey, N.W. Production, composition and Pb2+ adsorption characteristics of capsular polysaccharides extracted from a cyanobacterium Gloeocapsa gelatinosa. Water Res. 2006, 40, 3759–3766. [Google Scholar] [CrossRef] [PubMed]
- Najdenski, H.M.; Gigova, L.G.; Iliev, I.I.; Pilarski, P.S.; Lukavský, J.; Tsvetkova, I.V.; Ninova, M.S.; Kussovski, V.K. Antibacterial and antifungal activities of selected microalgae and Cyanobacteria. Int. J. Food Sci. Technol. 2013, 48, 1533–1540. [Google Scholar] [CrossRef]
- Micheletti, E.; Pereira, S.; Mannelli, F.; Moradas-Ferreira, P.; Tamagnini, P.; De Philippis, R. Sheathless mutant of cyanobacterium Gloeothece sp. Strain PCC 6909 with increased capacity to remove copper ions from aqueous solutions. Appl. Environ. Microbiol. 2008, 74, 2797–2804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gloaguen, V.; Morvan, H.; Hoffmann, L. Released and capsular polysaccharides of Oscillatoriaceae (Cyanophyceae, Cyanobacteria). Algol. Stud. 1995, 78, 53–69. [Google Scholar] [CrossRef]
- Gloaguen, V.; Morvan, H.; Hoffmann, L.; Plancke, Y.; Wieruszeski, J.-M.; Lippens, G.; Strecker, G. Capsular polysaccharide produced by the thermophilic cyanobacterium Mastigocladus laminosus; Structural study of an undecasaccharide obtained by lithium degradation. Eur. J. Biochem. 1999, 266, 762–770. [Google Scholar] [CrossRef]
- Forni, C.; Telo, F.R.; Caiola, M.G. Comparative analysis of the polysaccharides produced by different species of Microcystis (Chroococcales, Cyanophyta). Phycologia 1997, 36, 181–185. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Y.; Paulsen, B.S.; Klaveness, D. Studies on polysaccharides from three edible species of Nostoc (Cyanobacteria) with different colony morphologies: Comparison of monosaccharide compositions and viscosities of polysaccharides from field colonies and suspension cultures. J. Phycol. 1998, 34, 962–968. [Google Scholar] [CrossRef]
- Helm, R.F.; Huang, Z.; Edwards, D.; Leeson, H.; Peery, W.; Potts, M. Structural characterization of the released polysaccharide of desiccation-tolerant Nostoc commune DRH-1. J. Bacteriol. 2000, 182, 974–982. [Google Scholar] [CrossRef] [Green Version]
- Hokputsa, S.; Hu, C.; Paulsen, B.S.; Harding, S.E. A physico-chemical comparative study on extracellular carbohydrate polymers from five desert algae. Carbohydr. Polym. 2003, 54, 27–32. [Google Scholar] [CrossRef]
- Sakamoto, T.; Kumihashi, K.; Kunita, S.; Masaura, T.; Inoue-Sakamoto, K.; Yamaguchi, M. The extracellular-matrix-retaining cyanobacterium Nostoc verrucosum accumulates trehalose but is sensitive to desiccation. FEMS Microbiol. Ecol. 2011, 77, 385–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silipo, A.; Molinaro, A.; Molteni, M.; Rossetti, C.; Parrilli, M.; Lanzetta, R. Full Structural Characterization of an Extracellular Polysaccharide Produced by the Freshwater Cyanobacterium Oscillatoria planktothrix FP1. Eur. J. Org. Chem. 2010, 29, 5594–5600. [Google Scholar] [CrossRef]
- Phlips, E.J.; Zeman, C.; Hansen, P. Growth, photosynthesis, nitrogen fixation and carbohydrate production by a unicellular cyanobacterium Synechococcus sp. (Cyanophyta). J. Appl. Phycol. 1989, 1, 137–145. [Google Scholar] [CrossRef]
- Geresh, S.; Dubinsky, O.; Arad, S.; Christiaen, D. Structure of a 3-O-(α-Dglucopyranosyluronic acid)-L-galactopyranose, an aldobiouronic acid isolated from the polysaccharides of various unicellular red algae. Carbohydr. Res. 1990, 208, 301–305. [Google Scholar] [CrossRef]
- Gloaguen, V.; Ruiz, G.; Morvan, H.; Mouradi-Givernaud, A.; Maes, E.; Krausz, P.; Strecker, G. The extracellular polysaccharide of Porphyridium sp.: An NMR study of lithium-resistant oligosaccharidic fragments. Carbohydr. Res. 2004, 339, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Shah, V.; Ray, A.; Garg, N.; Madamwar, D. Characterization of the extracellular polysaccharide produced by a marine cyanobacterium, Cyanothece sp. ATCC 51142, and its exploitation toward metal removal from solutions. Curr. Microbiol. 2000, 40, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Garozzo, D.; Impallomeni, G.; Spina, E.; Sturiale, L. The structure of the exocellular polysaccharide from the cyanobacterium Cyanospira capsulata. Carbohydr. Res. 1998, 307, 113–124. [Google Scholar] [CrossRef]
- Pereira, S.; Mota, R.; Santos, C.L.; De Philippis, R.; Tamagnini, P. Assembly and Export of Extracellular Polymeric Substances (EPS) in Cyanobacteria: A Phylogenomic Approach. Adv. Bot. Res. 2013, 65, 235–279. [Google Scholar]
- Li, S.-Y.; Lellouche, J.-P.; Shabtai, Y.; Arad, S.M. Fixed carbon partitioning in the red microalga Porphyridium sp. (rhodophyta). J. Phycol. 2001, 37, 289–297. [Google Scholar] [CrossRef]
- Li, S.-Y.; Shabtai, Y.; Arad, S.M. Floridoside as a carbon precursor for the synthesis of cell-wall polysaccharides in the red microalga Porphyridium sp. (rhodophyta). J. Phycol. 2002, 38, 931–938. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Monsigny, M.; Petit, C.; Roche, A.C. Colorimetric determination of neutral sugars by a resorcinol sulfuric acid micromethod. Anal. Biochem. 1988, 175, 525–530. [Google Scholar] [CrossRef]
- Nichols, C.A.M.; Nairn, K.M.; Glattauer, V.; Blackburn, S.I.; Ramshaw, J.A.M.; Graham, L.D. Screening Microalgal Cultures in Search of Microbial Exopolysaccharides with Potential as Adhesives. J. Adhes. 2009, 85, 97–125. [Google Scholar] [CrossRef]
