

Characterisation of Bioactive Peptides from Red Alga Gracilariopsis chorda


 and
and
Abstract
:1. Introduction
2. Results and Discussion
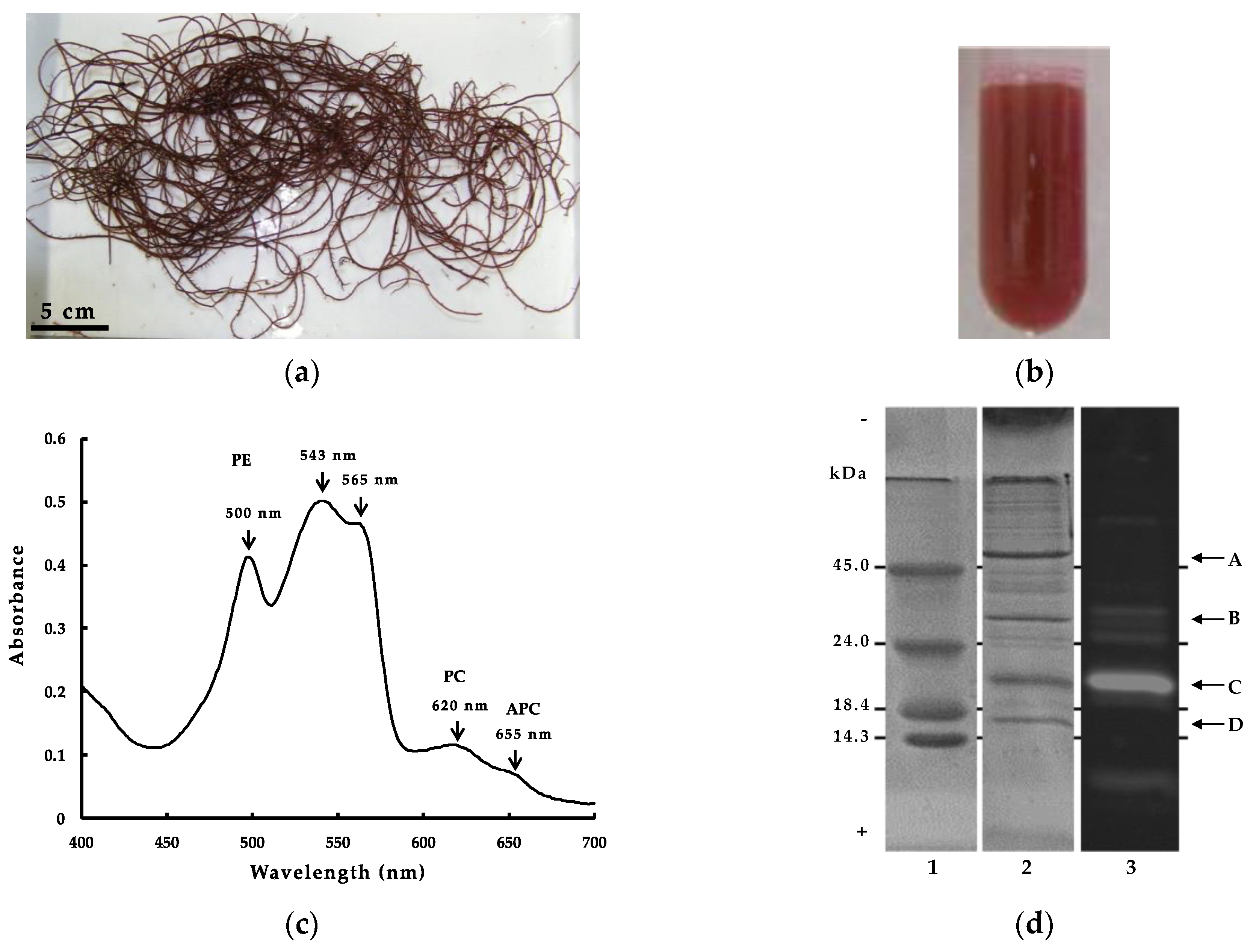
2.1. Characteristics