
Marine Arthropods as a Source of Antimicrobial Peptides

 ,
,  ,
,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Bioactive Peptides in the Subphylum Crustacea
Diversity and Distribution
3. Defense Mechanisms in Crustaceans
3.1. Hemocytes
3.2. Cellular Reactions
3.3. Other Immune Tissues
3.4. Humoral Response
3.5. Recognition of Foreign Molecules
3.6. Chemical Structure, Properties and Function of AMPs
3.6.1. Penaeidins
Structure
Function and Role against Microorganisms
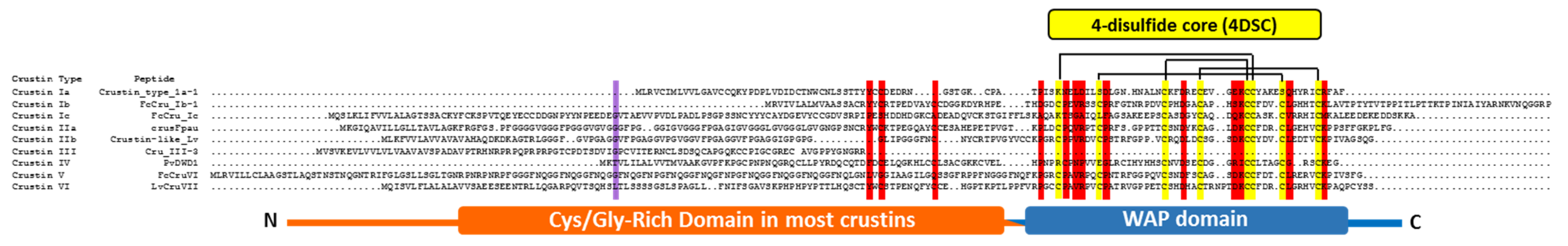
3.6.2. Crustins
Structure
Function and Role against Microorganisms
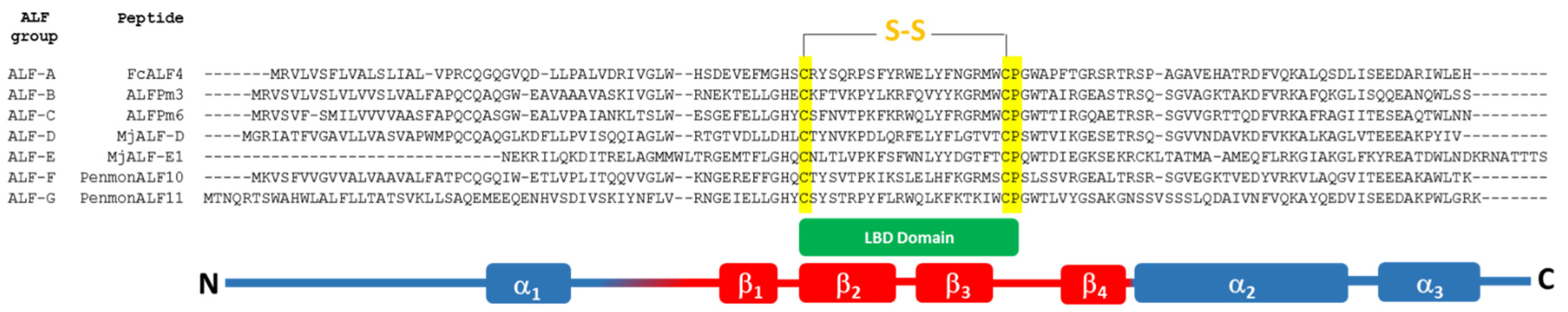
3.6.3. Anti-Lipopolysaccharide Factors (ALFs)
Structure
Function and Role against Microorganisms

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ALF | Peptide Name/Code | Shrimp Species | Activity | Reference |
|---|---|---|---|---|
| A | FcALF4 (JX853777) | Penaeus chinensis | Antiviral (WSSV) | [159] |
| ALFPm1 (EF523560) | Penaeus monodon | Not determined | [148] | |
| ALFPm2 (EF523561) | Penaeus monodon | Broad spectrum antibacterial | [148] | |
| B | ALFPm3 (EF523559) | Penaeus monodon | Antifungal | [148,160] |
| ALFFc (KJ004399) | Penaeus chinensis | Anti-Gram negative, antifungal | [28] | |
| C | ALFPm6 (AER45468) | Penaeus monodon | Broad spectrum antibacterial Antiviral | [161] |
| RspALF1 | Rimicaris sp. | Broad spectrum antibacterial Antifungal Antiviral | [150] | |
| MjALF2 (AB453738) | Penaeus japonicus | Not determined | [162] | |
| D | MjALF-D (MN416688) | Penaeus japonicus | Anti-Gram negative | [163] |
| PenmonALF8 | Penaeus monodon | Anti-Gram negative (moderate) | [151] | |
| E | MjALF-E1 | Penaeus japonicus | Anti-Gram negative | [164] |
| MjALF-E2 | Penaeus japonicus | Anti-Gram negative Promotes the clearance of bacteria in vivo | [164] | |
| F | ALFPm10 (XP_037782682) | Penaeus monodon | Anti-Gram positive | [46] [165] |
| G | ALFPm11 | Penaeus monodon | Broad spectrum antibacterial Antiviral | [152] |
3.6.4. Stylicins
Structure
Function and Role against Microorganisms
4. Marine Crustacean AMPs as Therapeutic Drugs
5. Antimicrobial Peptides from Chelicerates
6. Synthesis of Antimicrobial Peptides from Marine Arthropods
7. Conclusions and Future Perspectives
Funding
Conflicts of Interest
References
- Chaplin, D.D. Overview of the Immune Response. J. Allergy Clin. Immunol. 2010, 2 (Suppl. 2), 125. [Google Scholar] [CrossRef] [PubMed]
- Fosgerau, K.; Hoffmann, T. Peptide therapeutics: Current status and future directions. Drug Discov. Today 2015, 20, 122–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Gutierrez, E.; Mayer, M.J.; Cotter, P.D.; Narbad, A. Gut microbiota as a source of novel antimicrobials. Gut Microbes 2019, 10, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Aberturas, J.; Vior, N.M. Beyond Soil-Dwelling Actinobacteria: Fantastic Antibiotics and Where to Find Them. Antibiotics 2022, 11, 195. [Google Scholar] [CrossRef]
- D’Aloisio, V.; Dognini, P.; Hutcheon, G.A.; Coxon, C.R. PepTherDia: Database and structural composition analysis of approved peptide therapeutics and diagnostics. Drug Discov. Today 2021, 26, 1409–1419. [Google Scholar] [CrossRef] [PubMed]
- Mullard, A. Targeted protein degraders crowd into the clinic. Nat. Rev. Drug Discov. 2021, 20, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Muttenthaler, M.; King, G.; Adams, D.; Alewood, P. Trends in peptide drug discovery. Nat. Rev. Drug Discov. 2021, 20, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Lau, J.L.; Dunn, M.K. Therapeutic peptides: Historical perspectives, current development trends, and future directions. Bioorg. Med. Chem. 2018, 26, 2700–2707. [Google Scholar] [CrossRef] [PubMed]
- Pennington, M.W.; Czerwinski, A.; Norton, R.S. Peptide therapeutics from venom: Current status and potential. Bioorg. Med. Chem. 2018, 26, 2738–2758. [Google Scholar] [CrossRef]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial host defence peptides: Functions and clinical potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef] [PubMed]
- Shaer, A.; Musaimi, O.A.; Albericio, F.; de la Torre, B. 2019 FDA TIDES (Peptides and Oligonucleotides) Harvest. Pharmaceuticals 2020, 13, 40. [Google Scholar] [CrossRef] [Green Version]
- de la Torre, B.G.; Albericio, F. The Pharmaceutical Industry in 2019. An Analysis of FDA Drug Approvals from the Perspective of Molecules. Molecules 2020, 25, 745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ongey, L.; Pflugmacher, S.; Neubauer, P. Bioinspired Designs, Molecular Premise and Tools for Evaluating the Ecological Importance of Antimicrobial Peptides. Pharmaceuticals 2018, 11, 68. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Dong, C.; Li, X.; Han, W.; Su, X. Anticancer potential of bioactive peptides from animal sources (Review). Oncol. Rep. 2017, 38, 637–651. [Google Scholar] [CrossRef] [Green Version]
- Sable, R.; Parajuli, P.; Jois, S. Peptides, Peptidomimetics, and Polypeptides from Marine Sources: A Wealth of Natural Sources for Pharmaceutical Applications. Mar. Drugs 2017, 15, 124. [Google Scholar] [CrossRef] [Green Version]
- Destoumieux-Garzón, D.; Rosa, R.D.; Schmitt, P.; Barreto, C.; Vidal-Dupiol, J.; Mitta, G.; Gueguen, Y.; Bachère, E. Antimicrobial peptides in marine invertebrate health and disease. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150300. [Google Scholar] [CrossRef] [Green Version]
- Guzmán, F.; Wong, G.; Román, T.; Cárdenas, C.; Alvárez, C.; Schmitt, P.; Albericio, F.; Rojas, V. Identification of Antimicrobial Peptides from the Microalgae Tetraselmis suecica (Kylin) Butcher and Bactericidal Activity Improvement. Mar. Drugs 2019, 17, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, B.; Munoz-Garay, C. Marine Antimicrobial Peptides: A Promising Source of New Generation Antibiotics and Other Bio-active Molecules. Int. J. Pept. Res. Ther. 2019, 25, 1441–1450. [Google Scholar] [CrossRef]
- Dorin, J.R.; McHugh, B.J.; Cox, S.L.; Davidson, D.J. Chapter 30—Mammalian Antimicrobial Peptides; Defensins and Cathelicidins. In Molecular Medical Microbiology, 2nd ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 539–565. [Google Scholar]
- Patocka, J.; Nepovimova, E.; Klimova, B.; Wu, Q.; Kuca, K. Antimicrobial Peptides: Amphibian Host Defense Peptides. Curr. Med. Chem. 2019, 26, 5924–5946. [Google Scholar] [CrossRef]
- Shabir, U.; Ali, S.; Magray, A.R.; Ganai, B.A.; Firdous, P.; Hassan, T.; Nazir, R. Fish antimicrobial peptides (AMP’s) as essential and promising molecular therapeutic agents: A review. Microb. Pathog. 2018, 114, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Parisi, K.; Shafee, T.M.A.; Quimbar, P.; Weerden, N.L.v.d.; Bleackley, M.R.; Anderson, M.A. The evolution, function and mechanisms of action for plant defensins. Semin. Cell Dev. Biol. 2018, 88, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Patočka, J.; Kuča, K. Insect Antimicrobial Peptides, a Mini Review. Toxins 2018, 10, 461. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Blencke, H.-M.; Haug, T.; Stensvåg, K. Antimicrobial peptides in echinoderm host defense. Dev. Comp. Immunol. 2015, 49, 190–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Ren, Q. Research progress in innate immunity of freshwater crustaceans. Dev. Comp. Immunol. 2020, 104, 103569. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.J.; Dyrynda, E.A. Antimicrobial proteins: From old proteins, new tricks. Mol. Immunol. 2015, 68, 383–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tassanakajon, A.; Rimphanitchayakit, V.; Visetnan, S.; Amparyup, P.; Somboonwiwat, K.; Charoensapsri, W.; Tang, S. Shrimp humoral responses against pathogens: Antimicrobial peptides and melanization. Dev. Comp. Immunol. 2018, 80, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Tassanakajon, A.; Amparyup, P.; Somboonwiwat, K.; Supungul, P. Cationic Antimicrobial Peptides in Penaeid Shrimp. Mar. Biotechnol. 2010, 13, 487–505. [Google Scholar] [CrossRef] [PubMed]