- Badel, S.; Callet, F.; Laroche, C.; Gardarin, C.; Petit, E.; El Alaoui, H.; Bernardi, T.; Michaud, P. A new tool to detect high viscous exopolymers from microalgae. J. Ind. Microbiol. Biotechnol. 2011, 38, 319–326. [Google Scholar] [CrossRef]
- Lupi, F.M.; Fernandes, H.M.L.; Tomé, M.M.; Sá-Correia, I.; Novais, J.M. Influence of nitrogen source and photoperiod on exopolysaccharide synthesis by the microalga Botryococcus braunii UC 58. Enzym. Microb. Technol. 1994, 16, 546–550. [Google Scholar] [CrossRef]
- Kroen, W.K.; Rayburn, W.R. Influence of growth status and nutrients on extracellular polysaccharide synthesis by the soil alga Chlamydomonas mexicana (Chlorophyceae). J. Phycol. 1984, 20, 253–257. [Google Scholar] [CrossRef]
- Brányiková, I.; Marsalková, B.; Doucha, J.; Brányik, T.; Bisová, K.; Zachleder, V. Microalgae-novel highly efficient starch producers. Biotechnol. Bioeng. 2011, 108, 766–776. [Google Scholar] [CrossRef]
- Ramus, J. The production of extracellular polysaccharide by the unicellular red alga Porphyridium aeruginum. J. Phycol. 1972, 8, 97–111. [Google Scholar]
- Arad, S.M.; Lerental, Y.B.; Dubinsky, O. Effect of nitrate and sulfate starvation on polysaccharide formation in Rhodella reticulata. Biores. Technol. 1992, 42, 141–148. [Google Scholar] [CrossRef]
- Dubinsky, O.; Lerental, Y.B.; Christiaen, D.; Glaser, R.; Barak, Z.; Arad, S.M. Production and characterization of polysaccharides in the unicellular red alga Rhodella reticulata. In Algal Biotechnology; Stadler, T., Mollion, J., Verdus, M.C., Karamanos, Y., Morvan, H., Christiaen, D., Eds.; Elsevier: London, UK, 1988; pp. 451–461. [Google Scholar]
- Lourenço, S.O.; Marquez, U.M.L.; Mancini-Filho, J.; Barbarino, E.; Aidar, E. Changes in biochemical profile of Tetraselmis gracilis I. Comparison of two culture media. Aquaculture 1997, 148, 153–168. [Google Scholar] [CrossRef]
- Moreno, J.; Vargas, M.A.; Olivares, H.; Rivas, J.; Guerrero, M.G. Exopolysaccharide production by the cyanobacterium Anabaena sp. ATCC 33047 in batch and continuous culture. J. Biotechnol. 1998, 60, 175–182. [Google Scholar] [CrossRef]
- De Philippis, R.; Margheri, M.C.; Materassi, R.; Vincenzini, M. Potential of unicellular cyanobacteria from saline environments as exopolysaccharide producers. Appl. Environ. Microbiol. 1998, 64, 1130–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, K.J.; Soper, B.W.; Tang, J.; Bradley, R.L. Phenotypic variation in exopolysaccharide production in the marine, aerobic nitrogen-fixing unicellular cyanobacterium Cyanothece sp. World J. Microbiol. Biotechnol. 1996, 12, 311–318. [Google Scholar] [CrossRef]
- Underwood, G.J.C.; Boulcott, M.; Raines, C.A.; Waldron, K. Environmental effects on exopolymer production by marine benthic diatoms: Dynamics, changes in composition, and pathways of production. J. Phycol. 2004, 40, 293–304. [Google Scholar] [CrossRef]
- Fattom, A.; Shilo, M. Phormidium J-1 bioflocculant: Production and activity. Arch. Microbiol. 1984, 139, 421–426. [Google Scholar] [CrossRef]
- Fattom, A.; Shilo, M. Production of emulcyan by Phormidium J-1: Its activity and function. FEMS Microbiol. Ecol. 1985, 31, 3–9. [Google Scholar] [CrossRef]
- Angelaalincy, M.J.; Senthilkumar, N.; Karpagam, R.; Kumar, G.G.; Ashokkumar, B.; Varalakshmi, P. Enhanced Extracellular Polysaccharide Production and Self Sustainable Electricity Generation for PAMFCs by Scenedesmus sp. SB1. ACS Omega 2017, 2, 3754–3765. [Google Scholar] [CrossRef] [Green Version]
- Arad, S.M. Production of sulphated polysaccharides from red unicellular algae. In Algal Biotechnology; Stadler, T., Mollion, J., Verdus, M.C., Karamanos, Y., Morvan, H., Christiaen, D., Eds.; Cambridge University Press: Cambridge, UK, 1988; pp. 65–87. [Google Scholar]
- Kumar, A.S.; Mody, K.; Jha, B. Bacterial exopolysaccharides—A perception. J. Basic Microbiol. 2007, 47, 103–117. [Google Scholar] [CrossRef]
- Otero, A.; Vincenzini, M. Nostoc (cyanophyceae) goes nude: Extracellular polysaccharides serve as a sink for reducing power under unbalanced C/N metabolism. J. Phycol. 2004, 40, 74–81. [Google Scholar] [CrossRef]
- Iqbal, M.; Zafar, S.I. Effects of photon flux density, CO2, aeration rate, and inoculum density on growth and extracellular polysaccharide production by Porphyridium cruentum. Folia Microbiol. 1993, 38, 509–514. [Google Scholar] [CrossRef]
- Bergman, B. Glyoxylate induced changes in the carbon and nitrogen metabolism of the cyanobacterium Anabaena cylindrica. Plant Physiol. 1986, 80, 698–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, A.; Liu, Y.; Ge, J.; Wang, W.; Chen, Y.; Montagnes, D. Aggregate formation and polysaccharide content of Chlorella pyrenoidosa Chick (Chlorophyta) in response to simulated nutrient stress. Bioresour. Technol. 2010, 101, 8336–8341. [Google Scholar] [CrossRef] [PubMed]
- De Philippis, R.; Sili, C.; Vincenzini, M. Response of an exopolysaccharide-producing heterocystous cyanobacterium to changes in metabolic carbon flux. J. Appl. Phycol. 1996, 8, 275–281. [Google Scholar] [CrossRef]
- Tischer, R.G.; Davis, E.B. The effect of various nitrogen sources upon the production of extracellular polysaccharide by the blue-green alga Anabaena flos-aquae A-37. J. Exp. Bot. 1971, 22, 546–551. [Google Scholar] [CrossRef]
- Roux, J.M. Production of polysaccharide slime by microbial mats in the hypersaline environment of a Western Australian solar salt field. Int. J. Salt Lake Res. 1996, 5, 103–130. [Google Scholar] [CrossRef]