of G. cholda WSP
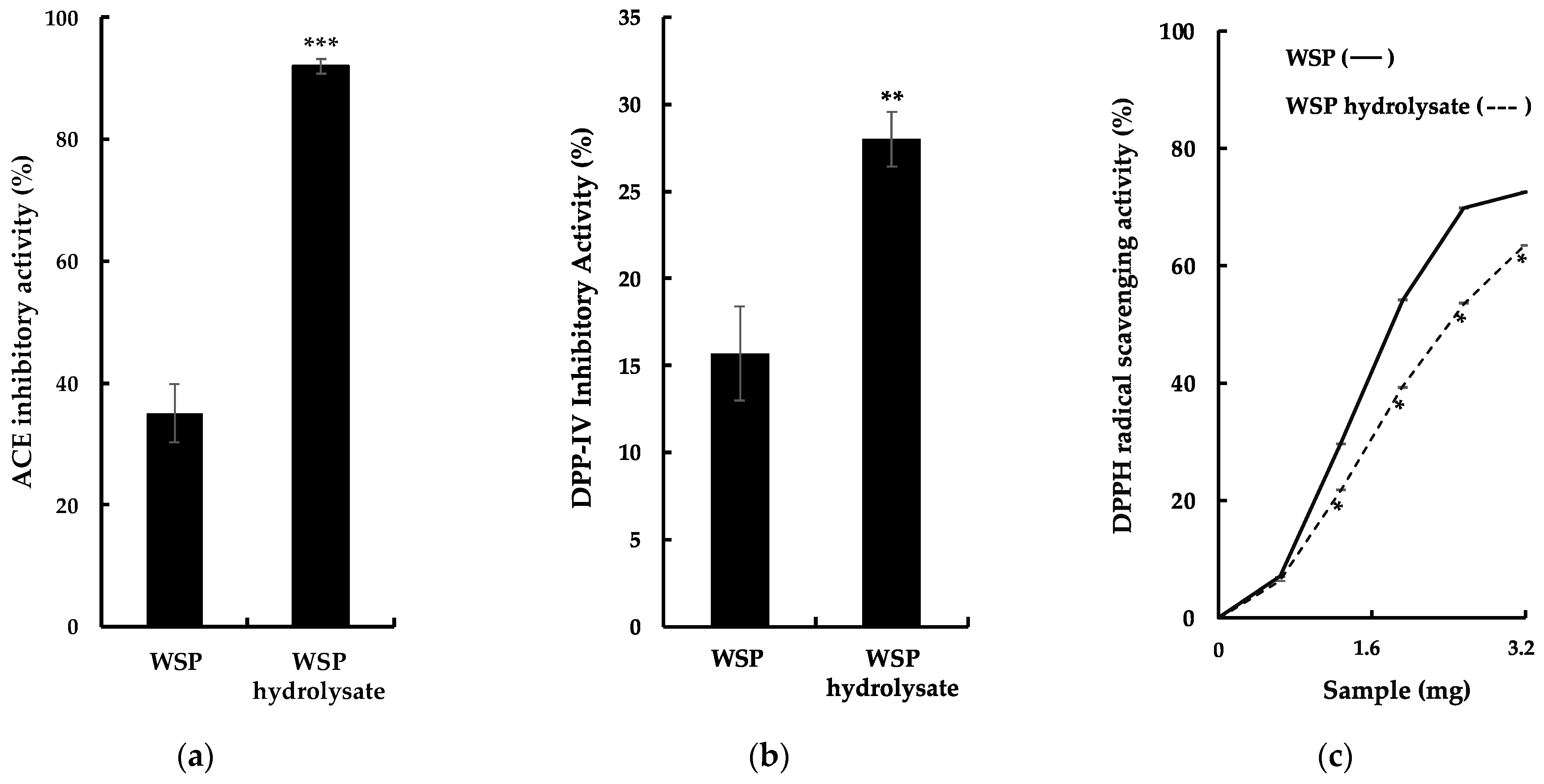
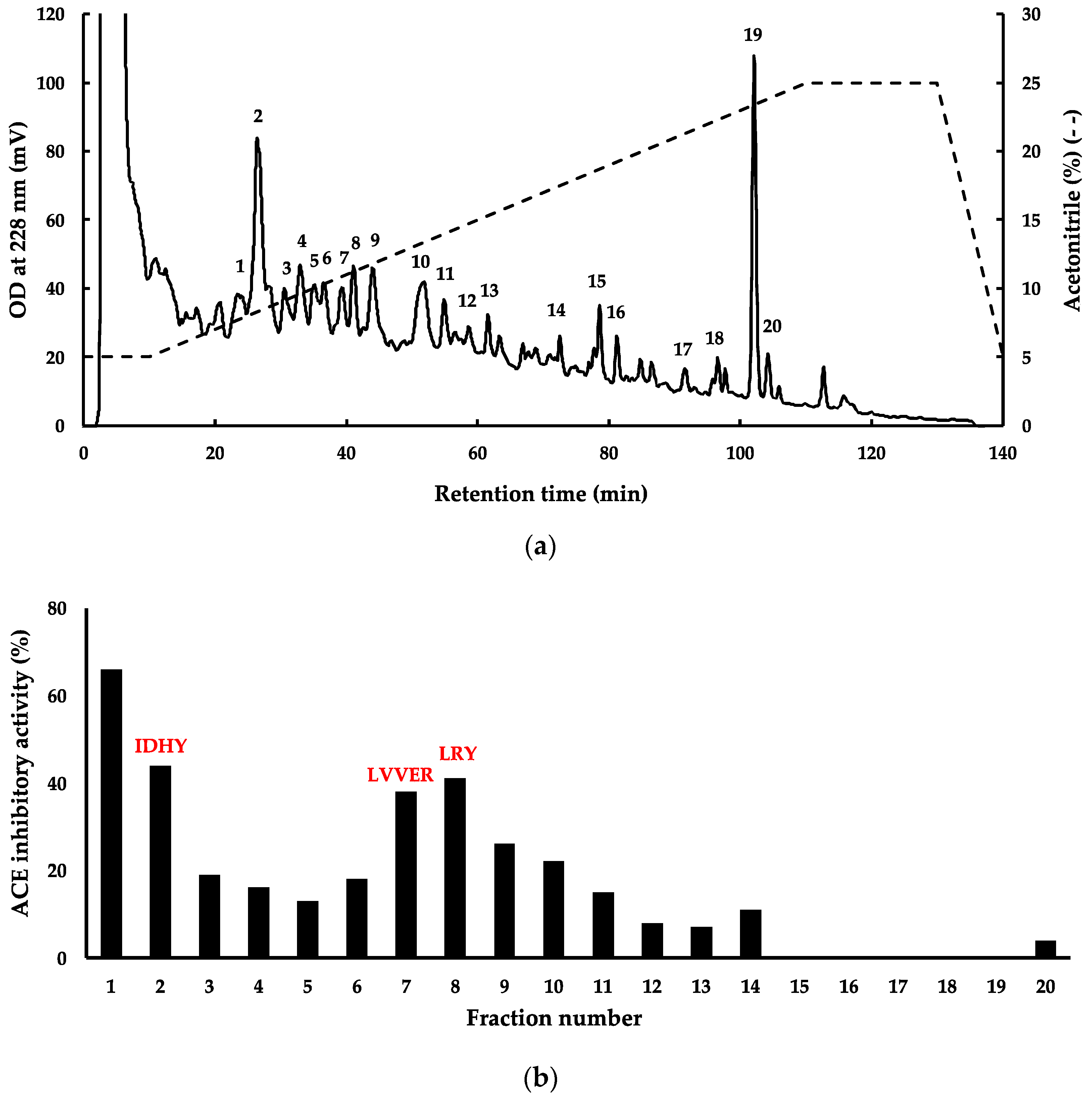