- Matos, M.; Rosa, R.D. On the silver jubilee of crustacean antimicrobial peptides. Rev. Aquac. 2022, 14, 594–612. [Google Scholar] [CrossRef]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef]
- Rosa, R.D.; Barracco, M.A. Antimicrobial peptides in crustaceans. Invertebr. Surviv. J. 2010, 7, 262–284. [Google Scholar]
- Mogensen, T.H. Pathogen Recognition and Inflammatory Signaling in Innate Immune Defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.-J.; Liu, S.; Liu, H.-P. Recent insights into hematopoiesis in crustaceans. Fish Shellfish Immunol. Rep. 2021, 2, 100040. [Google Scholar] [CrossRef]
- Liu, S.; Zheng, S.-C.; Li, Y.-L.; Li, J.; Liu, H.-P. Hemocyte-Mediated Phagocytosis in Crustaceans. Front. Immunol. 2020, 11, 268. [Google Scholar] [CrossRef] [Green Version]
- Xiao, B.; Fu, Q.; Niu, S.; Li, H.; Lǚ, K.; Wang, S.; Yin, B.; Weng, S.; Li, C.; He, J. Penaeidins are a novel family of antiviral effectors against WSSV in shrimp. BioRxiv 2018, 2018, 467571. [Google Scholar]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agent. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [Green Version]
- Yakovlev, A.; Lysøe, E.; Heldal, I.; Steen, H.; Hagen, S.B.; Clarke, J.L. Transcriptome profiling and in silico detection of the antimicrobial peptides of red king crab Paralithodes camtschaticus. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Schnapp, D.; Kemp, G.D.; Smith, V.J. Purification and characterization of a proline-rich antibacterial peptide, with sequence similarity to bactenecin-7, from the haemocytes of the shore crab, Carcinus maenas. Eur. J. Biochem. 1996, 240, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Khoo, L.; Robinette, D.W.; Noga, E.J. Callinectin, an Antibacterial Peptide from Blue Crab, Callinectes sapidus, Hemocytes. Mar. Biotechnol. 1999, 1, 44–51. [Google Scholar] [CrossRef]
- Noga, J.; Stone, K.L.; Wood, A.; Gordon, W.L.; Robinette, D. Primary structure and cellular localization of callinectin, an antimicrobial peptide from the blue crab. Dev. Comp. Immunol. 2011, 35, 409–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiravanichpaisala, P.; Leea, S.Y.; Kima, Y.-A.; Andren, T.; Söderhäll, I. Antibacterial peptides in hemocytes and hematopoietic tissue from freshwater crayfish Pacifastacus leniusculus: Characterization and expression pattern. Dev. Comp. Immunol. 2007, 31, 441–455. [Google Scholar] [CrossRef]
- Herbinière, J.; Braquart-Varnier, C.; Grève, P.; Strub, J.-M.; Frère, J.; Dorsselaer, A.V.; Martin, G. Armadillidin: A novel glycine-rich antibacterial peptide directed against gram-positive bacteria in the woodlouse Armadillidium vulgare (Terrestrial Isopod, Crustacean). Dev. Comp. Immunol. 2005, 29, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Battison, L.; Summerfield, R.; Patrzykat, A. Isolation and characterisation of two antimicrobial peptides from haemocytes of the American lobster Homarus americanus. Fish Shellfish Immunol. 2008, 25, 181–187. [Google Scholar] [CrossRef]
- Pisuttharachai, D.; Yasuike, M.; Aono, H.; Yano, Y.; Murakami, K.; Kondo, H.; Aoki, T.; Hirono, I. Characterization of two isoforms of Japanese spiny lobster Panulirus japonicus defensin cDNA. Dev. Comp. Immunol. 2009, 33, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Gross, P.S.; Bartlett, T.C.; Browdy, C.L.; Chapman, R.W.; Warr, G.W. Immune gene discovery by expressed sequence tag analysis of hemocytes and hepatopancreas in the Pacific White Shrimp, Litopenaeus vannamei, and the Atlantic White Shrimp, L. setiferus. Dev. Comp. Immunol. 2001, 25, 565–577. [Google Scholar] [CrossRef]
- Matos, M.; Schmitt, P.; Barreto, C.; Farias, N.D.; Toledo-Silva, G.; Guzmán, F.; Destoumieux-Garzón, D.; Perazzolo, L.M.; Rosa, R.D. Massive Gene Expansion and Sequence Diversification Is Associated with Diverse Tissue Distribution, Regulation and Antimicrobial Properties of Anti-Lipopolysaccharide Factors in Shrimp. Mar. Drugs 2018, 16, 381. [Google Scholar] [CrossRef] [Green Version]
- Yedery, R.D.; Reddy, K.V.R. Purification and characterization of antibacterial proteins from granular hemocytes of Indian mud crab, Scylla serrata. Acta Biochim. Pol. 2009, 56, 71–82. [Google Scholar] [CrossRef]
- Barreto, C.; Coelho, J.D.R.; Yuan, J.; Xiang, J.; Perazzolo, L.M.; Rosa, R.D. Specific Molecular Signatures for Type II Crustins in Penaeid Shrimp Uncovered by the Identification of Crustin-Like Antimicrobial Peptides in Litopenaeus vannamei. Mar. Drugs 2018, 16, 31. [Google Scholar] [CrossRef] [Green Version]
- Shan, Z.; Zhu, K.; Peng, H.; Chen, B.; Liu, J.; Chen, F.; Ma, X.; Wang, S.; Qiao, K.; Wang, K. The New Antimicrobial Peptide SpHyastatin from the Mud Crab Scylla paramamosain with Multiple Antimicrobial Mechanisms and High Effect on Bacterial Infection. Front. Microbiol. 2016, 7, 1140. [Google Scholar] [CrossRef] [Green Version]
- Stensvåg, K.; Haug, T.; Sperstad, S.V.; Rekdal, Ø.; Indrevoll, B.; Styrvold, O.B. Arasin 1, a proline–arginine-rich antimicrobial peptide isolated from the spider crab, Hyas Araneus. Dev. Comp. Immunol. 2008, 32, 275–285. [Google Scholar] [CrossRef]
- Ravichandran, G.; Kumaresan, V.; Arasu, M.V.; Al-Dhabi, N.A.; Ganesh, M.-R.; Mahesh, A.; Dhayalan, A.; Pasupuleti, M.; Arockiaraj, J. Pellino-1 derived cationic antimicrobial prawn peptide: Bactericidal activity, toxicity and mode of action. Mol. Immunol. 2016, 78, 171–182. [Google Scholar] [CrossRef]
- Patat, S.A.; Carnegie, R.B.; Kingsbury, C.; Gross, P.S.; Chapman, R.; Schey, K.L. Antimicrobial activity of histones from hemocytes of the Pacific white shrimp. Eur. J. Biochem. 2004, 271, 4825–4833. [Google Scholar] [CrossRef] [PubMed]
- Sruthy, S.; Nair, A.; Antony, S.P.; Puthumana, J.; Singh, I.B.; Philip, R. A histone H2A derived antimicrobial peptide, Fi-Histin from the Indian White shrimp, Fenneropenaeus indicus: Molecular and functional characterization. Fish Shellfish Immunol. 2019, 92, 667–679. [Google Scholar] [CrossRef]
- Destoumieux, D.; Bulet, P.; Strub, J.-M.; Dorsselaer, A.V.; Bachère, E. Recombinant expression and range of activity of penaeidins, antimicrobial peptides from penaeid shrimp. Eur. J. Biochem. 2001, 266, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Lee, B.L.; Soderhall, K. Processing of an Antibacterial Peptide from Hemocyanin of the Freshwater Crayfish Pacifastacus leniusculus. J. Biol. Chem. 2003, 278, 7927–7933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Huang, H.; Wang, F.; Aweya, J.J.; Zheng, Z.; Zhang, Y. Prediction and characterization of a novel hemocyanin-derived antimicrobial peptide from shrimp Litopenaeus vannamei. Amino Acids 2018, 50, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Wan, H.; Zeng, X.; Zhang, Z.; Wang, Y. Characterization and antimicrobial evaluation of a new Spgly-AMP, glycine-rich antimicrobial peptide from the mud crab Scylla paramamosain. Fish Shellfish Immunol. 2020, 106, 384–392. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, F.; Chen, H.-Y.; Peng, H.; Hao, H.; Wang, K.-J. A Novel Antimicrobial Peptide Scyreprocin From Mud Crab Scylla paramamosain Showing Potent Antifungal and Anti-biofilm Activity. Front. Microbiol. 2020, 1589. [Google Scholar] [CrossRef]