- Fresnedo, O.; Serra, J.L. Effect of nitrogen starvation on the biochemistry of Phormidium laminosum (Cyanophyceae). J. Phycol. 1992, 28, 786–793. [Google Scholar] [CrossRef]
- Boonchai, R.; Kaewsuk, J.; Seo, G. Effect of nutrient starvation on nutrient uptake and extracellular polymeric substance for microalgae cultivation and separation. Desal Water Treat 2014, 55, 360–367. [Google Scholar] [CrossRef]
- Urbani, R.; Magaletti, E.; Sist, P.; Cicero, A.M. Extracellular carbohydrates released by the marine diatoms Cylindrotheca closterium, Thalassiosira pseudonana and Skeletonema costatum: Effect of P-depletion and growth status. Sci. Total Environ. 2005, 353, 300–306. [Google Scholar] [CrossRef]
- Guerrini, F.; Cangini, M.; Boni, L.; Trost, P.; Pistocchi, R. Metabolic responses of the diatom Achnanthes brevipes (Bacillariophyceae) to nutrient limitation. J. Phycol. 2000, 36, 882–890. [Google Scholar] [CrossRef]
- De Philippis, R.; Sili, C.; Tassinato, G.; Vincenzini, M.; Materassi, R. Effects of growth conditions on exopolysaccharide production by Cyanospira capsulata. Biores. Technol. 1991, 38, 101–104. [Google Scholar] [CrossRef]
- Shnyukova, E.I.; Zolotareva, E.K. Diatom exopolysaccharides: A review. Int. J. Algae 2015, 17, 50–67. [Google Scholar] [CrossRef]
- Vincenzini, M.; De Philippis, R.; Sili, C.; Materassi, R. Studies on exopolysaccharide release by diazotrophic batch cultures of Cyanospira capsulata. Appl. Microbiol. Biotechnol. 1990, 34, 392–396. [Google Scholar] [CrossRef]
- Fernandes, H.L.; Tomme, M.M.; Lupi, F.M.; Fialho, A.M.; Sa-Correia, I.; Novais, J.M. Biosynthesis of high concentration of an exopolysaccharide during cultivation of the microalga Botryococcus braunii. Biotechnol. Lett. 1989, 11, 433–436. [Google Scholar] [CrossRef]
- Myklestad, S.; Holm-Hansen, O.; Vårum, K.M.; Volcani, B.E. Rate of release of extracellular amino acids and carbohydrates from the marine diatom Chaetoceros affinis. J. Plankton Res. 1989, 11, 763–773. [Google Scholar] [CrossRef]
- Gantar, M.; Rowell, P.; Kerby, N.W.; Sutherland, I.W. Role of extracellular polysaccharide in the colonization of wheat (Triticum vulgare L.) roots by N2-fixing cyanobacteria. Biol. Fertil. Soils 1995, 19, 41–48. [Google Scholar] [CrossRef]
- Mehta, V.B.; Vaidya, B.S. Cellular and extracellular polysaccharides of the bluegreen alga Nostoc. J. Exp. Bot. 1978, 29, 1423–1430. [Google Scholar] [CrossRef]
- Mishra, A.; Jha, B. Isolation and characterization of extracellular polymeric substances from micro-algae Dunaliella salina under salt stress. Biores. Technol. 2009, 100, 3382–3386. [Google Scholar] [CrossRef]
- Chen, L.Z.; Li, D.H.; Song, L.R.; Hu, C.X.; Wang, G.H.; Liu, Y.D. Effects of salt stress on carbohydrate metabolism in desert soil alga Microcoleus vaginatus. J. Integr. Plant Biol. 2006, 48, 914–919. [Google Scholar] [CrossRef]
- Ozturk, S.; Aslim, B. Modification of exopolysaccharide composition and production by three cyanobacterial isolates under salt stress. Environ. Sci. Pollut. Res. 2010, 17, 595–602. [Google Scholar] [CrossRef]
- Fereira, A.S.; Mendonça, I.; Povoa, I.; Carvalho, H.; Correia, A.; Vilanova, M.; Silva, T.H.; Coimbra, M.A.; Nunes, C. Impact of growth medium salinity on galactoxylan exopolysaccharides of Porphyridium purpureum. Algal Res. 2021, 59, 102439. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Bilger, W.; Scherer, S. UV-B-induced synthesis of photoprotective pigments and extracellular polysaccharides in the terrestrial cyanobacterium Nostoc commune. J. Bacteriol. 1997, 179, 1940–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, P.-P.; Shen, S.-G.; Wang, H.-Y.; Sun, Y.; Dai, Y.-J.; Jia, S.-R. Comparative metabolomic analysis of the effects of light quality on polysaccharide production of cyanobacterium Nostoc flagelliforme. Algal Res. 2015, 9, 143–150. [Google Scholar] [CrossRef]
- Farges, B.; Laroche, C.; Cornet, J.-F.; Dussap, C.-G. Spectral kinetic modelling and long-term behavior assessment of Arthrospira platensis growth in photobioreactor under red (620 nm) light illumination. Biotechnol. Prog. 2009, 25, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Lupi, F.M.; Fernandes, H.M.L.; Sá-Correia, I.; Novais, J.M. Temperature profiles of cellular growth and exopolysaccharide synthesis by Botryococus braunii Kütz UC 58. J. Appl. Phycol. 1991, 3, 35–42. [Google Scholar] [CrossRef]
- Deamici, K.M.; de Morais, M.G.; Santos, L.O.; Muylaert, K.; Gardarin, C.; Costa, J.A.V.; Laroche, C. Static magnetic fields effects on polysaccharides production by different microalgae strains. Appl. Sci. 2021, 11, 5299. [Google Scholar] [CrossRef]
- Gonzalez-Fernandez, C.; Ballestros, M. Linking microalgae and cyanobacteria culture conditions and key-enzymes for carbohydrate accumulation. Biotechnol. Adv. 2012, 30, 1655–1661. [Google Scholar] [CrossRef] [PubMed]
- Ball, S.G.; Dirick, L.; Decq, A.; Martiat, J.-C.; Matagne, R. Physiology of starch storage in the monocellular alga Chlamydomonas reinhardtii. Plant Sci. 1990, 66, 1–9. [Google Scholar] [CrossRef]
- Razaghi, A.; Godhe, A.; Albers, E. Effects of nitrogen on growth and carbohydrate formation in Porphyridium cruentum. Cent. Eur. J. Biol. 2014, 9, 156–162. [Google Scholar] [CrossRef]
- Zhang, S.J.; Santschi, P.H. Application of cross-flow ultrafiltration for isolating exopolymeric substances from a marine diatom (Amphora sp.). Limnol. Oceanogr. Meth. 2009, 7, 419–429. [Google Scholar] [CrossRef]