2.2. Bioactive Properties of G. chorda WSP Hydrolysate and Isolation of ACE Inhibitory Peptides
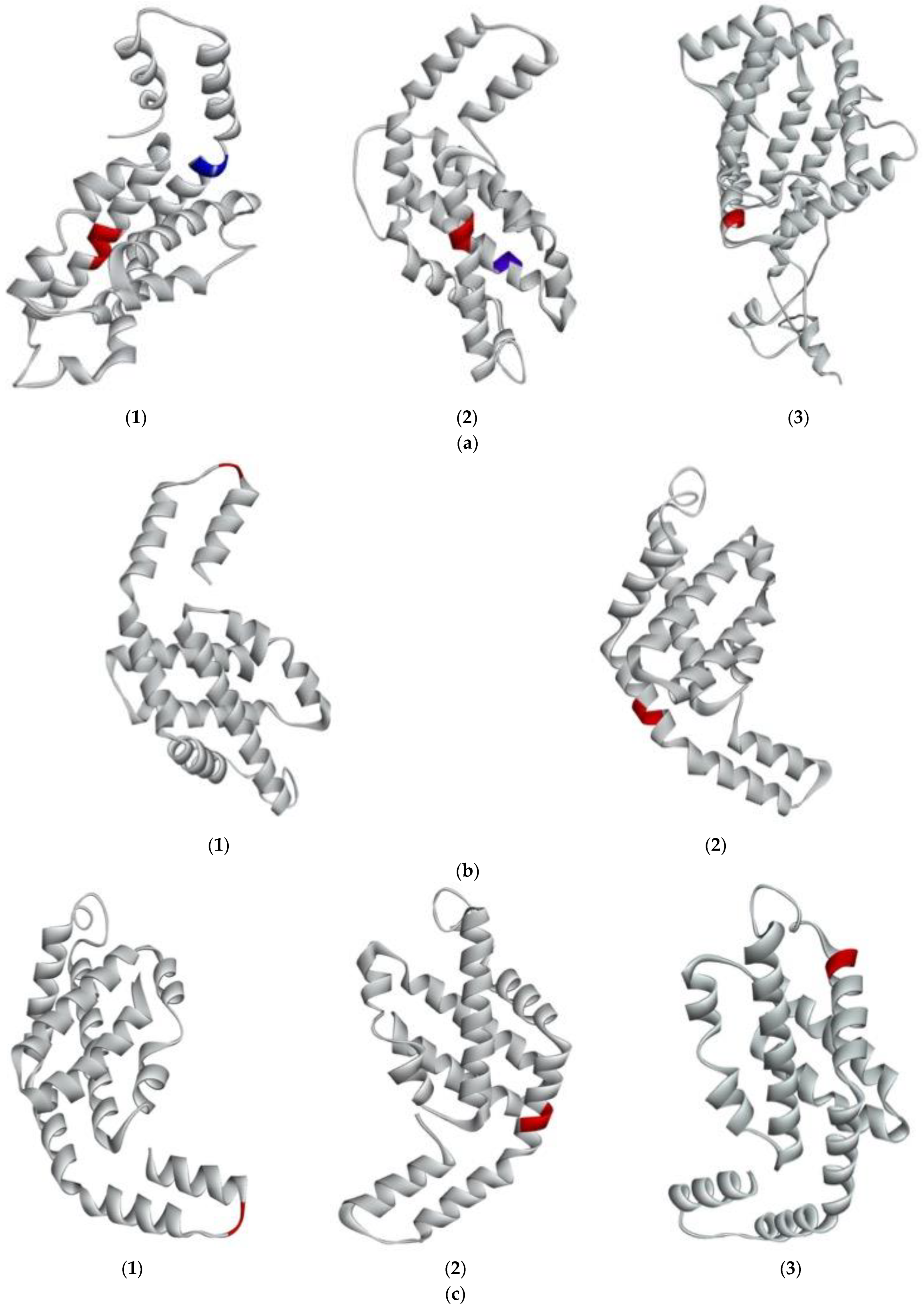
2.3. Structural Characterization of G. chorda WSP and Structure-Function