- Ravichandran, G.; Kumaresan, V.; Bhatt, P.; Arasu, M.V.; Al-Dhabi, N.A.; Arockiaraj, J. A Cumulative Strategy to Predict and Characterize Antimicrobial Peptides (AMPs) from Protein Database. Int. J. Pept. Res. Ther. 2016, 23, 281–290. [Google Scholar] [CrossRef]
- Farg, A.; Sundaramoorthy, V.; Sultana, J.M.; Yang, S.; Atkinson, R.A.; Levina, V.; Halloran, M.A.; Gleeson, P.A.; Blair, I.P.; Soo, K.Y.; et al. C9ORF72, implicated in amytrophic lateral sclerosis and frontotemporal dementia, regulates endosomal trafficking. Hum. Mol. Genet. 2014, 23, 3579–3595. [Google Scholar] [CrossRef] [PubMed]
- Welch, G.; Li, W.; Hossain, M.A.; Separovic, F.; O’Brien-Simpson, N.M.; Wade, J.D. (Re) Defining the Proline-Rich Antimicrobial Peptide Family and the Identification of Putative New Members. Front. Chem. 2020, 8, 607769. [Google Scholar] [CrossRef]
- Bullard, R.; Sharma, S.R.; Das, P.K.; Morgan, S.E.; Karim, S. Repurposing of Glycine-Rich Proteins in Abiotic and Biotic Stresses in the Lone-Star Tick (Amblyomma americanum). Front. Physiol. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauton, C. The scope of the crustacean immune system for disease control. J. Invertebr. Pathol. 2012, 110, 251–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, P.; Falomir, M.P.; Gozalbo, D. Chitin: A Structural Biopolysaccharide with Multiple Applications; John Wiley & Sons: Hoboken, NJ, USA, 2014; pp. 1–10. [Google Scholar]
- Lu, X.; Lu, H.; Guo, L.; Zhang, Z.; Zhao, X.; Zhong, M.; Li, S.; Zhang, Y. Cloning and Characterization of a Novel Hemocyanin Variant LvHMCV4 from Shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2015, 46, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Petit, V.; Jean-Luc, R.; Alain, B.; Chantal, C.; Chakib, D.; Jean, P.; Christophe, G.; Evelyne, B.; Joëlle, D.; Delphine, D.-G.; et al. A hemocyanin-derived antimicrobial peptide from the penaeid shrimp adopts an alpha-helical structure that specifically permeabilizes fungal membranes. Biochim. Biophys. Acta BBA 2016, 1860, 557–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.; Li, X.; Lu, H.; Lin, R.; Aweya, J.J.; Zhang, Y. N-terminal diversity of Litopenaeus vannamei hemocyanin and immunity. Mol. Immunol. 2019, 112, 360–368. [Google Scholar] [CrossRef]
- Lin, X.; Söderhäll, I. Crustacean hematopoiesis and the astakine cytokines. Blood 2011, 117, 6417–6424. [Google Scholar] [CrossRef] [Green Version]
- Sanfratello, P.M.; Celi, M.; Vazzana, M. Hemocyte types and some plasmatic properties of two edible crabs Cancer borealis and Cancer pagurus. Invertebr. Surviv. J. 2015, 15, 195–202. [Google Scholar]
- Wu, F.; Xie, Z.; Yan, M.; Li, Q.; Song, J.; Hu, M.; Wang, Y. Classification and characterization of hemocytes from two Asian horseshoe crab species Tachypleus tridentatus and Carcinoscorpius rotundicauda. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayasree, S.; Keerthana, C.K.; Anjali Menon, K.; Bhasura, K.; Rahna, S.; Reeha Mashirin, K.; Nidhila, M.; Akshaya, V.; Greeshma, M. Study on the effect of environmental stress on metabolic and immunological parameters in the haemolymph of Fenneropenaeus indicus H. Milne Edwards, 1837. Int. J. Fish. Aquat. Stud. 2017, 5, 490–497. [Google Scholar]
- Day, R.D.; Fitzgibbon, Q.P.; Gardner, C. The impact of holding stressors on the immune function and haemolymph biochemistry of Southern Rock Lobsters (Jasus edwardsii). Fish Shellfish Immunol. 2019, 89, 660–671. [Google Scholar] [CrossRef]
- Braak, B.T.v.d.; Botterblom, M.H.A.; Liu, W.; Taverne, N.; Knaap, W.P.W.v.d.; Rombout, J.H.W.M. The role of the haematopoietic tissue in haemocyte production and maturation in the black tiger shrimp (Penaeus monodon). Fish. Shellfish. Immunol. 2002, 12, 253–272. [Google Scholar] [CrossRef] [PubMed]
- Söderhäll, I. Crustacean hematopoiesis. Dev. Comp. Immunol. 2016, 58, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, M.; Vandenbulcke, F.; Saulnier, D.; Bachère, E. Expression and distribution of penaeidin antimicrobial peptides are regulated by haemocyte reactions in microbial challenged shrimp. Eur. J. Biochem. 2002, 269, 2678–2689. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Soderhall, K. Early events in crustacean innate immunity. Fish Shellfish Immunol. 2002, 12, 421–437. [Google Scholar] [PubMed] [Green Version]
- Cerenius, L.; Lee, B.L.; Soderhall, K. The proPO-system: Pros and cons for its role in invertebrate immunity. Trends Immunol. 2008, 29, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Ponprateep, S.; Vatanavicharn, T.; Lo, C.F.; Tassanakajon, A.; Rimphanitchayakit, V. Alpha-2-macroglobulin is a modulator of prophenoloxidase system in pacific white shrimp Litopenaeus vannamai. Fish Shellfish Immunol. 2017, 62, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Koiwai, K.; Alenton, R.R.R.; Kondo, H.; Hirono, I. Extracellular trap formation in kuruma shrimp (Marsupenaeus japonicus) hemocytes is coupled with c-type lysozyme. Fish Shellfish Immunol. 2016, 52, 206–209. [Google Scholar] [CrossRef]
- Rodríguez, J.; Ruiz, J.; Maldonado, M.; Echeverría, F. Immunodetection of hemocytes, peneidins and α2-macroglobulin in the lymphoid organ of white spot syndrome virus infected shrimp. Microbiol. Immunol. 2012, 56, 562–571. [Google Scholar] [CrossRef]
- Rusaini; Owens, L. Insight into the lymphoid organ of penaeid prawns: A review. Fish Shellfish Immunol. 2010, 29, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Hasson, W.; Lightner, D.V.; Mohney, L.L.; Redman, R.M.; White, B.M. Role of lymphoid organ spheroids in chronic Taura syndrome virus (TSV) infections in Penaeus vannamei. Dis. Aquat. Org. 1999, 38, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Khanobdee, K.; Soowannayan, C.; Flegel, T.W.; Ubol, S.; Withyachumnarnkul, B. Evidence for apoptosis correlated with mortality in the giant black tiger shrimp Penaeus monodon infected with yellow head virus. Dis. Aquat. Org. 2002, 48, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leu, J.H.; Lin, S.J.; Huang, J.Y.; Chen, T.C.; Lo, C.F. A model for apoptotic interaction between white spot syndrome virus and shrimp. Fish Shellfish Immunol. 2013, 34, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Perdomo-Moralesa, R.; Montero-Alejoa, V.; Pererab, E. The clotting system in decapod crustaceans: History, current knowledge and what we need to know beyond the models. Fish Shellfish. Immunol. 2019, 84, 204–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Göpel, T.; Wirkner, C.S. The circulatory system of Penaeus vannamei Boone, 1931—Lacunar function and a reconsideration of the “open vs. closed system” debate. J. Morphol. 2020, 281, 500–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Lv, X.; Yu, Y.; Zhang, X.; Li, F. Molecular and Functional Diversity of Crustin-Like Genes in the Shrimp Litopenaeus vannamei. Mar. Drugs 2020, 18, 361. [Google Scholar] [CrossRef]
- Destoumieux, D.; Munoz, M.; Bulet, P.; Bachère, E. Penaeidins, a family of antimicrobial peptides from penaeid shrimp (Crustacea, Decapoda). Cell. Mol. Life Sci. 2000, 57, 1260–1271. [Google Scholar] [CrossRef]
- Cuthbertson, J.; Yang, Y.; Bachère, E.; Büllesbach, E.E.; Gross, P.S.; Aumelas, A. Solution Structure of Synthetic Penaeidin-4 with Structural and Functional Comparisons with Penaeidin-3. J. Biol. Chem. 2005, 280, 16009–16018. [Google Scholar] [CrossRef] [Green Version]
