
Gut and Gill-Associated Microbiota of the Flatfish European Plaice (Pleuronectes platessa): Diversity, Metabolome and Bioactivity against Human and Aquaculture Pathogens


Abstract
:1. Introduction
2. Results
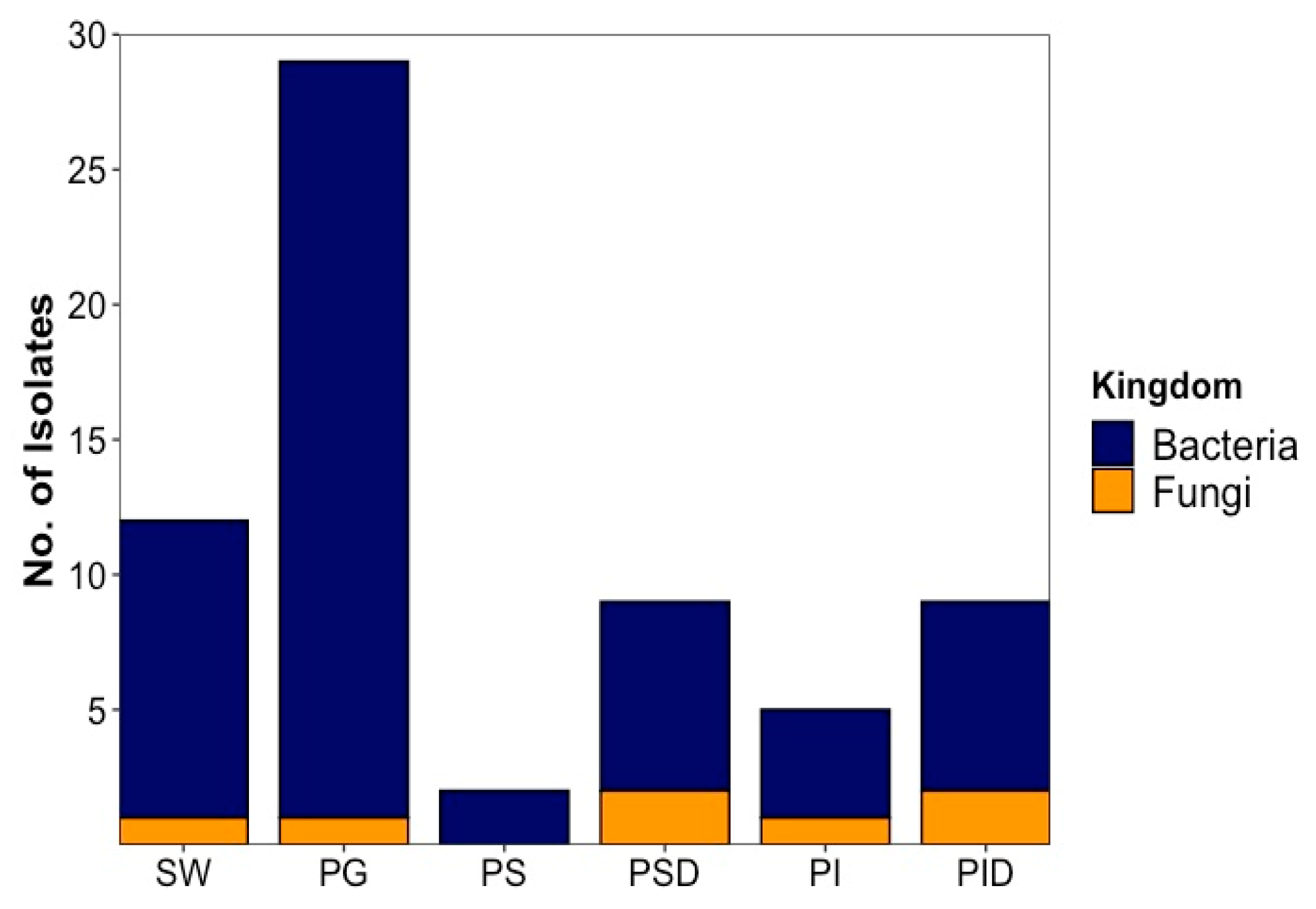
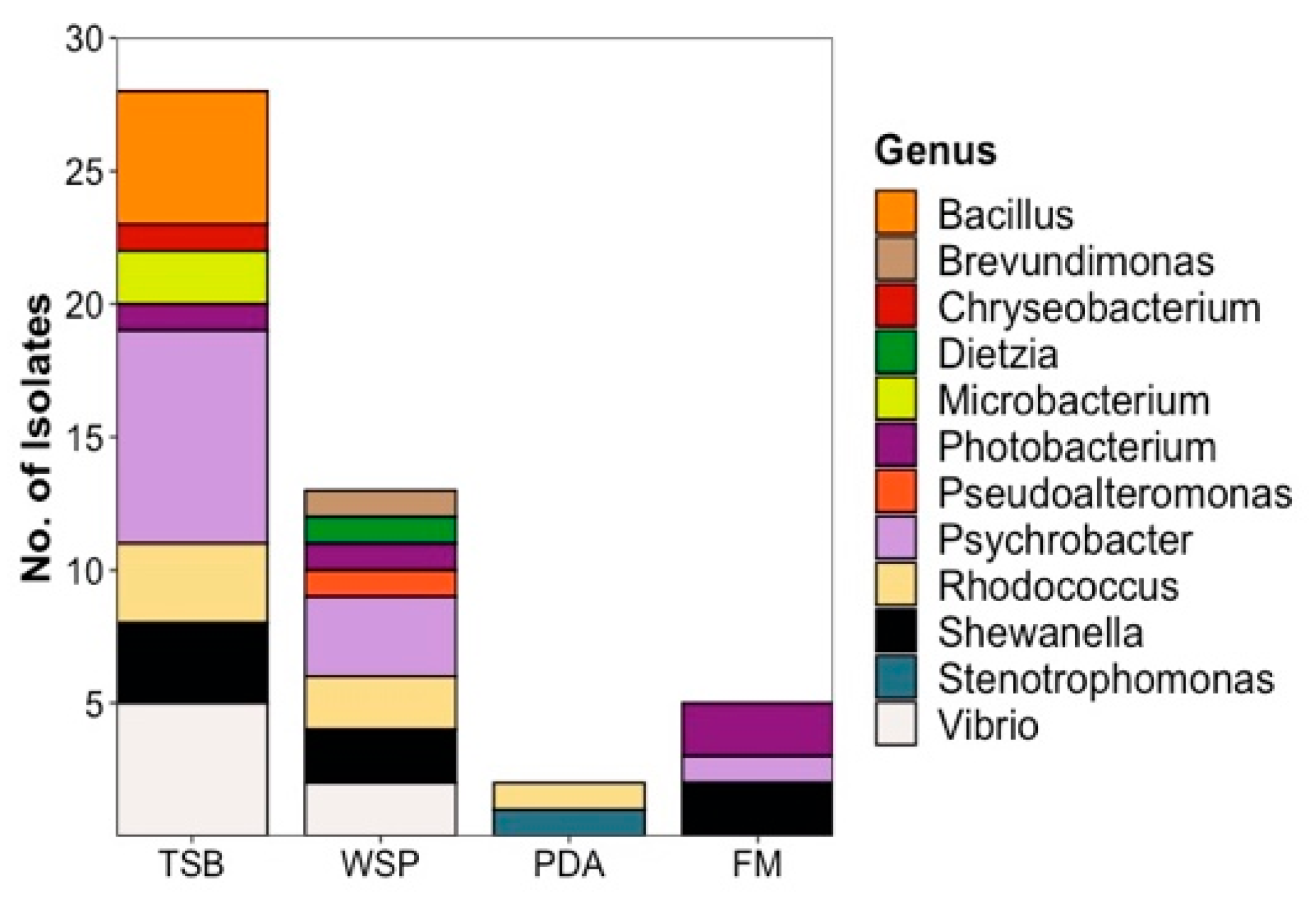
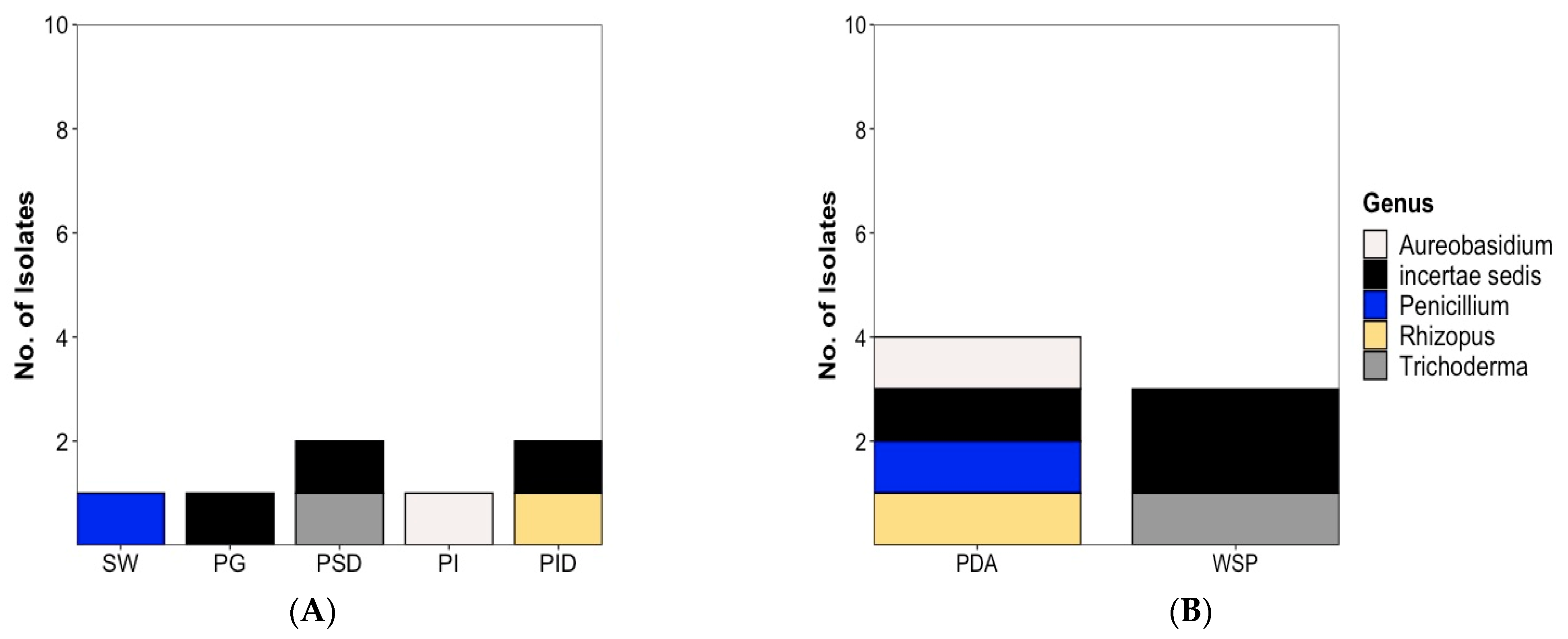
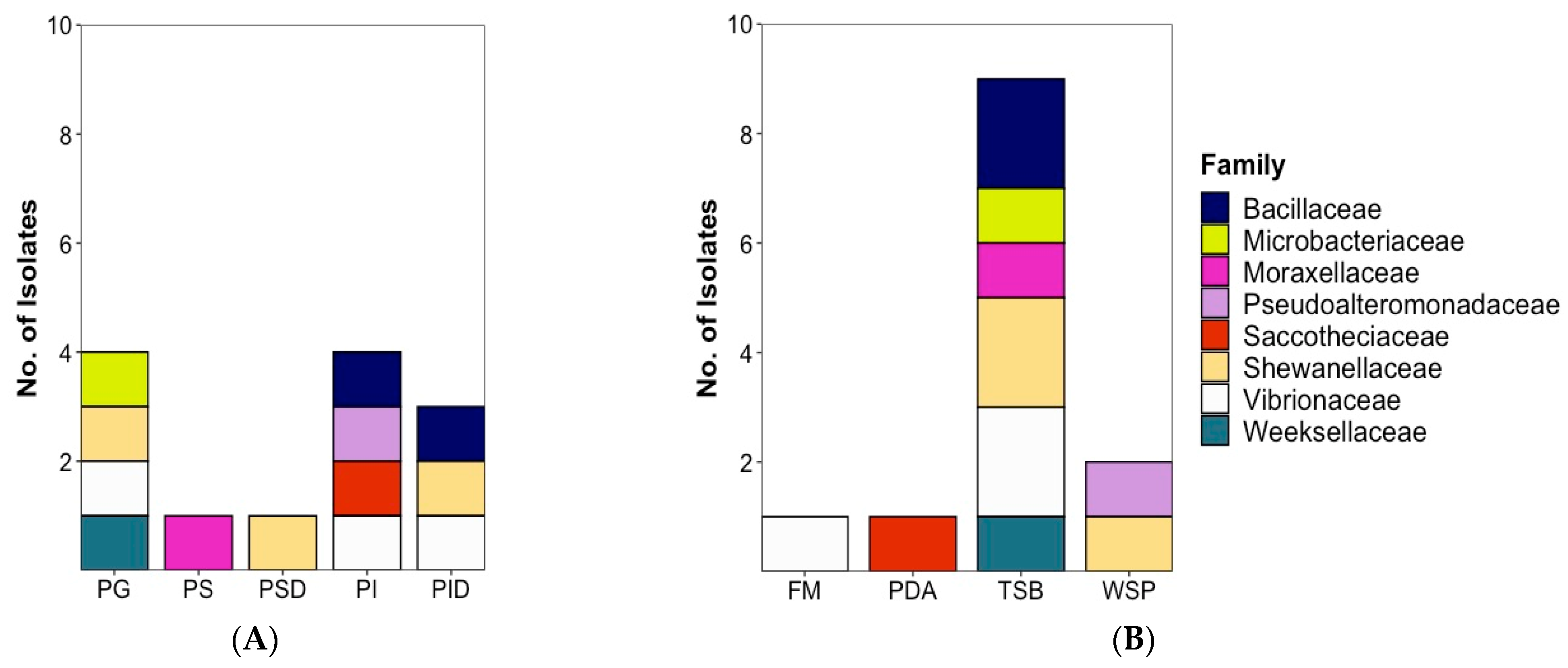
2.1. Strain Isolation and Cultivation
2.1.1. Bacterial Isolates
2.1.2. Fungal Isolates

2.2. Cultivation, Extraction and Bioactivity Assessments
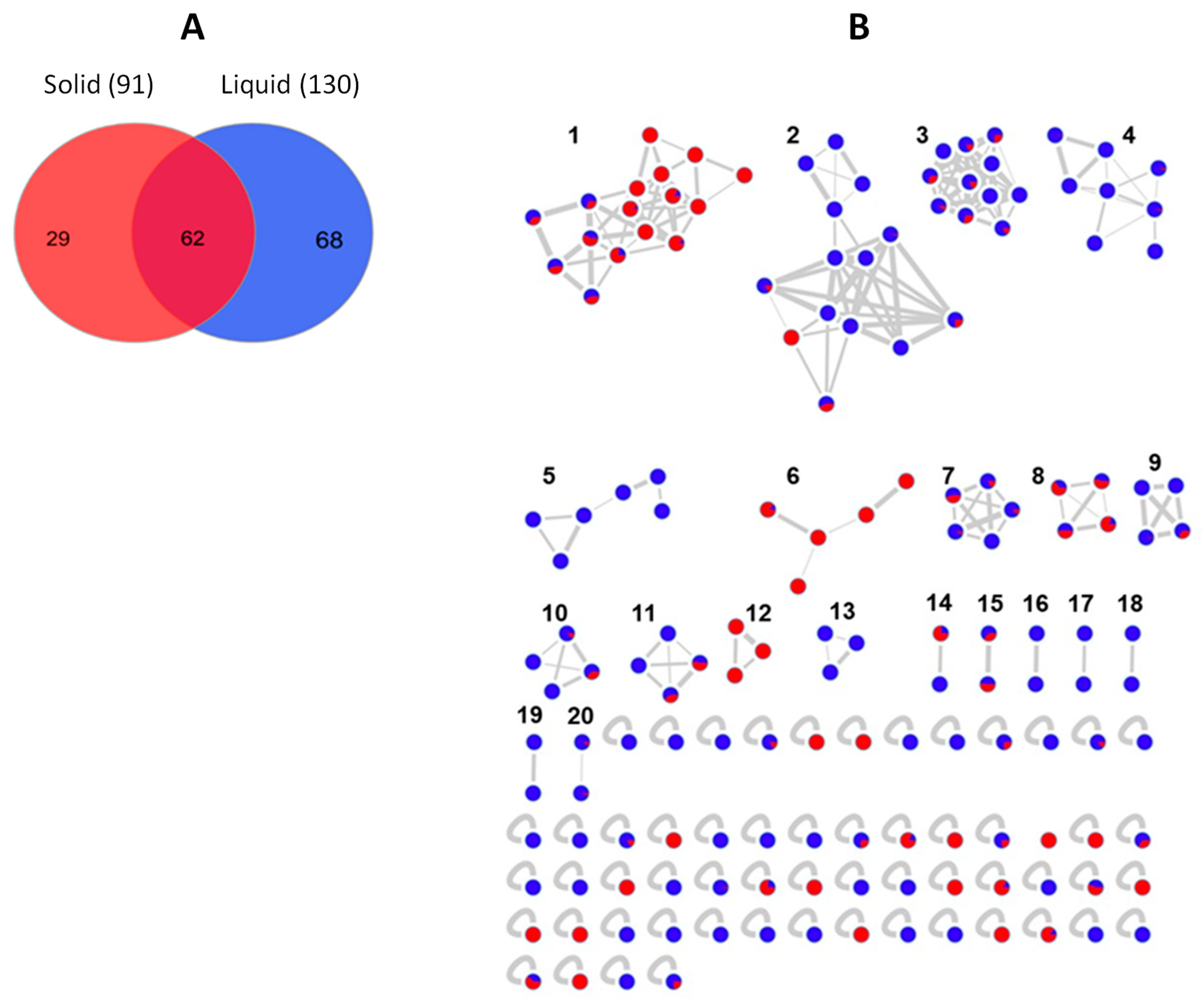
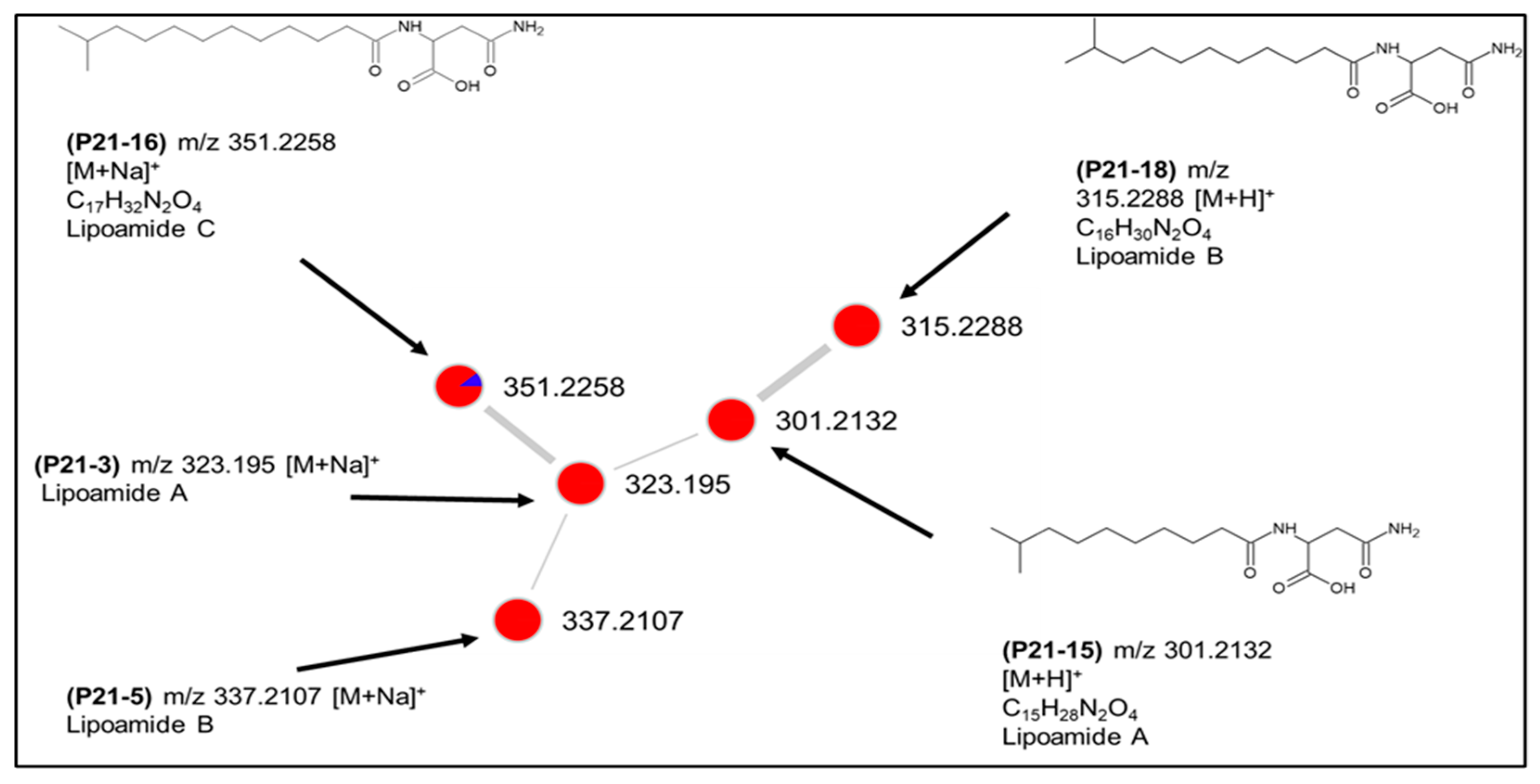
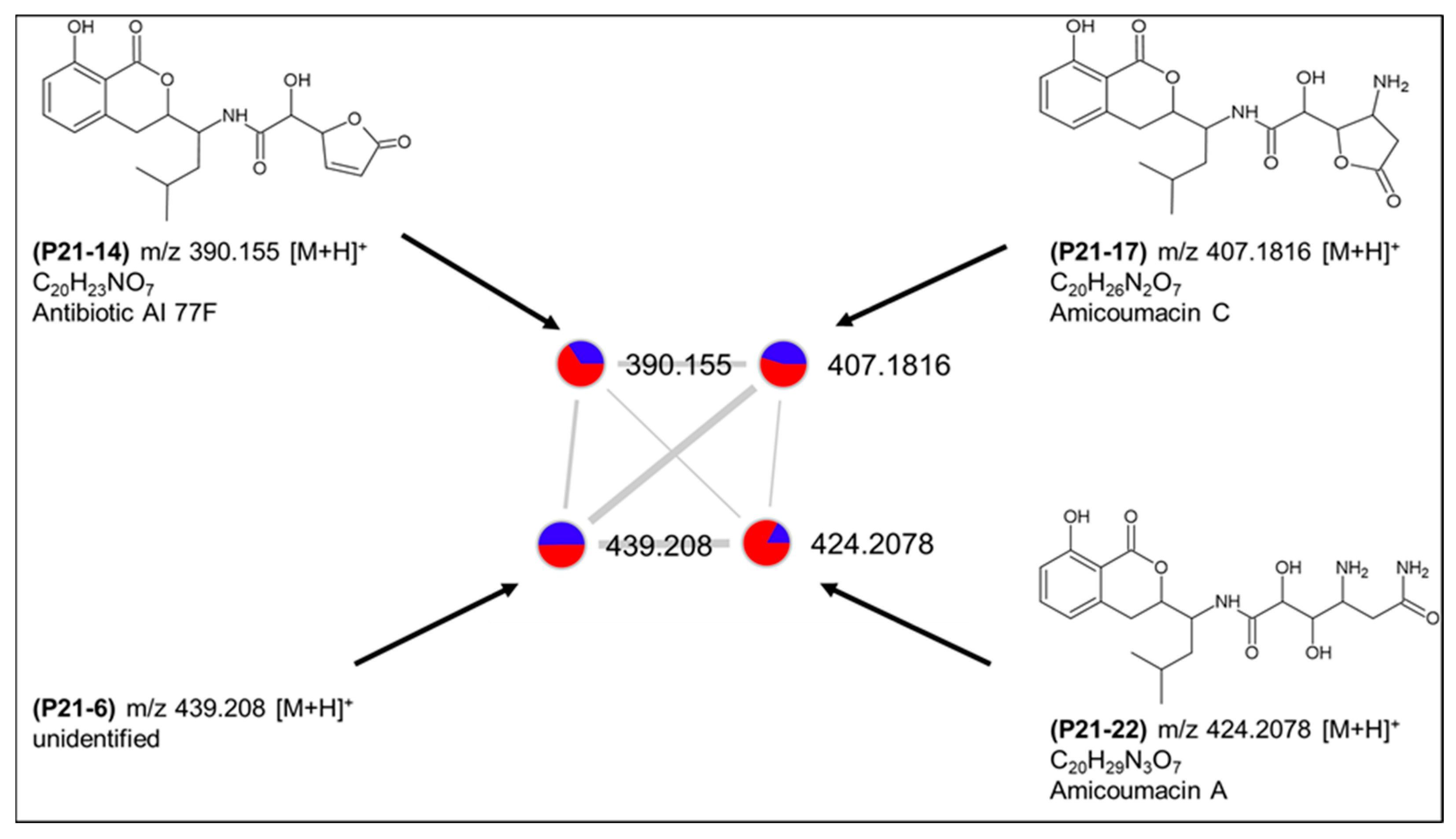
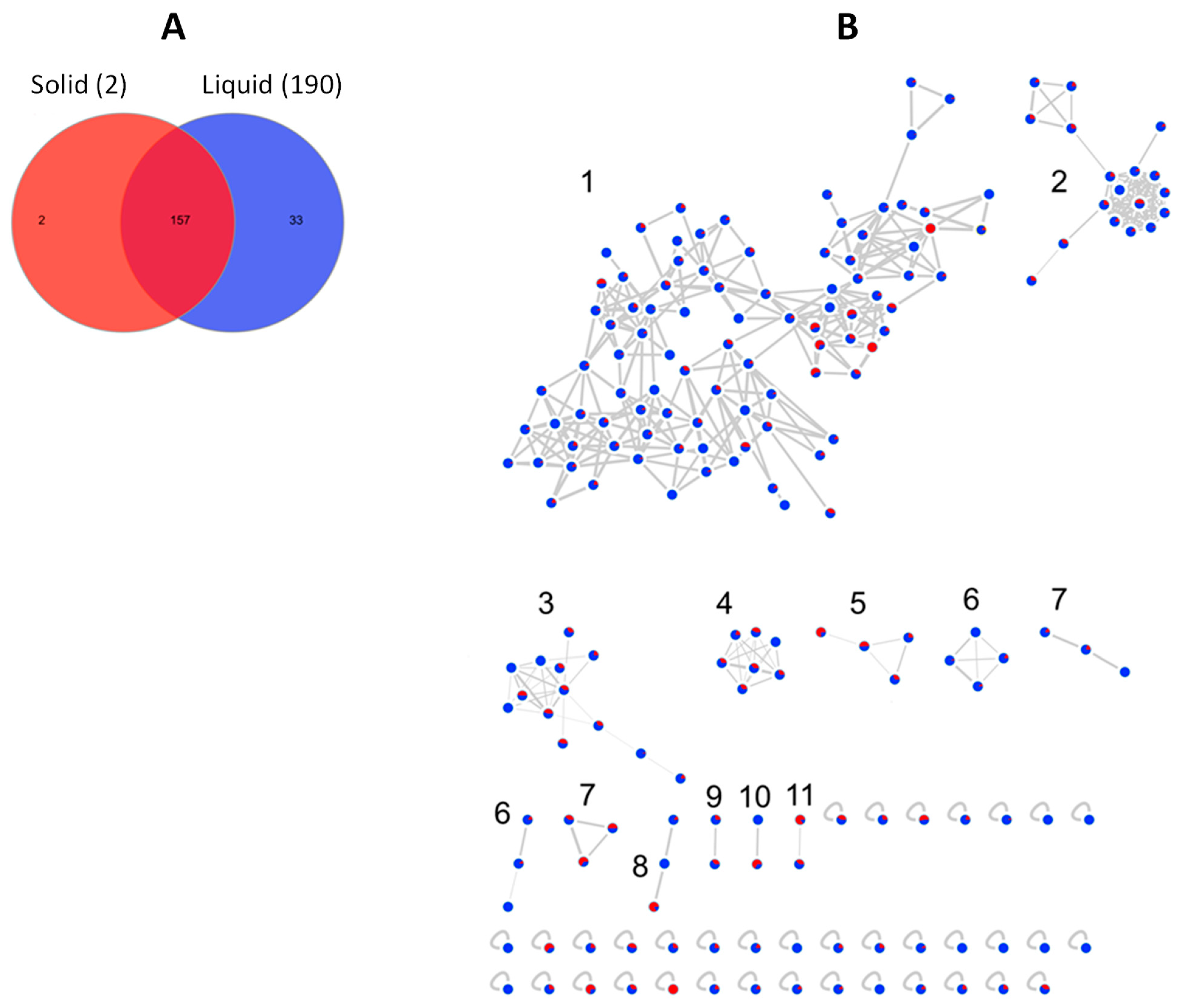
2.3. Molecular Network-Based Untargeted Metabolomics
3. Discussion