- Li, H.; Li, Z.; Xiong, S.; Zhang, H.; Li, N.; Zhou, S.; Liu, Y.; Huang, Z. Pilot-scale isolation of bioactive extracellular polymeric substances from cell-free media of mass microalgal cultures using tangential-flow ultrafiltration. Process. Biochem. 2011, 46, 1104–1109. [Google Scholar] [CrossRef]
- Ye, Y.; Clech, P.L.; Chen, V.; Fane, A.G. Evolution of fouling during crossflow filtration of model EPS solutions. J. Membr. Sci. 2005, 264, 190–199. [Google Scholar] [CrossRef]
- Marcati, A.; Ursu, A.V.; Laroche, C.; Soanen, N.; Marchal, L.; Jubeau, S.; Djelveh, G.; Michaud, P. Extraction and fractionation of polysaccharides and B-phycoerythrin from the microalga Porphyridium cruentum by membrane technology. Algal Res. 2014, 5, 258–263. [Google Scholar] [CrossRef]
- Patel, A.K.; Laroche, C.; Marcati, A.; Ursu, A.V.; Jubeau, S.; Marchal, L.; Petit, E.; Djelveh, G.; Michaud, P. Separation and fractionation of exopolysaccharides from Porphyridium cruentum. Biores. Technol. 2013, 145, 345–350. [Google Scholar] [CrossRef] [PubMed]
- De Brouwer, J.F.C.; Stal, L.J. Does warm-water extraction of benthic diatoms yield extracellular polymeric substances or does it extract intracellular chrysolaminaran? Eur. J. Phycol. 2004, 39, 129–131. [Google Scholar] [CrossRef]
- Rickman, M.; Pellegrino, J.; Davis, R. Fouling phenomena during membrane filtration of microalgae. J. Membr. Sci. 2012, 423–424, 33–42. [Google Scholar] [CrossRef]
- Jhaveri, J.H.; Murthy, Z.V.P. A comprehensive review on anti-fouling nanocomposite membranes for pressure driven membrane separation processes. Desalination 2016, 379, 137–154. [Google Scholar] [CrossRef]
- Rossi, N.; Jaouen, P.; Legentilhomme, P.; Petit, I. Harvesting of cyanobacterium Arthrospira platensis using organic filtration membranes. Food Bioprod. Process. 2004, 82, 244–250. [Google Scholar] [CrossRef]
- Rossi, N.; Petit, I.; Jaouen, P.; Legentilhomme, P.; Derouiniot, M. Harvesting of cyanobacterium Arthrospira platensis using inorganic filtration membranes. Sep. Sci. Technol. 2005, 40, 3033–3050. [Google Scholar] [CrossRef]
- Brown, M.J.; Lester, J.N. Comparison of bacterial extracellular polymer extraction methods. Appl. Environ. Microbiol. 1980, 40, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Comte, S.; Guibaud, G.; Baudu, M. Relations between extraction protocols for activated sludge extracellular polymeric substances (EPS) complexation properties: Part I Comparison of the efficiency of eight EPS extraction methods. Enzym. Microb. Technol. 2006, 38, 237–245. [Google Scholar] [CrossRef]
- Frølund, B.; Palmgren, R.; Keiding, K.; Nielsen, P.H. Extraction of extracellular polymers from activated sludge using a cation exchange resin. Water Res. 1996, 30, 1749–1758. [Google Scholar] [CrossRef]
- Ogutu, F.; Mu, T.-H.; Elahi, R.; Zhang, M.; Sun, H.-N. Ultrasonic modification of selected polysaccharides—Review. Food Process. Technol. 2015, 6, 446–453. [Google Scholar] [CrossRef] [Green Version]
- Aida, T.M.; Yamagata, T.; Watanabe, M.; Smith, R.L. Depolymerization of sodium alginate under hydrothermal conditions. Carbohydr. Polym. 2010, 80, 296–302. [Google Scholar] [CrossRef]
- Chiovitti, A.; Molino, P.; Crawford, S.A.; Ten, R.; Spurck, T.; Wetherbee, R. The glucans extracted with warm water from diatoms are mainly derived from intracellular chrysolaminaran and not extracellular polysaccharides. Eur. J. Phycol. 2004, 39, 117–128. [Google Scholar] [CrossRef]
- Takahashi, E.; Ledauphin, J.; Goux, D.; Orvain, F. Optimizing extraction of extracellular polymeric substances (EPS) from benthic diatoms: Comparison of the efficiency of six EPS extraction methods. Austr. J. Mar. Freshwat. Res. 2009, 60, 1201–1210. [Google Scholar] [CrossRef] [Green Version]
- De Brouwer, J.F.C.; Wolfstein, K.; Stal, L.J. Physical characterization and diel dynamics of different fractions of extracellular polysaccharides in an axenic culture of a benthic diatom. Eur. J. Phycol. 2002, 37, 37–44. [Google Scholar] [CrossRef]
- Cloete, T.E.; Jacobs, L.; Brozel, V.S. The chemical control of biofouling in industrial water systems. Biodegradation 1998, 9, 23–37. [Google Scholar] [CrossRef] [Green Version]
- McDonnell, G.; Russell, A.D. Antiseptics and disinfectants: Activity, action and resistance. Clin. Microbiol. Rev. 1999, 12, 147–179. [Google Scholar] [CrossRef] [Green Version]
- Azeredo, J.; Henriques, M.; Sillankorva, S.; Oliveira, R. Extraction of exopolymers from biofilms: The protective effect of glutaraldehyde. Water Sci. Technol. 2003, 47, 175–179. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Fang, H.P. Extraction of extracellular polymeric substances (EPS) of sludges. J. Biotechnol. 2002, 95, 249–256. [Google Scholar] [CrossRef]
- Herrero, M.; Martin-Alvarez, P.J.; Senoráns, F.J.; Cifuentes, A.; Ibánez, E. Optimization of accelerated solvent extraction of antioxidants from Spirulina platensis microalga. Food Chem. 2005, 93, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Gao, Y.T.; Dai, Y.; Yang, Y.L.; Wang, X.M. Scavenging effects of Spirulina and polysaccharides Spirulina platensis on active oxygens and its antioxidation in vitro. Chem. Bioeng. 2007, 24, 55–57. [Google Scholar]
- Sun, L.; Wang, C.; Shi, Q.; Ma, C. Preparation of different molecular weight polysaccharides from Porphyridium cruentum and their antioxidant activities. Int. J. Biol. Macromol. 2009, 45, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Tannin-Spitz, T.; Bergman, M.; van-Moppes, D.; Grossman, S.; Arad, S.M. Antioxidant activity of the polysaccharide of the red microalga Porphyridium sp. J. Appl. Phycol. 2005, 17, 215–222. [Google Scholar] [CrossRef]