- An, M.-Y.; Gao, J.; Zhao, X.-F.; Wang, J.-X. A new subfamily of penaeidin with an additional serine-rich region from kuruma shrimp (Marsupenaeus japonicus) contributes to antimicrobial and phagocytic activities. Dev. Comp. Immunol. 2016, 59, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Zhang, C.; Qin, X.; Shi, L.; Zhao, M. Identification and function of penaeidin 3 and penaeidin 5 in Fenneropenaeus merguiensis. Fish Shellfish Immunol. 2019, 89, 623–631. [Google Scholar] [CrossRef]
- Afsal, V.V.; Antony, S.P.; Philip, R.; Singh, I.S.B. Molecular Characterization of a Newly Identified Subfamily Member of Penaeidin from two Penaeid Shrimps, Fenneropenaeus indicus and Metapenaeus monoceros. Probiotics Antimicrob. Proteins 2016, 8, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Tassanakajon, A.; Somboonwiwat, K.; Supungul, P.; Tang, S. Discovery of immune molecules and their crucial functions in shrimp immunity. Fish Shellfish Immunol. 2013, 34, 954–967. [Google Scholar] [CrossRef] [PubMed]
- Destoumieux, D.; Bulet, P.; Loew, D.; Dorsselaer, A.V.; Rodriguez, J.; Bachère, E. Penaeidins, a new family of antimicrobial peptides isolated from the shrimp Penaeus vannamei (Decapoda). J. Biol. Chem. 1997, 272, 28398–28406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, S.; Chao, Y.; Tsao, H.; Sakai, M.; Chou, H.; Song, Y. Molecular cloning and recombinant expression of tiger shrimp Penaeus monodon penaeidin. Fish Pathol. 2004, 39, 15–23. [Google Scholar] [CrossRef]
- Chen, J.-Y.; Pan, C.-Y.; Kuo, C.-M. Molecular cloning and sequencing of shrimp (Penaeus monodon) penaeidin-5 cDNA. Fish Shellfish Immunol. 2004, 16, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.-H.; Song, Y.-L. Cloning of penaeidin gene promoter in tiger shrimp (Penaeus monodon). Fish Shellfish Immunol. 2009, 27, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Imjongjirak, C.; Amphaiphan, P.; Charoensapsri, W.; Amparyup, P. Characterization and antimicrobial evaluation of SpPR-AMP1, a proline-rich antimicrobial peptide from the mud crab Scylla paramamosain. Dev. Comp. Immunol. 2017, 74, 209–216. [Google Scholar] [CrossRef]
- Mishra, K.; Choi, J.; Moon, E.; Baek, K.-H. Tryptophan-Rich and Proline-Rich Antimicrobial Peptides. Molecules 2018, 23, 815. [Google Scholar] [CrossRef] [Green Version]
- Vaseeharan, B.; Shanthi, S.; Chen, J.-C.; Espiñeira, M. Molecular cloning, sequence analysis and expression of Fein-Penaeidin from the haemocytes of Indian white shrimp Fenneropenaeus indicus. Results Immunol. 2012, 2, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Knappe, D.; Schmidt, R.; Adermann, K.; Hoffmann, R. Continuous Subcutaneous Delivery of Proline-Rich Antimicrobial Peptide Api137 Provides Superior Efficacy to Intravenous Administration in a Mouse Infection Model. Front. Microbiol. 2019, 10, 2283. [Google Scholar] [CrossRef] [PubMed]
- Lele, S.; Talat, S.; Kumari, S.; Srivastava, N.; Kaur, K.J. Understanding the importance of glycosylated threonine and stereospecific action of Drosocin, a Proline rich antimicrobial peptide. Eur. J. Med. Chem. 2015, 92, 637–647. [Google Scholar] [CrossRef]
- Cuthbertson, J.; Shepard, E.F.; Chapman, R.W.; Gross, P.S. Diversity of the penaeidin antimicrobial peptides in two shrimp species. Immunogenetics 2002, 54, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Poncet, J.; Garnier, J.; Zatylny, C.; Bachère, E.; Aumelas, A. Solution Structure of the Recombinant Penaeidin-3, a Shrimp Antimicrobial Peptide. J. Biol. Chem. 2003, 278, 36859–36867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barracco, A.; Lorgeril, J.d.; Gueguen, Y.; Bachère, E. Molecular characterization of penaeidins from two Atlantic brazilian shrimp species, Farfantepenaeus paulensis and Litopenaeus schmitti. FEMS Microbiol. Lett. 2005, 250, 117–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Leary, A.; Gross, P.S. Genomic structure and transcriptional regulation of the penaeidin gene family from Litopenaeus vannamei. Gene 2006, 371, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.-J.; Wang, J.-X.; Zhao, X.-F.; Yang, X.-M.; Shao, H.-L.; Xiang, J.-H. Molecular cloning and expression analysis of Ch-penaeidin, an antimicrobial peptide from Chinese shrimp, Fenneropenaeus chinensis. Fish Shellfish Immunol. 2004, 16, 513–525. [Google Scholar] [CrossRef]
- Kang, C.-J.; Xue, J.-F.; Liu, N.; Zhao, X.-F.; Wang, J.-X. Characterization and expression of a new subfamily member of penaeidin antimicrobial peptides (penaeidin 5) from Fenneropenaeus chinensis. Mol. Immunol. 2007, 44, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Fu, Q.; Niu, S.; Zhu, P.; He, J.; Li, C. Penaeidins restrict white spot syndrome virus infection by antagonizing the envelope proteins to block viral entry. Emerg. Microbes Infect. 2019, 9, 390–412. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, G. APD: The Antimicrobial Peptide Database. Nucleic Acids Res. 2004, 32, D590–D592. [Google Scholar] [CrossRef] [Green Version]
- Relf, J.M.; Chisholm, J.R.; Kemp, G.D.; Smith, V.J. Purification and characterization of a cysteine-rich 11.5-kDa antibacterial protein from the granular haemocytes of the shore crab, Carcinus maenas. Eur. J. Biochem. 1999, 264, 350–357. [Google Scholar] [CrossRef] [Green Version]
- Arayamethakorn, S.; Supungul, P.; Tassanakajon, A.; Krusong, K. Characterization of molecular properties and regulatory pathways of CrustinPm1 and CrustinPm7 from the black tiger shrimp Penaeus monodon. Dev. Comp. Immunol. 2017, 67, 18–29. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Lian, Y.-Y.; He, H.-H.; Yuan, K.; Zhang, C.-Z.; Yue, G.H.; He, J.-G. Functional characterization of an ER-stress responding Crustin gene in Litopenaeus vannamei. Fish Shellfish Immunol. 2018, 84, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, T.C.; Cuthbertson, B.J.; Shepard, E.F.; Chapman, R.W.; Gross, P.S.; Warr, G.W. Crustins, homologues of an 11.5-kDa antibacterial peptide, from two species of penaeid shrimp, Litopenaeus vannamei and Litopenaeus setiferus. Mar. Biotechnol 2002, 4, 278–293. [Google Scholar] [CrossRef]
- Rosa, R.D.; Bandeira, P.T.; Barracco, M.A. Molecular cloning of crustins from the hemocytes of Brazilian penaeid shrimps. FEMS Microbiol. Lett. 2007, 274, 287–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tandel, M.; Hipolito, S.G.; Kondo, H.; Hirono, I. Comparative sequence analysis of crustin isoform MjCRS7 and MjWFDC-like gene from kuruma shrimp Marsupenaeus japonicus shows variant of the WFDC domain. Infect. Genet. Evol. 2018, 64, 139–148. [Google Scholar] [CrossRef]
- Vargas-Albores, F.; Martínez-Porchas, M. Crustins are distinctive members of the WAP-containing protein superfamily: An improved classification approach. Dev. Comp. Immunol. 2017, 76, 9–17. [Google Scholar] [CrossRef]
- Smith, V.J.; Fernandes, J.M.O.; Kemp, G.D.; Hauton, C. Crustins: Enigmatic WAP domain-containing antibacterial proteins from crustaceans. Dev. Comp. Immunol. 2008, 32, 758–772. [Google Scholar] [CrossRef] [Green Version]
- Tassanakajon, A.; Somboonwiwat, K.; Amparyup, P. Sequence diversity and evolution of antimicrobial peptides in invertebrates. Dev. Comp. Immunol. 2015, 48, 324–341. [Google Scholar] [CrossRef]
- Liu, N.; Lan, J.-F.; Sun, J.-J.; Jia, W.-M.; Zhao, X.-F.; Wang, J.-X. A novel crustin from Marsupenaeus japonicus promotes hemocyte phagocytosis. Dev. Comp. Immunol. 2015, 49, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Niu, S.; Gao, J.; Zuo, H.; Yuan, J.; Weng, J.H.S.; Xu, X. A single WAP domain (SWD)-containing protein with antiviral activity from Pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2018, 73, 167–174. [Google Scholar] [CrossRef]