4. Materials and Methods
4.1. General Procedures
4.2. Sampling and Dissections
4.3. Isolation and Identification of Microorganisms
4.4. Cultivation
4.5. Extraction
4.6. UPLC-QToF-MS/MS-Based Metabolome Analyses
4.7. Bioactivity Assessments
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bjorbaekmo, M.F.M.; Evenstad, A.; Rosaeg, L.L.; Krabberod, A.K.; Logares, R. The planktonic protist interactome: Where do we stand after a century of research? ISME J. 2020, 14, 544–559. [Google Scholar] [CrossRef]
- Christie-Oleza, J.A.; Sousoni, D.; Lloyd, M.; Armengaud, J.; Scanlan, D.J. Nutrient recycling facilitates long-term stability of marine microbial phototroph-heterotroph interactions. Nat. Microbiol. 2017, 2, 17100. [Google Scholar] [CrossRef] [PubMed]
- Troussellier, M.; Escalas, A.; Bouvier, T.; Mouillot, D. Sustaining rare marine microorganisms: Macroorganisms as repositories and dispersal agents of microbial diversity. Front. Microbiol. 2017, 8, 947. [Google Scholar] [CrossRef]
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef]
- Dhanasiri, A.K.; Brunvold, L.; Brinchmann, M.F.; Korsnes, K.; Bergh, O.; Kiron, V. Changes in the intestinal microbiota of wild Atlantic cod Gadus morhua L. upon captive rearing. Microb. Ecol. 2011, 61, 20–30. [Google Scholar] [CrossRef]
- Tarnecki, A.M.; Burgos, F.A.; Ray, C.L.; Arias, C.R. Fish intestinal microbiome: Diversity and symbiosis unravelled by metagenomics. J. Appl. Microbiol. 2017, 123, 2–17. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Buonocore, F.; Gerdol, M. Alternative adaptive immunity strategies: Coelacanth, cod and shark immunity. Mol. Immunol. 2016, 69, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Wiles, T.J.; Jemielita, M.; Baker, R.P.; Schlomann, B.H.; Logan, S.L.; Ganz, J.; Melancon, E.; Eisen, J.S.; Guillemin, K.; Parthasarathy, R. Host gut motility promotes competitive exclusion within a model intestinal microbiota. PLoS Biol. 2016, 14, e1002517. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, W.; Tan, R. Symbionts, a promising source of bioactive natural products. Sci. China Chem. 2015, 58, 1097–1109. [Google Scholar] [CrossRef]
- FishBase. Available online: https://www.fishbase.de (accessed on 3 June 2022).