- Chen, B.; You, W.; Huang, J.; Yu, Y.; Chen, W. Isolation and antioxidant property of the extracellular polysaccharide from Rhodella reticulata. World J. Microbiol. Biotechnol. 2010, 26, 833–840. [Google Scholar] [CrossRef]
- Wang, L.; Sun, L.Q.; Zhao, X.H. Physicochemical property analysis of polysaccharides from marine microalgae and their antioxidation, hygroscopicity and moisture retention activities. Fine Chem. 2012, 1, 20–25. [Google Scholar]
- Wang, S.Y.; Jiang, Y.; Meng, C.; OuYang, Y.H.; Lin, X.Z. Preparation of extracellular polysaccharide from fermentation liquor of marine microalgae Schizochytrium and study on the bioactivities. J. Fuzhou Univ. 2011, 39, 786–791. [Google Scholar]
- Sun, Y.; Wang, H.; Guo, G.; Pu, Y.; Yan, B. The isolation and antioxidant activity of polysaccharides from the marine microalgae Isochrysis galbana. Carbohydr. Polym. 2014, 113, 22–31. [Google Scholar] [CrossRef]
- Hussein, M.H.; Abou-ElWafa, G.S.; Shaaban-Dessuuki, S.A.; Hassan, N.I. Characterization and Antioxidant Activity of Exopolysaccharide Secreted by Nostoc carneum. Int. J. Pharmacol. 2015, 11, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Gargouch, N.; Elleuch, F.; Karkouch, I.; Tabbene, O.; Pichon, C.; Gardarin, C.; Rihouey, C.; Picton, L.; Abdelkafi, S.; Fendri, I.; et al. Potential of exopolysaccharide from Porphyridium marinum to contend with bacterial proliferation, biofilm formation, and breast cancer. Mar. Drugs 2021, 19, 66. [Google Scholar] [CrossRef]
- Huheihel, M.; Ishanu, V.; Tal, J.; Arad, S.M. Activity of Porphyridium sp. polysaccharide against Herpes simplex viruses in vitro and in vivo. J. Biochem. Biophys. Meth. 2002, 50, 189–200. [Google Scholar] [CrossRef]
- Huheihel, M.; Ishanu, V.; Tal, J.; Arad, S.M. Antiviral effect of red microalgal polysaccharides on Herpes simplex and Varicella zoster viruses. J. Appl. Phycol. 2001, 13, 127–134. [Google Scholar] [CrossRef]
- Hayashi, K.; Hayashi, T.; Kojima, I. A natural sulphated polysaccharide, calcium spirulan, isolated from Spirulina platensis: In vitro and ex vivo evaluation of anti-herpes simplex virus and anti-human immunodeficiency virus. AIDS Res. Hum. Retrovir. 1996, 12, 1463–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, T.; Hayashi, K.; Maeda, M.; Kojima, I. Calcium spirulan, an inhibitor of enveloped virus replication, from a blue-green alga Spirulina platensis. J. Nat. Prod. 1996, 59, 83–87. [Google Scholar] [CrossRef]
- Rechter, S.; König, T.; Auerochs, S.; Thulke, S.; Walter, H.; Dörnenburg, H.; Walter, C.; Marschall, M. Antiviral activity of Arthrospira-derived spirulan-like substances. Antivir. Res. 2006, 72, 197–206. [Google Scholar] [CrossRef]
- Sun, L.; Wang, L.; Zhou, Y. Immunomodulation and antitumor activities of different-molecular-weight polysaccharides from Porphyridium cruentum. Carbohydr. Polym. 2012, 87, 1206–1210. [Google Scholar] [CrossRef]
- Yim, J.H.; Son, E.; Pyo, S.; Lee, H.K. Novel sulfated polysaccharide derived from red-tide microalga Gyrodinium impudicum strain KG03 with immunostimulating activity in vivo. Mar. Biotechnol. 2005, 7, 331–338. [Google Scholar] [CrossRef]
- Risjani, Y.; Mutmainnah, N.; Manurung, P.; Wulan, S.N.; Yunianta. Exopolysaccharide from Porphyridium cruentum (purpureum) is Not Toxic and Stimulates Immune Response against Vibriosis: The Assessment Using Zebrafish and White Shrimp Litopenaeus vannamei. Mar. Drugs 2021, 19, 133. [Google Scholar] [CrossRef]
- Uhliarikova, I.; Šutovská, M.; Barboríková, J.; Molitorisová, M.; Kim, H.J.; Park, Y.; Matulová, M.; Lukavský, J.; Hromadková, Z.; Capek, P. Structural characteristics and biological effects of exopolysaccharide produced by cyanobacterium Nostoc sp. Int. J. Biol. Macromol. 2020, 160, 364–371. [Google Scholar] [CrossRef]
- Matsui, M.S.; Muizzuddin, N.; Arad, S.M.; Marenus, K. Sulfated polysaccharides from red microalgae have antiinflammatory properties in vitro and in vivo. Appl. Biochem. Biotechnol. 2003, 104, 13–22. [Google Scholar] [CrossRef]
- Nosálová, G.; Capek, P.; Matáková, T.; Nosál, S.; Flešková, D.; Jureček, L. Antitussive activity of an extracellular Rhodella grisea proteoglycan on the mechanically induced cough reflex. Carbohydr. Polym. 2012, 87, 752–756. [Google Scholar] [CrossRef] [PubMed]
- Hayman, J.R.; Southern, T.R.; Nash, T.E. Role of sulfated glycans in adherence of the microsporidian Encephalitozoon intestinalis to host cells in vitro. Inf. Immun. 2005, 73, 841–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hintze, V.; Miron, A.; Moeller, S.; Schnabelrauch, M.; Wiesmann, H.P.; Worch, H.; Scharnweber, D. Sulfated hyaluronan and chondroitin sulfate derivatives interact differently with human transforming growth factor-β1 (TGF-β1). Acta Biomater. 2012, 8, 2144–2152. [Google Scholar] [CrossRef] [PubMed]
- Kreuger, J.; Spillmann, D.; Li, J.P.; Lindahl, U. Interactions between heparan sulfate and proteins: The concept of specificity. J. Cell Biol. 2006, 174, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Corsuto, L.; Rother, S.; Koehler, L.; Bedini, E.; Moeller, S.; Schnabelrauch, M.; Hintze, V.; Schiraldi, C.; Scharnweber, D. Sulfation degree not origin of chondroitin sulfate derivatives modulates keratinocyte response. Carbohydr. Polym. 2018, 191, 53–64. [Google Scholar] [CrossRef]