- Smith, V.J. Phylogeny of whey acidic protein (WAP) four-disulfide core proteins and their role in lower vertebrates and invertebrates. Biochem. Soc. Trans. 2011, 39, 1403–1408. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.-W.; Man, X.; Wang, Q.-S.S.Y.; Stanley, D.; Hui, K.-M.; Zhang, X.-W. Characterization of a double WAP domain-containing protein from the red swamp crayfish Procambarus clarkia. Fish Shellfish Immunol. 2017, 71, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Visetnan, S.; Supungul, P.; Tassanakajon, A.; Donpudsa, S.; Rimphanitchayakit, V. A single WAP domain-containing protein from Litopenaeus vannamei possesses antiproteinase activity against subtilisin and antimicrobial activity against AHPND-inducing Vibrio parahaemolyticus. Fish Shellfish Immunol. 2017, 68, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Sakunwattana, T.; Jaree, P.; Rimphanitchayakit, V.; Tassanakajon, A.; Tharntada, S. Antibacterial and antiproteinase activities of a double whey acidic protein domain-containing protein from Penaeus vannamei Boone, 1931 (Decapoda, Penaeidae). Crustaceana 2020, 93, 51–69. [Google Scholar] [CrossRef]
- Li, M.; Ma, C.; Li, H.; Peng, J.; Zeng, D.; Chen, X.; Li, C. Molecular cloning, expression, promoter analysis and functional characterization of a new Crustin from Litopenaeus vannamei. Fish Shellfish Immunol. 2018, 73, 42–49. [Google Scholar] [CrossRef]
- Suleiman, S.; Smith, V.J.; Dyrynda, E.A. Unusual tissue distribution of carcinin, an antibacterial crustin, in the crab, Carcinus maenas, reveals its multi-functionality. Dev. Comp. Immunol. 2017, 76, 274–284. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Ma, C.; Zhu, P.; Yang, Y.; Lei, A.; Chen, X.; Liang, W.; Chen, M.; Xiong, J.; Li, C. A new crustin is involved in the innate immune response of shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 94, 398–406. [Google Scholar] [CrossRef]
- Dai, X.; Huang, X.; Zhang, Z.; Zhang, R.; Cao, X.; Zhang, C.; Wang, K.; Ren, Q. Molecular cloning and expression analysis of two type II crustin genes in the oriental river prawn, Macrobrachium nipponense. Fish Shellfish Immunol. 2020, 98, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, P.; Guertler, C.; Bachère, E.; Souza, C.R.B.d.; Rosa, R.D.; Perazzolo, L.M. Molecular signatures at imminent death: Hemocyte gene expression profiling of shrimp succumbing to viral and fungal infections. Dev. Comp. Immunol. 2014, 42, 294–301. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Wang, W.; Ren, Q. Dorsal transcription factor is involved in regulating expression of crustin genes during white spot syndrome virus infection. Dev. Comp. Immunol. 2016, 63, 18–26. [Google Scholar] [CrossRef]
- Liu, N.; Zhang, R.-R.; Fan, Z.-X.; Zhao, X.-F.; Wang, X.-W.; Wang, J.-X. Characterization of a type-I crustin with broad-spectrum antimicrobial activity from red swamp crayfish Procambarus clarkia. Dev. Comp. Immunol. 2016, 61, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Sruthy, K.S.; Nair, A.; Puthumana, J.; Antony, S.P.; Singh, I.S.B.; Philip, R. Molecular cloning, recombinant expression and functional characterization of an antimicrobial peptide, Crustin from the Indian white shrimp, Fenneropenaeus indicus. Fish Shellfish Immunol. 2017, 71, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.-Q.; Li, B.; Shen, X.-L.; Wang, K.; Du, J.; Yu, X.-D.; Yuan, J.-J. A new antimicrobial peptide isoform, Pc-crustin 4 involved in antibacterial innate immune response in fresh water crayfish, Procambarus clarkia. Fish Shellfish Immunol. 2019, 94, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Bandeira, T.; Vernal, J.; Matos, G.M.; Farias, H.T.N.D.; Pinto, A.R.; Barracco, M.A.; Rosa, R.D. A Type IIa crustin from the pink shrimp Farfantepenaeus paulensis (crusFpau) is constitutively synthesized and stored by specific granule-containing hemocyte subpopulations. Fish Shellfish Immunol. 2020, 97, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Rekha, R.; Vaseeharan, B.; Ishwarya, R.; Anjugam, M.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Al-Anbr, M.N.; Govindarajan, M. Searching for crab-borne antimicrobial peptides: Crustin from Portunus pelagicus triggers biofilm inhibition and immune responses of Artemia salina against GFP tagged Vibrio parahaemolyticus Dahv2. Mol. Immunol. 2018, 101, 396–408. [Google Scholar] [CrossRef] [PubMed]
- Sivakamavalli, J.; James, R.A.; Park, K.; Kwak, I.-S.; Vaseeharan, B. Purification of WAP domain-containing antimicrobial peptides from green tiger shrimp Peaneaus semisulcatus. Microb. Pathog. 2020, 140, 103920. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, J.-X.; Wang, Y.; Fang, W.-H.; Wang, Y.; Zhou, J.-F.; Zhao, S.; Li, X.-C. Newly identified type II crustin (SpCrus2) in Scylla paramamosain contains a distinct cysteine distribution pattern exhibiting broad antimicrobial activity. Dev. Comp. Immunol. 2018, 84, 1–13. [Google Scholar] [CrossRef]
- Dai, X.; Wang, K.; Zhang, R.; Zhang, C.; Cao, X.; Huang, X.; Zhang, Y.; Ren, Q. Identification of two carcinin isoforms (MnCarc1 and MnCarc2) and their function in the antimicrobial immunity of Macrobrachium nipponense. Fish Shellfish Immunol. 2020, 106, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Bloa, L.; Boidin-Wichlacz, C.; Cueff-Gauchard, V.; Rosa, R.D.; Cuvillier-Hot, V.; Durand, L.; Methou, P.; Pradillon, F.; Cambon-Bonavita, M.-A.; Tasiemski, A. Antimicrobial Peptides and Ectosymbiotic Relationships: Involvement of a Novel Type IIa Crustin in the Life Cycle of a Deep-Sea Vent Shrimp. Front. Immunol. 2020, 11, 1511. [Google Scholar] [CrossRef]
- Yan, M.C.; Wang, Y.H.; Ji, D.W.; Li, F.; Hu, L.H.; Zhang, M. Expression profiles of three crustin-like genes in Litopenaeus vannamei under Vibrio harveyi infection. Aquacult Int. 2020, 28, 615–624. [Google Scholar] [CrossRef]
- Suthianthong, P.; Pulsook, N.; Supungul, P.; Tassanakajon, A.; Rimphanitchayakit, V. A double WAP domain-containing protein PmDWD from the black tiger shrimp Penaeus monodon is involved in the controlling of proteinase activities in lymphoid organ. Fish Shellfish Immunol. 2011, 30, 783–790. [Google Scholar] [CrossRef]
- Du, Z.Q.; Ren, Q.; Zhao, X.; Wang, J. A double WAP domain (DWD)-containing protein with proteinase inhibitory activity in Chinese white shrimp, Fenneropenaeus chinensis. Comp. Biochem. Physiol. Part B 2009, 154, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Sruthy, K.S.; Chaithanya, E.R.; Sathyan, N.; Nair, A.; Antony, S.P.; Singh, I.S.B.; Philip, R. Molecular Characterization and Phylogenetic Analysis of Novel Isoform of Anti-lipopolysaccharide Factor from the Mantis Shrimp, Miyakea nepa. Probiotics Antimicrob. Proteins 2015, 7, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Nakamura, T.; Morita, T.; Iwanaga, S. Limulus anti-LPS factor: An anticoagulant which inhibits the endotoxin mediated activation of Limulus coagulation system. Biochem. Biophys. Res. Commun. 1982, 105, 717–723. [Google Scholar] [CrossRef]