- Sullam, K.E.; Essinger, S.D.; Lozupone, C.A.; O’Connor, M.P.; Rosen, G.L.; Knight, R.; Kilham, S.S.; Russell, J.A. Environmental and ecological factors that shape the gut bacterial communities of fish: A meta-analysis. Mol. Ecol. 2012, 21, 3363–3378. [Google Scholar] [CrossRef] [Green Version]
- Millner, R.; Walsh, S.J.; Diaz de Astarloa, J.M. Atlantic flatfish fisheries. In Flatfishes: Biology and Exploitation; Gibson, R.N., Ed.; Blackwell Science: Oxford, UK, 2005; pp. 240–271. [Google Scholar]
- Hoarau, G.; Rijnsdorp, A.D.; van der Veer, H.W.; Stam, T.; Olsen, J.L. Population structure of plaice (Pleuronectes platessa L.) in northern Europe: Microsatellites revealed large-scale spatial and temporal homogeneity. Mol. Ecol. 2002, 11, 1165–1176. [Google Scholar] [CrossRef]
- Picot, L.; Bordenave, S.; Didelot, S.; Fruitier-Arnaudin, I.; Sannier, F.; Thorkelsson, G.; Berge, J.P.; Guerard, F.; Chabeaud, A.; Piot, J.M. Antiproliferative activity of fish protein hydrolysates on human breast cancer cell lines. Process Biochem. 2006, 41, 1217–1222. [Google Scholar] [CrossRef]
- Gilmour, A.; McCallum, M.F.; Allan, M.C. A study on the bacterial types occurring on the skin and in the intestines of farmed plaice, Pleuronectes platessa. Aquaculture 1976, 7, 161–172. [Google Scholar] [CrossRef]
- Wegner, K.M.; Shama, L.N.S.; Kellnreitner, F.; Pockberger, M. Diversity of immune genes and associated gill microbes of European plaice Pleuronectes platessa. Estuar. Coast. Shelf Sci. 2012, 108, 87–96. [Google Scholar] [CrossRef]
- Morris, M.T.; Hauton, C.; Baylay, A.J.; Peruzza, L.; Targett, T.E.; Ciotti, B.J. Spatial variation in the gastrointestinal microbiome, diet and nutritional condition of a juvenile flatfish among coastal habitats. Mar. Environ. Res. 2021, 170, 105413. [Google Scholar] [CrossRef]
- Griffith, G.W.; Easton, G.L.; Detheridge, A.; Roderick, K.; Edwards, A.; Worgan, H.J.; Nicholson, J.; Perkins, W.T. Copper deficiency in potato dextrose agar causes reduced pigmentation in cultures of various fungi. FEMS Microbiol. Lett. 2007, 276, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Pincus, D.H.; Orenga, S.; Chatellier, S. Yeast identification—Past, present, and future methods. Med. Mycol. 2007, 45, 97–121. [Google Scholar] [CrossRef] [PubMed]
- Chianese, G.; Esposito, F.P.; Parrot, D.; Ingham, C.; de Pascale, D.; Tasdemir, D. Linear aminolipids with moderate antimicrobial activity from the Antarctic gram-negative bacterium Aequorivita sp. Mar. Drugs 2018, 16, 187. [Google Scholar] [CrossRef]
- Palma Esposito, F.; Ingham, C.J.; Hurtado-Ortiz, R.; Bizet, C.; Tasdemir, D.; de Pascale, D. Isolation by miniaturized culture chip of an Antarctic bacterium Aequorivita sp. with antimicrobial and anthelmintic activity. Biotechnol. Rep. 2018, 20, e00281. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [Green Version]
- Nothias, L.F.; Petras, D.; Schmid, R.; Duhrkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-based molecular networking in the GNPS analysis environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef]
- Vanounou, S.; Parola, A.H.; Fishov, I. Phosphatidylethanolamine and phosphatidylglycerol are segregated into different domains in bacterial membrane. A study with pyrene-labelled phospholipids. Mol. Microbiol. 2003, 49, 1067–1079. [Google Scholar] [CrossRef]
- Berrue, F.; Ibrahim, A.; Boland, P.; Kerr, R.G. Newly isolated marine Bacillus pumilus (SP21): A source of novel lipoamides and other antimicrobial agents. Pure Appl. Chem. 2009, 81, 1027–1031. [Google Scholar] [CrossRef]
- Itoh, J.; Omoto, S.; Nishizawa, N.; Kodama, Y.; Inouye, S. Chemical structures of amicoumacins produced by Bacillus pumilus. Agric. Biol. Chem. 1982, 46, 2659–2665. [Google Scholar] [CrossRef]
- Shimojima, Y.; Hayashi, H.; Ooka, T.; Shibukawa, M.; Iitaka, Y. Studies on AI-77s, microbial products with gastroprotective activity. Structures and the chemical nature of AI-77s. Tetrahedron 1984, 40, 2519–2527. [Google Scholar] [CrossRef]
- Park, H.B.; Perez, C.E.; Perry, E.K.; Crawford, J.M. Activating and attenuating the amicoumacin antibiotics. Molecules 2016, 21, 824. [Google Scholar] [CrossRef] [PubMed]
- Vértesy, L.; Fehlhaber, H.-W.; Kogler, H.; Schindler, P.W. Enkastines: Amadori products with a specific inhibiting action against endopeptidase–24.11–from Streptomyces albus and by synthesis. Liebigs Annalen 1996, 1996, 121–126. [Google Scholar] [CrossRef]
- Isono, K.; Nagatsu, J.; Kobinata, K.; Sasaki, K.; Suzuki, S. Studies on polyoxins, antifungal antibiotics. Agric. Biol. Chem. 1967, 31, 190–199. [Google Scholar]
- Zhang, J.; Jiang, Y.; Cao, Y.; Liu, J.; Zheng, D.; Chen, X.; Han, L.; Jiang, C.; Huang, X. Violapyrones A-G, alpha-pyrone derivatives from Streptomyces violascens isolated from Hylobates hoolock feces. J. Nat. Prod. 2013, 76, 2126–2130. [Google Scholar] [CrossRef]
- Cvejic, J.H.; Bodrossy, L.; Kovacs, K.L.; Rohmer, M. Bacterial triterpenoids of the hopane series from the methanotrophic bacteria Methylocaldum spp.: Phylogenetic implications and first evidence for an unsaturated aminobacteriohopanepolyol. FEMS Microbiol. Lett. 2000, 182, 361–365. [Google Scholar] [CrossRef]
- Li, H.; Shinde, P.B.; Lee, H.J.; Yoo, E.S.; Lee, C.O.; Hong, J.; Choi, S.H.; Jung, J.H. Bile acid derivatives from a sponge-associated bacterium Psychrobacter sp. Arch. Pharm. Res. 2009, 32, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Engene, N.; Choi, H.; Esquenazi, E.; Byrum, T.; Villa, F.A.; Cao, Z.; Murray, T.F.; Dorrestein, P.C.; Gerwick, L.; Gerwick, W.H. Phylogeny-guided isolation of ethyl tumonoate A from the marine cyanobacterium cf. Oscillatoria margaritifera. J. Nat. Prod. 2011, 74, 1737–1743. [Google Scholar] [CrossRef] [PubMed]