- Martinez, M.J.A.; Del Olmo, L.M.B.; Benito, P.B. Antiviral activities from natural sources. In Studies in Natural Products Chemistry; Atta-ur-Rahman, B.V., Ed.; Elsevier: London, UK, 2005; Volume 30, pp. 393–418. [Google Scholar]
- Kim, M.; Yim, J.H.; Kim, S.-Y.; Kim, H.S.; Lee, W.G.; Kim, S.J.; Kang, P.S.; Lee, C.K. In vitro inhibition of influenza A virus infection by marine microalga-derived polysaccharide p-KG03. Antivir. Res. 2012, 93, 253–259. [Google Scholar] [CrossRef]
- Damonte, E.B.; Matulewicz, M.C.; Cerezo, A.S. Sulfated seaweed polysaccharides as antiviral agents. Curr. Med. Chem. 2004, 11, 2399–2419. [Google Scholar] [CrossRef]
- Radonic, A.; Thulke, S.; Achenbach, J.; Kurth, A.; Vreemann, A.; König, T.; Walter, C.; Possinger, K.; Nitsche, A. Anionic polysaccharides from phototrophic microorganisms exhibit antiviral activities to Vaccinia virus. J. Antivir. Antiretrovir. 2010, 2, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Rashid, Z.M.; Lahaye, E.; Defer, D.; Douzenel, P.; Perrin, B.; Bourgougnon, N.; Sire, O. Isolation of a sulfated polysaccharide from a recently discovered sponge species (Celtodoryx girardae) and determination of its anti-herpetic activity. Int. J. Biol. Macromol. 2009, 44, 286–293. [Google Scholar] [CrossRef]
- Kanekiyo, K.; Hayashi, K.; Takenaka, H.; Lee, J.B.; Hayashi, T. Anti-herpes simplex virus target of an acidic polysaccharide, Nostoflan, from the edible blue-green alga Nostoc flagelliforme. Biol. Pharmac. Bull 2007, 30, 1573–1575. [Google Scholar] [CrossRef] [Green Version]
- Ropellato, J.; Carvalho, M.M.; Ferreira, L.G.; Noseda, M.D.; Zuconelli, C.R.; Gonçalves, A.G.; Ducatti, D.R.B.; Kenski, J.C.N.; Nasato, P.L.; Winnischofer, S.M.B.; et al. Sulfated heterorhamnans from the green seaweed Gayralia oxysperma: Partial depolymerization, chemical structure and antitumor activity. Carbohydr. Polym. 2015, 117, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, Y.; Hayashi, T.; Lee, J.-B.; Srisomporn, P.; Maeda, M.; Ozawa, T.; Sakuragawa, N. Inhibition of thrombin by sulfated polysaccharides isolated from green algae. Biochim. Biophys. Acta 2000, 1543, 86–94. [Google Scholar] [CrossRef]
- Funami, T. In vivo and rheological approaches for characterizing food oral processing and usefulness of polysaccharides as texture modifiers—A review. Food Hydrocoll. 2017, 68, 2–14. [Google Scholar] [CrossRef]
- Geresh, S.; Mamontov, A.; Weinstein, J. Sulfation of extracellular polysaccharides of red microalgae: Preparation, characterization and properties. J. Biochem. Biophys. Meth. 2002, 50, 179–187. [Google Scholar] [CrossRef]
- Geresh, S.; Arad, S.M. The extracellular polysaccharides of the red microalgae: Chemistry and rheology. Biores. Technol. 1991, 38, 195–201. [Google Scholar] [CrossRef]
- Eteshola, E.; Karpasas, M.; Arad, S.M.; Gottlieb, M. Red microalga exopolysaccharides: 2. Study of the rheology, morphology and thermal gelation of aqueous preparations. Acta Polym. 1998, 49, 549–556. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; de Morais, R.M.S.C.; de Morais, A.M.M.B. Bioactivity and applications of sulphated polysaccharides from marine microalgae. Mar. Drugs 2013, 11, 233–252. [Google Scholar] [CrossRef] [Green Version]
- Imeson, A. Thickening and Gelling Agents for Food, 2nd ed.; Springer: New York, NY, USA, 1997; p. 320. [Google Scholar]
- Imeson, A. Food Stabilisers, Thickeners and Gelling Agents; John Wiley & Sons: Chichester, UK, 2011; p. 354. [Google Scholar]
- Rao, M.A. Rheology of Fluid and Semisolid Foods: Principles and Applications, 2nd ed.; Springer: New York, NY, USA, 2010; p. 481. [Google Scholar]
- Medina-Cabrera, V.; Gansbiller, M.; Rühmann, B.; Schmid, J.; Sieber, V. Rheological characterization of Porphyridium sordidum and Porphyridium purpureum exopolysaccharides. Carbohydr. Polym. 2021, 253, 117237. [Google Scholar] [CrossRef]
- Arad, S.M.; Rapoport, L.; Moshkovich, A.; van Moppes, D.; Karpasas, M.; Golan, R.; Golan, Y. Superior biolubricant from a species of red microalga. Langmuir 2006, 22, 7313–7317. [Google Scholar] [CrossRef]
- Ginzberg, A.; Korin, E.; Arad, S.M. Effect of drying on the biological activities of a red microalga polysaccharide. Biotechnol. Bioeng. 2008, 99, 411–420. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; de Morais, A.M.M.B.; de Morais, R.M.S.C. Bioactivity and applications of polysaccharides from marine microalgae. In Polysaccharides; Ramawat, K., Mérillon, J.M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 1683–1727. [Google Scholar]
- Vincenzini, M.; De Philippis, R.; Sili, C.; Materassi, R. A novel exopolysaccharide from a filamentous cyanobacterium: Production, chemical characterization and theological properties. In Novel Biodegradable Microbial Polymers; Dawes, E.A., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990; pp. 295–310. [Google Scholar]
- Han, P.-P.; Sun, Y.; Wu, X.-Y.; Yuan, Y.-J.; Dai, Y.-J.; Jia, S.-R. Emulsifying, flocculating, and physicochemical properties of exopolysaccharide produced by cyanobacterium Nostoc flagelliforme. Appl. Biochem. Biotechnol. 2014, 172, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Chentir, I.; Hamdi, M.; Doumandji, A.; HadjSadok, A.; Ben Ouada, H.; Nasri, M.; Jridi, M. Enhancement of extracellular polymeric substances (EPS) production in Spirulina (Arthrospira sp.) by two-step cultivation process and partial characterization of their polysaccharidic moiety. Int. J. Biol. Macromol. 2017, 105, 1412–1420. [Google Scholar] [CrossRef] [PubMed]