- Ohashi, K.; Niwa, M.; Nakamura, T.; Morita, T.; Iwanaga, S. Anti-LPS factor in the horseshoe crab, Tachypleus tridentatus. Its hemolytic activity on the red blood cell sensitized with lipopolysaccharide. FEBS Lett. 1984, 176, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Boze, H.; Chemardin, P.; Padilla, A.; Moulin, G.; Tassanakajon, A.; Pugnière, M.; Roquet, F.; Destoumieux-Garzón, D.; Gueguen, Y.; et al. NMR structure of rALF-Pm3, an anti-lipopolysaccharide factor from shrimp: Model of the possible lipid A-binding site. Biopolymers 2008, 91, 207–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tharntada, S.; Somboonwiwat, K.; Rimphanitchayakit, V.; Tassanakajon, A. Anti-lipopolysaccharide factors from the black tiger shrimp, Penaeus monodon, are encoded by two genomic loci. Fish Shellfish Immunol. 2008, 24, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Kamsaeng, P.; Tassanakajon, A.; Somboonwiwat, K. Regulation of antilipopolysaccharide factors, ALFPm3 and ALFPm6, in Penaeus monodon. Sci. Rep. 2017, 7, 12694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, H.-J.; Sun, Q.-L.; Jiang, S.; Zhang, J.; Sun, L. First characterization of an anti-lipopolysaccharide factor (ALF) from hydrothermal vent shrimp: Insights into the immune function of deep-sea crustacean ALF. Dev. Comp. Immunol. 2018, 84, 382–395. [Google Scholar] [CrossRef]
- Zhou, L.; Li, G.; Li, A.; Jiao, Y.; Li, S.; Huang, J.; Yang, L.; Wang, C. Characterization of a group D anti-lipopolysaccharide factor (ALF) involved in anti-Vibrio response in Penaeus monodon. Fish Shellfish Immunol. 2019, 89, 384–392. [Google Scholar] [CrossRef]
- Zhou, L.; Li, G.; Jiao, Y.; Huang, D.; Li, A.; Chen, H.; Liu, Y.; Li, S.; Li, H.; Wang, C. Molecular and antimicrobial characterization of a group G anti-lipopolysaccharide factor (ALF) from Penaeus monodon. Fish Shellfish Immunol. 2019, 94, 149–156. [Google Scholar] [CrossRef]
- Sruthy, K.S.; Nair, A.; Puthumana, J.; Singh, B.; Philip, R. Molecular cloning, recombinant expression and functional characterization of a novel isoform of anti-lipopolysaccharide factor from the crucifix crab, Charybdis feriatus. Fish Shellfish Immunol. 2016, 53, 116. [Google Scholar]
- Li, S.; Lv, X.; Li, F.; Xiang, J. Characterization of a Lymphoid Organ Specific Anti-lipopolysaccharide Factor From Shrimp Reveals Structure-Activity Relationship of the LPS-Binding Domain. Front. Immunol. 2019, 10, 872. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Li, S.; Liu, F.; Li, F.; Xiang, J. Identification and function analysis of an anti-lipopolysaccharide factor from the ridgetail prawn Exopalaemon carinicauda. Dev. Comp. Immunol. 2017, 70, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Srisapoome, P.; Klongklaew, N.; Areechon, N.; Wongpanya, R. Molecular and functional analyses of novel anti-lipopolysaccharide factors in giant river prawn (Macrobrachium rosenbergii, De Man) and their expression responses under pathogen and temperature exposure. Fish Shellfish Immunol. 2018, 80, 357–375. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.-Y.; Gao, Y.; Wang, H.; Zhang, Q.-X.; Zeng, C.-L.; Liu, H.-P. Identification of an anti-lipopolysacchride factor possessing both antiviral and antibacterial activity from the red claw crayfish Cherax quadricarinatus. Fish Shellfish Immunol. 2016, 57, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Nam, B.H.; Park, E.H.; Shin, E.H.; Kim, Y.O.; Kim, D.G.; Kong, H.J.; Park, J.Y.; Seo, J.K. Development of novel antimicrobial peptides derived from anti-lipopolysaccharide factor of the swimming crab, Portunus trituberculatus. Fish Shellfish Immunol. 2019, 84, 664–672. [Google Scholar] [CrossRef]
- Li, S.; Zhang, X.; Sun, Z.; Li, F.; Xiang, J. Transcriptome Analysis on Chinese Shrimp Fenneropenaeus chinensis during WSSV Acute Infection. PLoS ONE 2013, 8, e58627. [Google Scholar] [CrossRef] [Green Version]
- Supungul, P.; Klinbunga, S.; Pichyangkura, R.; Jitrapakdee, S.; Hirono, I.; Aoki, T.; Tassanakajon, A. Identification of immune-related genes in hemocytes of black tiger shrimp (Penaeus monodon). Mar. Biotechnol. 2002, 4, 487–494. [Google Scholar] [CrossRef]
- Somboonwiwat, K.; Marcos, M.; Tassanakajon, A.; Klinbunga, S.; Aumelas, A.; Romestand, B.; Gueguen, Y.; Boze, H.; Moulin, G.; Bachère, E. Recombinant expression and anti-microbial activity of anti-lipopolysaccharide factor (ALF) from the black tiger shrimp Penaeus monodon. Dev. Comp. Immunol. 2005, 29, 841–851. [Google Scholar] [CrossRef]
- Mekata, T.; Sudhakaran, R.; Okugawa, S.; Kono, T.; Sakai, M.; Itami, T. Molecular cloning and transcriptional analysis of a newly identified anti-lipopolysaccharide factor gene in kuruma shrimp, Marsupenaeus japonicus. Lett. Appl. Microbiol. 2010, 50, 112–119. [Google Scholar] [CrossRef]
- Zhang, H.; Cheng, W.; Zheng, L.; Wang, P.; Liu, Q.; Li, Z.; Li, T.; Wei, Y.; Mao, Y.; Yu, X. Identification of a group D anti-lipopolysaccharide factor (ALF) from kuruma prawn (Marsupenaeus japonicus) with antibacterial activity against Vibrio parahaemolyticus. Fish Shellfish Immunol. 2020, 102, 368–380. [Google Scholar] [CrossRef]
- Jiang, H.-S.; Zhang, Q.; Zhao, Y.-R.; Jia, W.-M.; Zhao, X.-F.; Wang, J.-X. A new group of anti-lipopolysaccharide factors from Marsupenaeus japonicus functions in antibacterial response. Dev. Comp. Immunol. 2015, 48, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhou, L.; Guoqiang, L.; Zhangli, H. A kind of Penaeus monodon Cecropin A LFpm10 and Preparation Method Thereof. Chinese Patent CN109021088A, 10 September 2021. [Google Scholar]
- Farias, N.D.; Falchetti, M.; Matos, G.M.; Schmitt, P.; Barreto, C.; Argenta, N.; Rolland, J.-L.; Bachère, E.; Perazzolo, L.M.; Rosa, R.D. Litopenaeus vannamei stylicins are constitutively produced by hemocytes and intestinal cells and are differentially modulated upon infections. Fish Shellfish Immunol. 2019, 86, 82–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soo, C.C.; Devadas, S.; Din, M.S.M.; Bhassu, S. Differential transcriptome analysis of the disease tolerant Madagascar–Malaysia crossbred black tiger shrimp, Penaeus monodon hepatopancreas in response to acute hepatopancreatic necrosis disease (AHPND) infection: Inference on immune gene response and interaction. Gut Pathog. 2019, 11, 1–13. [Google Scholar]
- Edwards, A.; Elliott, A.G.; Kavanagh, A.M.; Zuegg, J.; Blaskovich, M.A.T.; Cooper, M.A. Contribution of Amphipathicity and Hydrophobicity to the Antimicrobial Activity and Cytotoxicity of β-Hairpin Peptides. ACS Infect. Dis. 2016, 2, 442–450. [Google Scholar] [CrossRef]
- Lee, B.-H.; Chen, B.-R.; Huang, C.-T.; Lin, C.-H. The Immune Activity of PT-Peptide Derived from Anti-Lipopolysaccharide Factor of the Swimming Crab Portunus trituberculatus Is Enhanced when Encapsulated in Milk-Derived Extracellular Vesicles. Mar. Drugs 2019, 17, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.C.; Lin, S.B.; Lee, S.C.; Lin, C.C.; Hui, C.F.; Chen, J.Y. Antimicrobial peptide of an anti-lipopolysaccharide factor modulates of the inflammatory response in RAW264.7 cells. Peptides 2010, 31, 1262–1272. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-C.; Lin, S.-B.; Chen, J.-C.; Hui, C.-F.; Chen, J.-Y. Shrimp anti-lipopolysaccharide factor peptide enhances the antitumor activity of cisplatin in vitro and inhibits HeLa cells growth in nude mice. Peptides 2010, 31, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, J.; Büllesbach, E.E.; Gross, P.S. Discovery of synthetic penaeidin activity against antibiotic-resistant fungi. Chem. Biol. Drug Des. 2006, 68, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Sruthy, S.; Philip, R. Anti-lipopolysaccharide Factor from Crucifix Crab Charybdis feriatus, Cf-ALF2: Molecular Cloning and Functional Characterization of the Recombinant Peptide. Probiotics Antimicrob. Proteins 2021, 13, 885–898. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Xie, X.Y.; Chen, K.; Chen, X.L.; Zhu, W.L.; Wang, H.L. Immune Responses to Gram-Negative Bacteria in Hemolymph of the Chinese Horseshoe Crab, Tachypleus tridentatus. Font. Inmunol. 2020, 11, 584808. [Google Scholar] [CrossRef] [PubMed]