- Roggo, B.E.; Petersen, F.; Delmendo, R.; Jenny, H.B.; Peter, H.H.; Roesel, J. 3-Alkanoyl-5-hydroxymethyl tetronic acid homologues and resistomycin: New inhibitors of HIV-1 protease. I. Fermentation, isolation and biological activity. J. Antibiot. 1994, 47, 136–142. [Google Scholar] [CrossRef]
- Huang, H.; Yao, Y.; He, Z.; Yang, T.; Ma, J.; Tian, X.; Li, Y.; Huang, C.; Chen, X.; Li, W.; et al. Antimalarial β-carboline and indolactam alkaloids from Marinactinospora thermotolerans, a deep sea isolate. J. Nat. Prod. 2011, 74, 2122–2127. [Google Scholar] [CrossRef]
- Gu, C.Z.; Yuan, S.H.; Lu, J.; Qiao, Y.J.; Song, Y.Y.; Abdalla Elzaki, M.E.; Yang, C.R.; Zhang, Y.J.; Zeng, R.S. Albocycline-type Macrolides with Antibacterial Activities from Streptomyces sp. 4205. Chem. Biodivers. 2019, 16, e1800344. [Google Scholar] [CrossRef]
- Kurosawa, T.; Sakai, K.; Nakahara, T.; Oshima, Y.; Tabuch, T. Extracellular accumulation of the polyol lipids, 3,5-dihydroxydecanoyl and 5-hydroxy-2-decenoyl esters of arabitol and mannitol, by Aureobasidium sp. Biosci. Biotechnol. Biochem. 1994, 58, 2057–2060. [Google Scholar] [CrossRef]
- Viegelmann, C.; Margassery, L.M.; Kennedy, J.; Zhang, T.; O’Brien, C.; O’Gara, F.; Morrissey, J.P.; Dobson, A.D.; Edrada-Ebel, R. Metabolomic profiling and genomic study of a marine sponge-associated Streptomyces sp. Mar. Drugs 2014, 12, 3323–3351. [Google Scholar] [CrossRef] [PubMed]
- Klausner, R.D.; Donaldson, J.G.; Lippincott-Schwartz, J. Brefeldin A: Insights into the control of membrane traffic and organelle structure. J. Cell Biol. 1992, 116, 1071–1080. [Google Scholar] [CrossRef]
- Pratte, Z.A.; Besson, M.; Hollman, R.D.; Stewart, F.J. The gills of reef fish support a distinct microbiome influenced by host-specific factors. Appl. Environ. Microbiol. 2018, 84, e00063-18. [Google Scholar] [CrossRef]
- Legrand, T.P.R.A.; Wynne, J.W.; Weyrich, L.S.; Oxley, A.P.A. A microbial sea of possibilities: Current knowledge and prospects for an improved understanding of the fish microbiome. Rev. Aquacult. 2020, 12, 1101–1134. [Google Scholar] [CrossRef]
- Yu, Y.-Y.; Ding, L.-G.; Huang, Z.-Y.; Xu, H.-Y.; Xu, Z. Commensal bacteria-immunity crosstalk shapes mucosal homeostasis in teleost fish. Rev. Aquacult. 2021, 13, 2322–2343. [Google Scholar] [CrossRef]
- Rawls, J.F.; Mahowald, M.A.; Ley, R.E.; Gordon, J.I. Reciprocal gut microbiota transplants from zebrafish and mice to germ-free recipients reveal host habitat selection. Cell 2006, 127, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Yi, Y.; Liang, L.; Shi, Q. High throughput identification of antimicrobial peptides from fish gastrointestinal microbiota. Toxins 2017, 9, 266. [Google Scholar] [CrossRef] [PubMed]
- Sharifuzzaman, S.M.; Austin, B. Probiotics for Disease Control in Aquaculture. In Diagnosis and Control of Diseases of Fish and Shellfish; Austin, B., Newaj-Fyzul, A., Eds.; John Wiley & Sons: Chichester, WestSussex, UK, 2017; pp. 189–209. [Google Scholar]
- Sanchez, L.M.; Wong, W.R.; Riener, R.M.; Schulze, C.J.; Linington, R.G. Examining the fish microbiome: Vertebrate-derived bacteria as an environmental niche for the discovery of unique marine natural products. PLoS ONE 2012, 7, e35398. [Google Scholar] [CrossRef]
- Yamada, T.; Kikuchi, T.; Tanaka, R.; Numata, A. Halichoblelides B and C, potent cytotoxic macrolides from a Streptomyces species separated from a marine fish. Tetrahedron Lett. 2012, 53, 2842–2846. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, K.; Wang, W.; Zhang, G.; Zhu, T.; Che, Q.; Gu, Q.; Li, D. Amphiepicoccins A-J: Epipolythiodioxopiperazines from the Fish-Gill-Derived Fungus Epicoccum nigrum HDN17-88. J. Nat. Prod. 2020, 83, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, O.G.; Salim, A.A.; Khalil, Z.G.; Elbanna, A.H.; Bernhardt, P.V.; Capon, R.J. Chrysosporazines F-M: P-glycoprotein inhibitory phenylpropanoid piperazines from an Australian marine fish derived fungus, Chrysosporium sp. CMB-F294. J. Nat. Prod. 2020, 83, 497–504. [Google Scholar] [CrossRef]
- Maina, J.N. Structure, function and evolution of the gas exchangers: Comparative perspectives. J. Anat. 2002, 201, 281–304. [Google Scholar] [CrossRef]
- Kelly, C.; Salinas, I. Under Pressure: Interactions between commensal microbiota and the teleost immune system. Front. Immunol. 2017, 8, 559. [Google Scholar] [CrossRef]
- Schram, E.; Roques, J.A.C.; Abbink, W.; Spanings, T.; de Vries, P.; Bierman, S.; de Vis, H.v.; Flik, G. The impact of elevated water ammonia concentration on physiology, growth and feed intake of African catfish (Clarias gariepinus). Aquaculture 2010, 306, 108–115. [Google Scholar] [CrossRef]
- Marshall, W.S.; Bellamy, D. The 50 year evolution of in vitro systems to reveal salt transport functions of teleost fish gills. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2010, 155, 275–280. [Google Scholar] [CrossRef]
- Peatman, E.; Lange, M.; Zhao, H.; Beck, B.H. Physiology and immunology of mucosal barriers in catfish (Ictalurus spp.). Tissue Barriers 2015, 3, e1068907. [Google Scholar] [CrossRef]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost microbiomes: The state of the art in their characterization, manipulation and importance in aquaculture and fisheries. Front. Microbiol. 2014, 5, 207. [Google Scholar] [CrossRef] [PubMed]
- Merrifield, D.L.; Rodiles, A. The fish microbiome and its interactions with mucosal tissues. In Mucosal Health in Aquaculture; Beck, B.H., Peatman, E., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 273–295. [Google Scholar]
- Lokesh, J.; Kiron, V. Transition from freshwater to seawater reshapes the skin-associated microbiota of Atlantic salmon. Sci. Rep. 2016, 6, 19707. [Google Scholar] [CrossRef]