- Urrutia, M.M. General bacteria sorption processes. In Biosorbents for Metal Ions; Wase, J., Forster, C., Eds.; Taylor & Francis: London, UK, 1997; pp. 39–66. [Google Scholar]
- De Philippis, R.; Paperi, R.; Sili, C. Heavy metal sorption by released polysaccharides and whole cultures of two exopolysaccharide-producing cyanobacteria. Biodegradation 2007, 18, 181–187. [Google Scholar] [CrossRef] [PubMed]
- De Philippis, R.; Colica, G.; Micheletti, E. Exopolysaccharide-producing cyanobacteria in heavy metal removal from water: Molecular basis and practical applicability of the biosorption process. Appl. Microbiol. Biotechnol. 2011, 92, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Micheletti, E.; Colica, G.; Viti, C.; Tamagnini, P.; De Philippis, R. Selectivity in the heavy metal removal by exopolysaccharide-producing cyanobacteria. J. Appl. Microbiol. 2008, 105, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, R.; Rockey, J.; Sutherland, I.W.; Roller, S. Novel bioemulsifiers from microorganisms for use in foods. J. Biotechnol. 1995, 40, 207–217. [Google Scholar] [CrossRef]
- Paniagua-Michel, J.; Olmos-Soto, J.; Morales-Guerrero, E.R. Algal and Microbial Exopolysaccharides: New Insights as Biosurfactants and Bioemulsifiers. Adv. Food Nut. Res. 2014, 73, 221–257. [Google Scholar]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar applications of protein hydrolysate, plant and seaweed extracts increase yield but differentially modulate fruit quality of greenhouse tomato. Hort. Sci. 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- Khan, W.; Hiltz, D.; Critchley, A.T.; Prithiviraj, B. Bioassay to detect Ascophyllum nodosum extract-induced cytokinin-like activity in Arabidopsis thaliana. J. Appl. Phycol. 2011, 23, 409–414. [Google Scholar] [CrossRef]
- El Arroussi, H.; Elbaouchi, A.; Benhima, R.; Bendaou, N.; Smouni, A.; Wahby, I. Halophilic microalgae Dunaliella salina extracts improve seed germination and seedling growth of Triticum aestivum L. under salt stress. Acta Hortic. 2016, 1148, 13–26. [Google Scholar] [CrossRef]
- Rachidi, F.; Benhima, R.; Sbabou, L.; El Arroussi, H. Microalgae polysaccharides bio-stimulating effect on tomato plants: Growth and metabolic distribution. Biotechnol. Rep. 2020, 25, e00426. [Google Scholar] [CrossRef] [PubMed]
- Plaza, B.M.; Gómez-Serrano, C.; Acién-Fernández, F.G.; Jimenez-Becker, S. Effect of microalgae hydrolysate foliar application (Arthrospira platensis and Scenedesmus sp.) on Petunia x hybrida growth. J. Appl. Phycol. 2018, 30, 2359–2365. [Google Scholar] [CrossRef]
- Barone, V.; Baglieri, A.; Stevanato, P.; Broccanello, C.; Bertoldo, G.; Bertaggia, M.; Cagnin, M.; Pizzeghello, D.; Moliterni, V.M.C.; Mandolino, G.; et al. Root morphological and molecular responses induced by microalgae extracts in sugar beet (Beta vulgaris L.). J. Appl. Phycol. 2017, 30, 1061–1071. [Google Scholar] [CrossRef]
- Guzmán-Murillo, M.A.; Ascencio, F.; Larrinaga-Mayoral, J.A. Germination and ROS detoxification in bell pepper (Capsicum annuum L.) under NaCl stress and treatment with microalgae extracts. Protoplasma 2012, 250, 33–42. [Google Scholar] [CrossRef] [PubMed]
- El Arroussi, H.; Benhima, R.; Elbaouchi, A.; Sijilmassi, B.; El Mernissi, N.; Aafsar, A.; Meftah-Kadmiri, I.; Bendou, N.; Smouni, A. Dunaliella salina exopolysaccharides: A promising biostimulant for salt stress tolerance in tomato (Solanum lycopersicum). J. Appl. Phycol. 2018, 30, 2929–2941. [Google Scholar] [CrossRef]
- Rachidi, F.; Chanda, M.-J.; Benhima, R.; El Nernissi, N.; Aasfar, A.; Sbabou, L.; El Aroussi, H. Efect of microalgae polysaccharides on biochemical and metabolomics pathways related to plant defense in Solanum lycopersicum. Appl. Biochem. Biotechnol. 2019, 188, 225–240. [Google Scholar]
- Gupta, P.; Ravi, I.; Sharma, V. Induction of β-1, 3-glucanase and chitinase activity in the defense response of Eruca sativa plants against the fungal pathogen Alternaria brassicicola. J. Plant Interact. 2013, 8, 155–161. [Google Scholar] [CrossRef]
- Fesel, P.H.; Zuccaro, A. β-glucan: Crucial component of the fungal cell wall and elusive MAMP in plants. Fungal Genet. Biol. 2016, 90, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Bixler, H.J.; Porse, H. A decade of change in the seaweed hydrocolloids industry. J. Appl. Phycol. 2011, 23, 321–335. [Google Scholar] [CrossRef]
- Potter, A.; Thibaut, S.; Ribaut, C. Utilisation De Polysaccharides Sulfatés Comme Agent Antipelliculaire. WO Patent 2013093307A1, 27 June 2013. [Google Scholar]
- Li, H.; Xu, J.; Liu, Y.; Ai, S.; Qin, F.; Li, Z.; Zhang, H.; Huang, Z. Antioxidant and moisture-retention activities of the polysaccharide from Nostoc commune. Carbohydr. Polym. 2011, 83, 1821–1827. [Google Scholar] [CrossRef]
- Okajima, M.K.; Bamba, T.; Kaneso, Y.; Hirata, K.; Fukusaki, E.; Kajiyama, S.I.; Kaneko, T. Supergiant ampholytic sugar chains with imbalanced charge ratio form saline ultra-absorbent hydrogels. Macromolecules 2008, 41, 4061–4064. [Google Scholar] [CrossRef]
- Okajima, M.K.; Miyazato, S.; Kaneko, T. Cyanobacterial megamolecule sacran efficiently forms LC gels with very heavy metal ions. Langmuir 2009, 25, 8526–8531. [Google Scholar] [CrossRef] [PubMed]
- Coragliotti, A.; Franklin, S.; Day, A.G.; Decker, S.M. Microalgal Polysaccharide Compositions. U.S. Patent 2012/0202768, 9 August 2012. [Google Scholar]