- Ariki, S.; Koori, K.; Osaki, T.; Motoyama, K.; Inamori, K.; Kawabata, S. A serine protease zymogen functions as a pattern-recognition receptor for lipopolysaccharides. Proc. Natl. Acad. Sci. USA 2004, 101, 953–958. [Google Scholar] [CrossRef] [Green Version]
- Iwanaga, S.; Lee, B.L. Recent Advances in the Innate Immunity of Invertebrate Animals. J. Biochem. Mol. Biol. 2005, 38, 128–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanjani, N.T.; Miranda-Saksena, M.; Cunningham, A.L.; Dehghani, F. Antimicrobial Peptides of Marine Crustaceans: The Potential and Challenges of Developing Therapeutic Agents. Curr. Med. Chem. 2018, 25, 2245–2259. [Google Scholar] [CrossRef] [PubMed]
- Muta, T.; Fujimoto, T.; Nakajima, H.; Iwanaga, S. Tachyplesins Isolated from Hemocytes of Southeast Asian Horseshoe Crabs (Carcinoscorpius rotundicauda and Tachypleus gigas): Identification of a New Tachyplesin, Tachyplesin III, and a Processing Intermediate of Its Precursor. J. Biochem. 1990, 108, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Miyata, T.; Tokunaga, F.; Yoneya, T.; Yoshikawa, K.; Iwanaga, S.; Niwa, M.; Takao, T.; Shimonishi, Y. Antimicrobial peptides, isolated from horseshoe crab hemocytes, tachyplesin II, and polyphemusins I and II: Chemical structures and biological activity. J. Biochem. 1989, 106, 663–668. [Google Scholar] [CrossRef]
- Morimoto, M.; Mori, H.; Otake, T.; Ueba, N.; Kunita, N.; Niwa, M.; Murakami, T.; Iwanaga, S. Inhibitory effect of tachyplesin I on the proliferation of human immunodeficiency virus in vitro. Chemotherapy 1991, 37, 206–211. [Google Scholar] [CrossRef]
- Tincu, J.A.; Taylor, S.W. Antimicrobial Peptides from Marine Invertebrates. ASM J./Antimicrob. Agents Chemother. 2004, 48, 3645–3654. [Google Scholar] [CrossRef] [Green Version]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Zhang, L.; Scott, M.G.; Yan, H.; Mayer, L.D.; Hancock, R.E. Interaction of polyphemusin I and structural analogs with bacterial membranes, lipopolysaccharide, and lipid monolayers. Biochemistry 2000, 39, 14504–14514. [Google Scholar] [CrossRef]
- Bulet, P.; Stöcklin, R.; Menin, L. Anti-microbial peptides: From invertebrates to vertebrates. Immunol. Rev. 2004, 198, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, S.I.; Nagayama, R.; Hirata, M.; Shigenaga, T.; Agarwala, K.L.; Saito, T.; Cho, J.; Nakajima, H.; Takagi, T.; Iwanaga, S. Tachycitin, a small granular component in horseshoe crab hemocytes, is an antimicrobial protein with chitin-binding activity. J. Biochem. 1996, 120, 1253–1260. [Google Scholar] [CrossRef]
- Kouno, T.; Fujitani, N.; Mizuguchi, M.; Osaki, T.; Nishimura, S.I.; Kawabata, S.I.; Aizawa, T.; Demura, M.; Nitta, K.; Kawano, K. A Novel β-Defensin Structure: A Potential Strategy of Big Defensin for Overcoming Resistance by Gram-Positive Bacteria. Biochemistry 2008, 47, 10611–10619. [Google Scholar] [CrossRef] [PubMed]
- Suetake, T.; Tsuda, S.; Kawabata, S.; Miura, K.; Iwanaga, S.; Hikichi, K.; Nitta, K.; Kawano, K. Chitin-binding proteins in invertebrates and plants comprise a common chitin-binding structural motif. J. Biol. Chem. 2000, 275, 17929–17932. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, C.; Sarkar, P.; Issa, R.; Haldar, J. Alternatives to Conventional Antibiotics in the Era of Antimicrobial Resistance. Trends Microbiol. 2018, 27, 323–338. [Google Scholar] [CrossRef]
- Behrendt, R.; White, P.; Offer, J. Advances in Fmoc solid-phase peptide synthesis. J. Pept. Sci. 2016, 22, 4–27. [Google Scholar] [CrossRef] [Green Version]
- Cuthbertson, B.; Bullesbach, E.; Fievet, J.; Bachere, E.; Gross, P. A new class (penaeidin class 4) of antimicrobial peptides from the Atlantic white shrimp (L. setiferus) exhibits target specificity and an independent proline-rich-domain function. Biochem. J. 2004, 381, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agouridas, V.; El Mahdi, O.; Diemer, V.; Cargoët, M.; Monbaliu, J.C.M.; Melnyk, O. Native Chemical Ligation and Extended Methods: Mechanisms, Catalysis, Scope, and Limitations. Chem. Rev. 2019, 119, 7328–7443. [Google Scholar] [CrossRef] [PubMed]












| AMP | Characteristics | Sequence Affinity with Other Animal Taxa | Order | Family | Species | References |
|---|---|---|---|---|---|---|
| Bac-like | 6.5 kDa, Pro-rich, cationic | Fifth iteration, mostly Pro-rich bacterial peptides of unknown functions * | Decapoda | Carcinidae | Carcinus maenas | [38] |
| Callinectin | 3.7 kDa, Pro/Arg-rich, similar to arasins, cationic | No sequence similarity detected * | Decapoda | Portunidae | Callinectes sapidus | [39,40] |
| Astacidin-2 | 1.8 kDa, Pro/Arg-rich, cationic | Similar to arasin and arasin-like proteins from Scylla spp., Hyas Araneus * | Decapoda | Astacidae | Pacifastacus leniusculus | [41] |
| Armadillidin | 5.3 kDa, Gly-rich, cationic | No sequence similarity detected * | Isopoda | Armadillidiidae | Armadillidium vulgare | [42] |
| Homarin (CAP-1) | 4–6 kDa, putatively amphipathic α-helical, cationic | Similar to amphibian temporins | Decapoda | Nephropidae | Homarus americanus | [43] |
| Defensins | 6.8–7.2 kDa, contains three disulfide bonds, cationic | Shares six putatively homologous Cys residues with mammalian β-defensins | Decapoda | Palinuridae | Panulirus japonicus | [44] |
| Anti LPS factor | 7–11 kDa, contains a highly hydrophobic N-terminal region and the two Cys residues. The 3D structure of shrimp ALF consists of three α-helices and four-stranded β-sheets; the functional LPS-binding domain is a conserved cluster of positively charged residues within a β-hairpin located between two conserved Cys residues | Widely distributed in Crustacea * | Decapoda | Penaeidae | Metapenaeus dobsoni, Penaeus spp. (9) | [24,45,46] |
| Atyidae | Neocaridina heteropoda | |||||
| Palaemonidae | Macrobrachium spp. (3), Palaemon carinicauda | |||||
| Nephropidae | Homarus americanus | |||||
| Parastacidae | Cherax quadricarinatus | |||||
| Astacidae | Pacifastacus leniusculus | |||||
| Cambaridae | Procambarus clarkii | |||||
| Varunidae | Eriocheir sinensis | |||||
| Oregoniidae | Cionoecetes opilio | |||||
| Portunidae | Charybdis spp. (2), Portunus spp. (2), Scylla spp. (4) | |||||
| Amphipoda | Hyaelellidae | Hyalella azteca | ||||
| Talitridae | Trinorchestia longiramus | |||||
| Isopoda | Armadillidiidae | Armadillidium spp. (2) | ||||
| Scygonadin | 10.8 kDa, contains two Cys residues reminiscent of ALFs, anionic | Found only in Scylla serrata seminal fluid; shows similarities to ALFs, which might indicate a common evolutionary origin | Decapoda | Astacidae | Scylla serrata | [14] |
| Scylla serrata antimicrobial protein (SSAP) | 11.4 kDa, anionic | A Scygonadin homolog | Decapoda | Astacidae | Scylla serrata | [47] |
| Penaeidin | 4.7–7.2 kDa, cationic, N-terminal Pro/Arg-rich domain, C-terminal domain contains an amphipathic helix and two coils constrained by three disulfide bonds | Contains at least four subgroups found only in penaeidae shrimp | Decapoda | Penaeidae | Penaeus spp. (12) Penaeus monoceros | [16] |