- Lowrey, L.; Woodhams, D.C.; Tacchi, L.; Salinas, I. Topographical mapping of the rainbow trout (Oncorhynchus mykiss) microbiome reveals a diverse bacterial community with antifungal properties in the skin. Appl. Environ. Microbiol. 2015, 81, 6915–6925. [Google Scholar] [CrossRef] [PubMed]
- Reverter, M.; Sasal, P.; Tapissier-Bontemps, N.; Lecchini, D.; Suzuki, M. Characterisation of the gill mucosal bacterial communities of four butterflyfish species: A reservoir of bacterial diversity in coral reef ecosystems. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef]
- Chen, P.; Li, J.; Li, Q.X.; Wang, Y.; Li, S.; Ren, T.; Wang, L. Simultaneous heterotrophic nitrification and aerobic denitrification by bacterium Rhodococcus sp. CPZ24. Bioresour. Technol. 2012, 116, 266–270. [Google Scholar] [CrossRef]
- Yoshida, N.; Inaba, S.; Takagi, H. Utilization of atmospheric ammonia by an extremely oligotrophic bacterium, Rhodococcus erythropolis N9T-4. J. Biosci. Bioeng. 2014, 117, 28–32. [Google Scholar] [CrossRef]
- Vianey, P.G.C.; del Carmen, M.D.M.; Ramirez Tórrez, J.A.; Ocampo Cervantes, J.A.; Castro Mejía, J. Rhodococcus sp. as probiotic bacteria for increase the survival, growth and coloration of fish Puntius conchonius. Sci. J. Anim. Sci. 2016, 5, 370–375. [Google Scholar]
- Sharifuzzaman, S.M.; Rahman, H.; Austin, D.A.; Austin, B. Properties of probiotics Kocuria SM1 and Rhodococcus SM2 isolated from fish guts. Probiotics Antimicrob. Proteins 2018, 10, 534–542. [Google Scholar] [CrossRef]
- Montalban-Arques, A.; De Schryver, P.; Bossier, P.; Gorkiewicz, G.; Mulero, V.; Gatlin, D.M., 3rd; Galindo-Villegas, J. Selective manipulation of the gut microbiota improves immune status in vertebrates. Front. Immunol. 2015, 6, 512. [Google Scholar] [CrossRef] [PubMed]
- Hansen, G.H.; Olafsen, J.A. Bacterial Colonization of Cod (Gadus morhua L.) and Halibut (Hippoglossus hippoglossus) Eggs in Marine Aquaculture. Appl. Environ. Microbiol. 1989, 55, 1435–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergh, Ø. Bacteria associated with early life stages of halibut, Hippoglossus hippoglossus L., inhibit growth of a pathogenic Vibrio sp. J. Fish Dis. 1995, 18, 31–40. [Google Scholar] [CrossRef]
- Grisez, L.; Reyniers, J.; Verdonck, L.; Swings, J.; Ollevier, F. Dominant intestinal microflora of sea bream and sea bass larvae, from two hatcheries, during larval development. Aquaculture 1997, 155, 387–399. [Google Scholar] [CrossRef]
- Wang, A.R.; Ran, C.; Ringø, E.; Zhou, Z.G. Progress in fish gastrointestinal microbiota research. Rev. Aquacult. 2018, 10, 626–640. [Google Scholar] [CrossRef]
- Roeselers, G.; Mittge, E.K.; Stephens, W.Z.; Parichy, D.M.; Cavanaugh, C.M.; Guillemin, K.; Rawls, J.F. Evidence for a core gut microbiota in the zebrafish. ISME J. 2011, 5, 1595–1608. [Google Scholar] [CrossRef]
- Navarrete, P.; Magne, F.; Araneda, C.; Fuentes, P.; Barros, L.; Opazo, R.; Espejo, R.; Romero, J. PCR-TTGE analysis of 16S rRNA from rainbow trout (Oncorhynchus mykiss) gut microbiota reveals host-specific communities of active bacteria. PLoS ONE 2012, 7, e31335. [Google Scholar] [CrossRef]
- Givens, C.E.; Ransom, B.; Bano, N.; Hollibaugh, J.T. Comparison of the gut microbiomes of 12 bony fish and 3 shark species. Mar. Ecol. Prog. Ser. 2015, 518, 209–223. [Google Scholar] [CrossRef]
- Ghanbari, M.; Kneifel, W.; Domig, K.J. A new view of the fish gut microbiome: Advances from next-generation sequencing. Aquaculture 2015, 448, 464–475. [Google Scholar] [CrossRef]
- Egerton, S.; Culloty, S.; Whooley, J.; Stanton, C.; Ross, R.P. The gut microbiota of marine fish. Front. Microbiol. 2018, 9, 873. [Google Scholar] [CrossRef]
- Estruch, G.; Collado, M.C.; Penaranda, D.S.; Tomas Vidal, A.; Jover Cerda, M.; Perez Martinez, G.; Martinez-Llorens, S. Impact of fishmeal replacement in diets for gilthead sea bream (Sparus aurata) on the gastrointestinal microbiota determined by pyrosequencing the 16S rRNA gene. PLoS ONE 2015, 10, e0136389. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.; Austin, B.; Austin, D.A.; Larsen, J.L. Vibrios associated with mortality in cultured plaice Pleuronectes platessa fry. Acta Vet. Scand. 1999, 40, 263–270. [Google Scholar] [CrossRef]
- Harrison, J.; Nelson, K.; Morcrette, H.; Morcrette, C.; Preston, J.; Helmer, L.; Titball, R.W.; Butler, C.S.; Wagley, S. The increased prevalence of Vibrio species and the first reporting of Vibrio jasicida and Vibrio rotiferianus at UK shellfish sites. Water Res. 2022, 211, 117942. [Google Scholar] [CrossRef] [PubMed]
- Rolig, A.S.; Parthasarathy, R.; Burns, A.R.; Bohannan, B.J.; Guillemin, K. Individual members of the microbiota disproportionately modulate host innate immune responses. Cell Host Microbe 2015, 18, 613–620. [Google Scholar] [CrossRef]
- Mohamed, O.G.; Khalil, Z.G.; Capon, R.J. Prolinimines: N-Amino-l-pro-methyl ester (hydrazine) schiff bases from a fish gastrointestinal tract-derived fungus, Trichoderma sp. CMB-F563. Org. Lett. 2018, 20, 377–380. [Google Scholar] [CrossRef]
- Elbanna, A.H.; Agampodi Dewa, A.; Khalil, Z.G.; Capon, R.J. Precursor-directed biosynthesis mediated amplification of minor aza phenylpropanoid piperazines in an Australian marine fish-gut-derived fungus, Chrysosporium sp. CMB-F214. Mar. Drugs 2021, 19, 478. [Google Scholar] [CrossRef]
- Elbanna, A.H.; Khalil, Z.G.; Bernhardt, P.V.; Capon, R.J. Scopularides revisited: Molecular networking guided exploration of lipodepsipeptides in Australian marine fish gastrointestinal tract-derived fungi. Mar. Drugs 2019, 17, 475. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, O.G.; Khalil, Z.G.; Santiago, V.; Capon, R.J. Metarhizides A–C and metarhizosides A–B: PKS-NRPS macrolides and aromatic glycosides from an Australian fish gut-derived fungus, Metarhizium sp. CMB-F624. Tetrahedron 2022, 113, 132759. [Google Scholar] [CrossRef]