- Yanhui, C. Cosmetic Composition with Functions of Repairing and Strengthening Skin Barrier and Application. CN Patent 107412042A, 12 August 2017. [Google Scholar]
- Potter, A.; Ghibaudo, M.; Baltenneck, C. Association De Polysaccharides Sulfatés Et De C-Glycoside Et Leurs Utilisations. WO Patent 2014174188A1, 30 October 2014. [Google Scholar]
- Bayona, K.C.D.; Gallón, S.M.N.; Lara, D.R.; Colorado, J.R. Activity of sulfated polysaccharides from microalgae Porphyridium cruentum over degenerative mechanisms of the skin. Int. J. Sci. Adv. Technol. 2012, 2, 85–92. [Google Scholar]
- Dillon, H.; Somanchi, A.; Zaman, A.; Rao, K.; Wolfson, J. Methods and Compositions for Improving the Health and Appearance of Skin. U.S. Patent 20070166266A1, 19 July 2007. [Google Scholar]
- Gonçalves, O.; Pruvost, J.; Massé, A.; Decamp, A.; Probert, I.; Maugard, T.; Arnaudin, I.; Bridiau, N.; Toucheteau, C.; Le Cerf, D.; et al. Nouveaux Exopolysaccharides Dépolymérisés, Issus De Micro-Algues, Leur Procédé De Préparation Et Leurs Utilisations En Cosmétique Pour Retarder Les Effets Du Vieillissement Cutané. FR Patent 2102020, 25 October 2021. [Google Scholar]
- Bernal, J.; Mendiola, J.A.; Ibáñez, E.; Cifuentes, A. Advanced analysis of nutraceuticals. J. Pharm. Biomed. Anal. 2011, 55, 758–774. [Google Scholar] [CrossRef] [Green Version]
- Arad, S.M.; Keristovesky, G.; Simon, B.; Barak, Z.; Geresh, S. Biodegradation of the sulphated polysaccharide of Porphyridium by soil bacteria. Phytochemistry 1993, 32, 287–290. [Google Scholar] [CrossRef]
- Dvir, I.; Stark, A.H.; Chayoth, R.; Madar, Z.; Arad, S.M. Hypocholesterolemic effects of nutraceuticals produced from the red microalga Porphyridium sp. in Rats. Nutrients 2009, 1, 156–167. [Google Scholar] [CrossRef] [Green Version]
- Dillon, H.; Somanchi, A.; Rao, K. Methods and Compositions for Cholesterol Reduction in Mammals. U.S. Patent 20070167396A1, 19 July 2007. [Google Scholar]
- Dillon, H.; Somanchi, A.; Rao, K. Methods and Compositions for Reducing Inflammation and Preventing Oxidative Damage. U.S. Patent 20070167398A1, 19 July 2007. [Google Scholar]
- Huang, J.; Liu, L.; Yu, Y.; Lin, W.; Chen, B.; Li, M.; Rao, X. Reduction in the blood glucose level of exopolysaccharide of Porphyridium cruentum in alloxan-induced diabetic mice. J. Fujian Norm. Univ. 2006, 22, 77–80. [Google Scholar]
- Arad, S.M.; Huliheil, M.; Tal, J. Antiviral Agents. WO Patent 1997000689A1, 1 September 1997. [Google Scholar]
- Arad, S.M.; Atar, D. Viscosupplementation with Algal Polysaccharides in the Treatment of Arthritis. U.S. Patent 9119870B2, 14 June 2006. [Google Scholar]
- Michaud, P.; Laroche, C.; Villay, A.; Roussel, M.; Diogon, M.; El Alaoui, H.; Delbac, F. Sulphated Polysaccharide Composition. WO Patent 2014076261A1, 22 May 2014. [Google Scholar]
- Piacquadio, D.; Nelson, D.B. Alginates: A “New” Dressing Alternative. Dermato. Surg. 1992, 18, 992–995. [Google Scholar] [CrossRef]
- Sweeney, I.R.; Miraftab, M.; Collyer, G. A critical review of modern and emerging absorbent dressings used to treat exuding wounds. Int. Wound J. 2012, 9, 601–612. [Google Scholar] [CrossRef]
- Alvarez, X.; Alves, A.; Ribeiro, M.P.; Lazzari, M.; Coutinho, P.; Otero, A. Biochemical characterization of Nostoc sp. exopolysaccharides and evaluation of potential use in wound healing. Carbohydr. Polym. 2021, 254, 117303. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, M.; Parwani, L.; Sharma, V.; Ganguly, J.; Bhatnagar, A. Exopolymers from Tolypothrix tenuis and three Anabaena sp. (Cyanobacteriaceae) as novel blood clotting agents for wound management. Carbohydr. Polym. 2014, 99, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Kraan, S. Algal polysaccharides, novel applications and outlook. In Carbohydrates—Comprehensive Studies on Glycobiology and Glycotechnology; Chang, C.-F., Ed.; InTech: Rijeka, Croatia, 2012; pp. 489–532. [Google Scholar]
- Gonçalves, O.; Pruvost, J.; Massé, A.; Decamp, A.; Probert, I.; Maugard, T.; Arnaudin, I.; Bridiau, N.; Toucheteau, C.; Le Cerf, D.; et al. Utilisation D’exopolysaccharides De Micro-Algues A Titre D’agents Texturants. FR Patent 2102024, 25 October 2021. [Google Scholar]



| Organisms | Structures | References |
|---|---|---|
| C. augustae |  | [77] |
| C. corrosa |  | [77] |
| A. densus |  | [73] |
| P. cruentum, P. aerugineum |  | [58] |
| Porphyridium sp., P. cruentum, P. aerugineum, R. reticulata |  | [129] |
| Porphyridium sp. |  | [108] |
| Porphyridium sp. |  | [130] |
| Cyanothece sp. |  | [131] |
| C. capsulata |  | [132] |
| N. commune |  | [124] |
| N. insulare |  | [60] |
| O. planktothrix FP1 |  | [127] |
| M. laminosus |  | [21] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laroche, C. Exopolysaccharides from Microalgae and Cyanobacteria: Diversity of Strains, Production Strategies, and Applications. Mar. Drugs 2022, 20, 336. https://doi.org/10.3390/md20050336
Laroche C. Exopolysaccharides from Microalgae and Cyanobacteria: Diversity of Strains, Production Strategies, and Applications. Marine Drugs. 2022; 20(5):336. https://doi.org/10.3390/md20050336
Chicago/Turabian StyleLaroche, Céline. 2022. "Exopolysaccharides from Microalgae and Cyanobacteria: Diversity of Strains, Production Strategies, and Applications" Marine Drugs 20, no. 5: 336. https://doi.org/10.3390/md20050336
APA StyleLaroche, C. (2022). Exopolysaccharides from Microalgae and Cyanobacteria: Diversity of Strains, Production Strategies, and Applications. Marine Drugs, 20(5), 336. https://doi.org/10.3390/md20050336