| Crustin | 6–22 kDa, cationic, contains whey acidic protein (WAP) domain, further characterized by Gly-rich regions and conserved Cys residues | Found throughout Crustacea, and even in some hymenopterans * | Anostraca | Artemiidae | Artemia salina | [16,48] |
| Decapoda | Penaeidae | Penaeus spp. (6) | ||||
| Atyidae | Neocaridina heteropoda | |||||
| Palaemonidae | Macrobrachium nipponense | |||||
| Pandalidae | Pandalus japonicus | |||||
| Alvinocarididae | Rimicaris exoculata | |||||
| Palinuridae | Panulirus spp. (2) | |||||
| Nephropidae | Homarus spp. (2) | |||||
| Parastasidae | Cherax quadricarinatus | |||||
| Astacidae | Pacifastacus leniusculus | |||||
| Lithodidae | Paralithodes camtschaticus | |||||
| Oregoniidae | Hyas araneus | |||||
| Portunidae | Portunus pelagicus Scylla spp. (3) | |||||
| Hyastatin | 11.7 kDa, cationic, contains a Gly-rich region, Pro-rich domain, and Cys-rich domain | High sequence similarity with SpHyastatin | Decapoda | Oregoniidae | Hyas araneus Cionoecetes opilio | [49] |
| Portunidae | Portunus trituberculatus | |||||
| SpHyastatin | 14.1 kDa, cationic, contains Pro-rich domain and Cys-rich domain | High sequence similarity with Hyastatin, but lacking a Gly-rich region | Decapoda | Portunidae | Scylla paramamosain | [49] |
| Arasin | 4.3–4.8 kDa, cationic, Pro- and Arg-rich | Similar to Astacidin-2 | Decapoda | Oregoniidae | Hyas araneus | [50] |
| Stylicin | 8.9 kDa, multidomain, N-terminal Pro/Arg-rich domain, C-terminal domain contains 13 Cys residues | Found only in Penaeidae. | Decapoda | Penaeidae | Penaeus spp. (4) Penaeus japonicus | [16] |
| Pellino-1-derived cationic antimicrobial prawn peptide | 8.0 kDa, β-sheet forming, cationic | Artificial, based on bioinformatic analyses of the structure of Pellino-1. | Decapoda | Palaemonidae | Macrobrachium rosenbergii | [51] |
| Histones or histone-derived peptides | Fi-Histidin: 2.9 kDa, enriched in Arg, Ala, Gly, Leu, Ser, cationic, α-helical structure H2A: 13.2 kDa, cationic H2B: 13.5 kDa, cationic | Highly conserved with great sequence similarity; antimicrobial action might be due to sequence at the N-terminus * | Decapoda | Penaeidae | Penaeus spp. (2) | [52,53] |
| Hemocyanin-derived peptides | C-terminus: 7.9–8.3 kDa, anionic, His-rich, α-helical structure Astacin-1: 1.9 kDa, cationic Predicted AMPs: 1.5–1.8 kDa | Highly conserved with great sequence similarity * | Decapoda | Penaeidae | Penaeus spp. (2) | [54,55,56] |
| Astacidae | Pacifastacus leniusculus | |||||
| Cambaridae | Procambarus clarkii | |||||
| Spgly-amp | 3.98 kDa, cationic, Gly-rich | Known from a single species with a hypothetical protein detected in Portunus trituberculatus, no sequence similarities detected in other Crustacean families *. | Decapoda | Portunidae | Scylla paramamosain | [57] |
| Scyreprocin | 9.1 kDa, cationic, Lys, Ala and Ser are the most common amino acids | Known from a single species with a hypothetical protein detected in Portunus trituberculatus, no sequence similarities detected in other Crustacean families *. | Decapoda | Portunidae | Scylla paramamosain | [58] |
| Penaeidin Class | Peptide Name/ Accession GB | Shrimp Species | Function/ Activity | Structure, pI, MW | Reference |
| PEN 1/2 | Lv2a (Y14925) | Litopenaeus vannamei | Anti-Gram positiveAntifungal | [94] | |
| Lv2b (AF390146) | Litopenaeus vannamei | Not determined | [103] | ||
| Ls2d (AY039205) | Litopenaeus setiferus | Not determined | [103] | ||
| PEN 3 | MmPEN (KF275674) | Metapenaeus monoceros |  | [92] | |
| FiPEN (JX657680) | Fenneropenaeus indicus | Anti-Gram negative | [92] | ||
| FmPEN3 (MK092338) | Fenneropenaeus merguiensis | Anti-Gram negative | [91] | ||
| PEN 4 | PEN4-1 | Litopenaeus schmitti |  | [105] | |
| PEN-4c (AF390149) | Litopenaeus vannamei | [103] | |||
| Penaeidin 4 (QHD40386) | Penaeus pulensis | Unpublished results | |||
| PEN 5 | FmPEN5 (MK092339) | Fenneropenaeus merguiensis | Anti-Gram negative moderate |  | [91] |
| PEN5 (FJ686018) | Penaeus monodon | Anti-Gram negative Anti-Gram positive Anti Fungi | [28] | ||
| PEN Family II | MjPen-II (AMH87234) | Marsupenaeus japonicus | Gram negative Gram positive Enhances agglutination |  | [35] |
| Crustin Type | Peptide Name/ Accession GB | Shrimp Species | Function/ Activity | Structure, Ip, MW | Reference |
|---|---|---|---|---|---|
| Crustin type Ia | MjCruI-1 | Penaeus japonicus | Anti-Gram negative Anti-Gram positive Phagocytosis-promoting activities |  | [120,139] |
| Crustin type 1a-1 MT375557 | Penaeus vannamei | Not determined | [87] | ||
| Crustin type 1a-2 MT375558 | Penaeus vannamei | Not determined | [87] | ||
| Crustin type Ib | FcCru Ib-1 MT375591 | Penaeus chinensis | Not determined |  | [87] |
| FcCru Ib-2 MT375592 | Penaeus chinensis | Not determined | [87] | ||
| LvCru Ib-1 (MT375567) | Penaeus vannamei | Not determined | [87] | ||
| Crustin type Ic | FcCru Ic (MT375593) | Penaeus chinensis | Not determined | [87] | |
| LvCru Ic (MT375569) | Penaeus vannamei | Not determined | [87] | ||
| Crustin type IIa | crusFpau (EF182747) | Penaeus paulensis | Anti-Gram negative Anti-Gram positive |  | [135] |
| LvCrustinB | Penaeus vannamei | Anti-Gram negative (low) Anti-WSSV (low) | [128] | ||
| ReCrustin (FJ573157) | Rimicaris exoculata | Anti-Gram positive | [140] | ||
| Crustin type IIb | Crustin-like Lv (JQ824114) | Penaeus vannamei | Discarded |  | [48] |
| LvCru IIb-1 (MT375581) | Penaeus vannamei | Not determined | [87] | ||
| LvCru IIb-2 (MT375582) | Penaeus vannamei | Not determined | [87] | ||
| LvCru IIb-3 (MT375583) | Penaeus vannamei | Not determined | [87] | ||
| Crustin type III | LvSWD3 | Penaeus vannamei | Antiviral(WSSV) |  | [121] |
| Cru III-3 (MT375586) | Penaeus vannamei | Not determined | [87] | ||
| Cru III-4 (MT375587) | Penaeus vannamei | Not determined | [87] | ||
| Crustin A (ARB15844) | Penaeus vannamei | Anti-Gram negative | [141] | ||
| Crustin type IV | PvDWD1 (ABR19819) | Penaeus vannamei | Anti-Gram positive, anti-proteinase |  | [125] |
| PmDWD (BI784457) | Penaeu monodon | Anti-proteinase (moderate) | [142] | ||
| FcDWD | Penaeu chinensis | Anti-Gram positive Anti-Gram negative Anti-proteinase | [143] | ||
| Crustin type VI | FcCruVI (MT375594) | Penaeus chinensis | Not determined |  | [87] |
| LvCruVI (MT375589) | Penaeus vannamei | Not determined | [87] | ||
| Crustin type VII | LvCruVII (MT375590) | Penaeus vannamei | Not determined |  | [87] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saucedo-Vázquez, J.P.; Gushque, F.; Vispo, N.S.; Rodriguez, J.; Gudiño-Gomezjurado, M.E.; Albericio, F.; Tellkamp, M.P.; Alexis, F. Marine Arthropods as a Source of Antimicrobial Peptides. Mar. Drugs 2022, 20, 501. https://doi.org/10.3390/md20080501
Saucedo-Vázquez JP, Gushque F, Vispo NS, Rodriguez J, Gudiño-Gomezjurado ME, Albericio F, Tellkamp MP, Alexis F. Marine Arthropods as a Source of Antimicrobial Peptides. Marine Drugs. 2022; 20(8):501. https://doi.org/10.3390/md20080501
Chicago/Turabian StyleSaucedo-Vázquez, Juan Pablo, Fernando Gushque, Nelson Santiago Vispo, Jenny Rodriguez, Marco Esteban Gudiño-Gomezjurado, Fernando Albericio, Markus P. Tellkamp, and Frank Alexis. 2022. "Marine Arthropods as a Source of Antimicrobial Peptides" Marine Drugs 20, no. 8: 501. https://doi.org/10.3390/md20080501
APA StyleSaucedo-Vázquez, J. P., Gushque, F., Vispo, N. S., Rodriguez, J., Gudiño-Gomezjurado, M. E., Albericio, F., Tellkamp, M. P., & Alexis, F. (2022). Marine Arthropods as a Source of Antimicrobial Peptides. Marine Drugs, 20(8), 501. https://doi.org/10.3390/md20080501