- Dehler, C.E.; Secombes, C.J.; Martin, S.A. Environmental and physiological factors shape the gut microbiota of Atlantic salmon parr (Salmo salar L.). Aquaculture 2017, 467, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Cahill, M.M. Bacterial flora of fishes: A review. Microb. Ecol. 1990, 19, 21–41. [Google Scholar] [CrossRef]
- Still, P.C.; Johnson, T.A.; Theodore, C.M.; Loveridge, S.T.; Crews, P. Scrutinizing the scaffolds of marine biosynthetics from different source organisms: Gram-negative cultured bacterial products enter center stage. J. Nat. Prod. 2014, 77, 690–702. [Google Scholar] [CrossRef]
- Masschelein, J.; Jenner, M.; Challis, G.L. Antibiotics from gram-negative bacteria: A comprehensive overview and selected biosynthetic highlights. Nat. Prod. Rep. 2017, 34, 712–783. [Google Scholar] [CrossRef]
- Timmermans, M.L.; Paudel, Y.P.; Ross, A.C. Investigating the Biosynthesis of natural products from marine proteobacteria: A survey of molecules and strategies. Mar. Drugs 2017, 15, 235. [Google Scholar] [CrossRef]
- Overy, D.P.; Bayman, P.; Kerr, R.G.; Bills, G.F. An assessment of natural product discovery from marine (sensu strictu) and marine-derived fungi. Mycology 2014, 5, 145–167. [Google Scholar] [CrossRef]
- Tasdemir, D. Marine fungi in the spotlight: Opportunities and challenges for marine fungal natural product discovery and biotechnology. Fung. Biol. Biotechnol. 2017, 4, 5. [Google Scholar] [CrossRef]
- Butt, R.L.; Volkoff, H. Gut microbiota and energy homeostasis in fish. Front. Endocrinol. 2019, 10, 9. [Google Scholar] [CrossRef]
- Utermann, C.; Parrot, D.; Breusing, C.; Stuckas, H.; Staufenberger, T.; Blumel, M.; Labes, A.; Tasdemir, D. Combined genotyping, microbial diversity and metabolite profiling studies on farmed Mytilus spp. from Kiel Fjord. Sci. Rep. 2018, 8, 7983. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Olivon, F.; Grelier, G.; Roussi, F.; Litaudon, M.; Touboul, D. MZmine 2 data-preprocessing to enhance molecular networking reliability. Anal. Chem. 2017, 89, 7836–7840. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Oresic, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Allen, F.; Pon, A.; Wilson, M.; Greiner, R.; Wishart, D. CFM-ID: A web server for annotation, spectrum prediction and metabolite identification from tandem mass spectra. Nucleic Acids Res. 2014, 42, W94–W99. [Google Scholar] [CrossRef]
- Cicek, S.S.; Wenzel-Storjohann, A.; Girreser, U.; Tasdemir, D. Biological activities of two major copaiba diterpenoids and their semi-synthetic derivatives. Rev. Bras. Farmacogn. 2020, 30, 18–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Taxonomical ID | Origin | Medium/ Regime | Vi | Lg | MRSA | Ef |
|---|---|---|---|---|---|---|---|
| PG1-B | Shewanella baltica | Gill | MA solid | >100 | >100 | 20.2 | 26.4 |
| MB liquid | >100 | 41.7 | 18.8 | 8.8 | |||
| PG10-B | Chryseobacterium carnis | Gill | MA solid | 18.6 | >100 | 39.0 | 41.9 |
| MB liquid | >100 | >100 | >100 | 51.0 | |||
| PG11-B | Microbacterium sp. | Gill | MA solid | >100 | >100 | 45.9 | 86.6 |
| MB liquid | 5.7 | >100 | >100 | >100 | |||
| PG12-B | Vibrio aestuarianus | Gill | MA solid | 17.2 | 27.8 | 19.1 | 34.4 |
| MB liquid | 43.9 | >100 | 61.0 | 43.4 | |||
| PID21-B | Bacillus sp. | Gut (Intestine) Digesta | MA solid | 4.7 | >100 | 10.2 | >100 |
| MB liquid | 28.4 | >100 | 5.9 | >100 | |||
| PI1-B | Bacillus sp. | Gut (Intestine) Epithelium | MA solid | >100 | 14.7 | 18.5 | 15.0 |
| MB liquid | >100 | 54.2 | 22.1 | 11.6 | |||
| PI2-B | Vibrio sp. | Gut (Intestine) Epithelium | MA solid | >100 | 14.1 | 15.8 | 17.5 |
| MB liquid | >100 | >100 | 36.4 | 9.0 | |||
| PI8-B | Pseudoalteromonas sp. | Gut (Intestine) Epithelium | MA solid | >100 | 28.3 | 21.2 | 50.4 |
| MB liquid | >100 | 36.6 | 8.1 | 8.2 | |||
| PS3-B | Psychrobacter sp. | Gut (Stomach) Epithelium | MA solid | >100 | >100 | 23.3 | 28.9 |
| MB liquid | >100 | >100 | 11.4 | 7.5 | |||
| MCC-PID1-B | Photobacterium sp. | Gut (Intestine) Digesta | MA solid | >100 | 40.2 | 35.0 | 69.0 |
| MB liquid | n.g. | n.g. | n.g. | n.g. | |||
| PSD4-B | Shewanella colwelliana | Gut (Stomach) Digesta | MA solid | >100 | 25.4 | 29.1 | 30.8 |
| MB liquid | n.g. | n.g. | n.g. | n.g. | |||
| PID2-B | Shewanella aestuarii | Gut (Intestine) Digesta | MA solid | n.g. | n.g. | n.g. | n.g. |
| MB liquid | >100 | >100 | 6.8 | 4.9 | |||
| PI9-F | Aureobasidium pullulans | Gut (Intestine) Epithelium | PDA solid | >100 | 22.6 | 40.8 | 30.7 |
| PDA liquid | >100 | >100 | 8.9 | 3.3 | |||
| Positive control | 0.4 | 0.5 | 1.5 | 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghotbi, M.; Kelting, O.; Blümel, M.; Tasdemir, D. Gut and Gill-Associated Microbiota of the Flatfish European Plaice (Pleuronectes platessa): Diversity, Metabolome and Bioactivity against Human and Aquaculture Pathogens. Mar. Drugs 2022, 20, 573. https://doi.org/10.3390/md20090573
Ghotbi M, Kelting O, Blümel M, Tasdemir D. Gut and Gill-Associated Microbiota of the Flatfish European Plaice (Pleuronectes platessa): Diversity, Metabolome and Bioactivity against Human and Aquaculture Pathogens. Marine Drugs. 2022; 20(9):573. https://doi.org/10.3390/md20090573
Chicago/Turabian StyleGhotbi, Marjan, Ole Kelting, Martina Blümel, and Deniz Tasdemir. 2022. "Gut and Gill-Associated Microbiota of the Flatfish European Plaice (Pleuronectes platessa): Diversity, Metabolome and Bioactivity against Human and Aquaculture Pathogens" Marine Drugs 20, no. 9: 573. https://doi.org/10.3390/md20090573
APA StyleGhotbi, M., Kelting, O., Blümel, M., & Tasdemir, D. (2022). Gut and Gill-Associated Microbiota of the Flatfish European Plaice (Pleuronectes platessa): Diversity, Metabolome and Bioactivity against Human and Aquaculture Pathogens. Marine Drugs, 20(9), 573. https://doi.org/10.3390/md20090573