Relationship
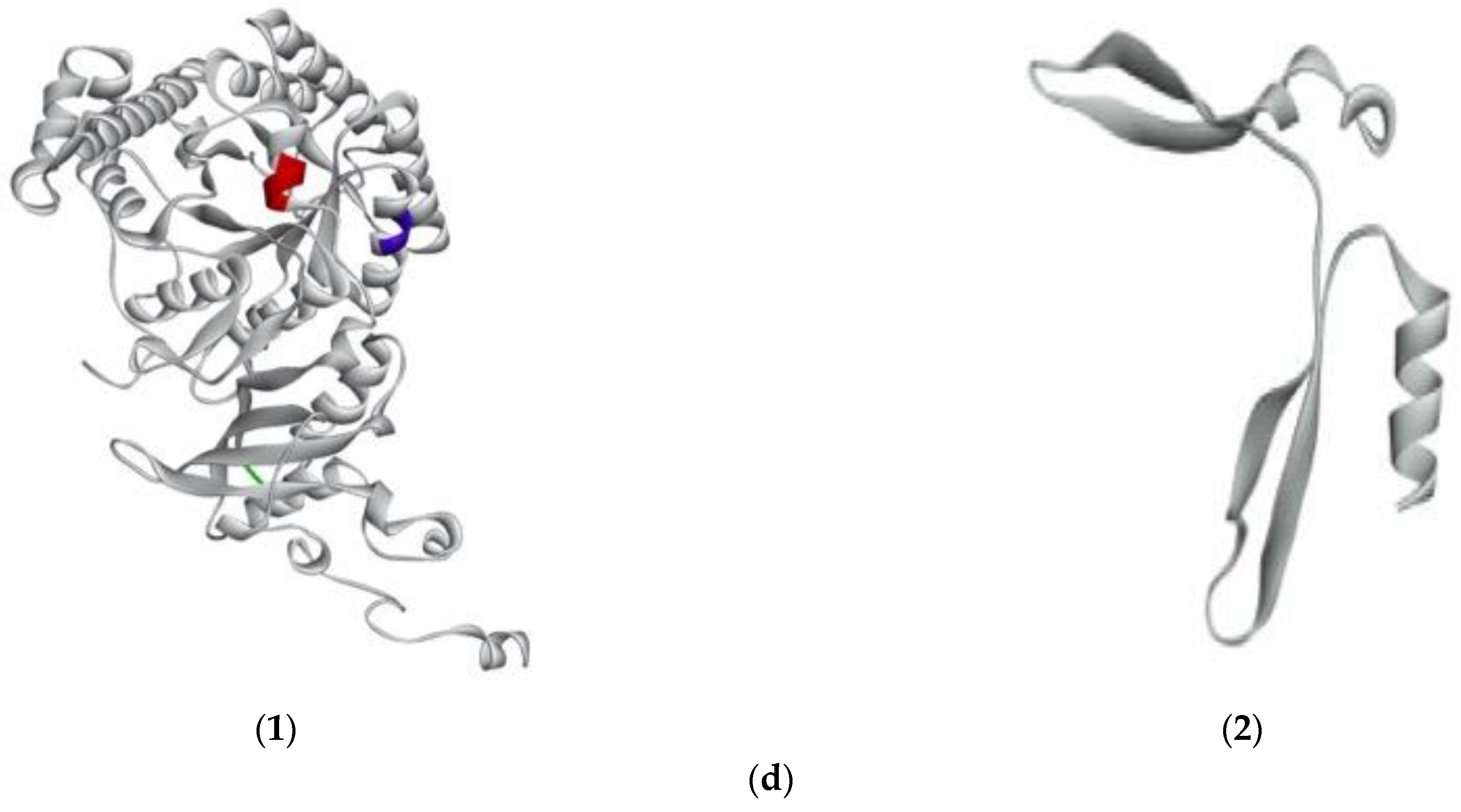
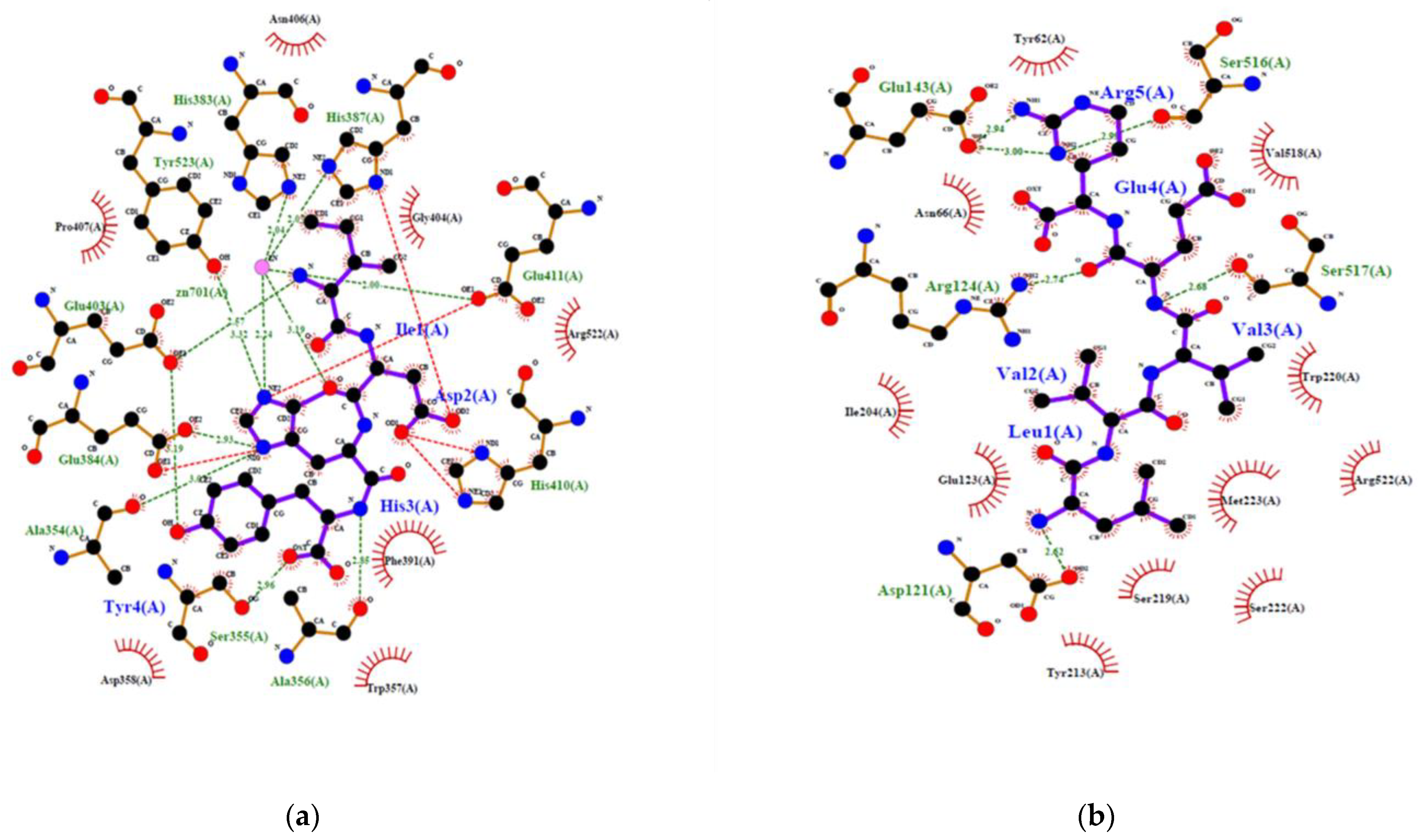
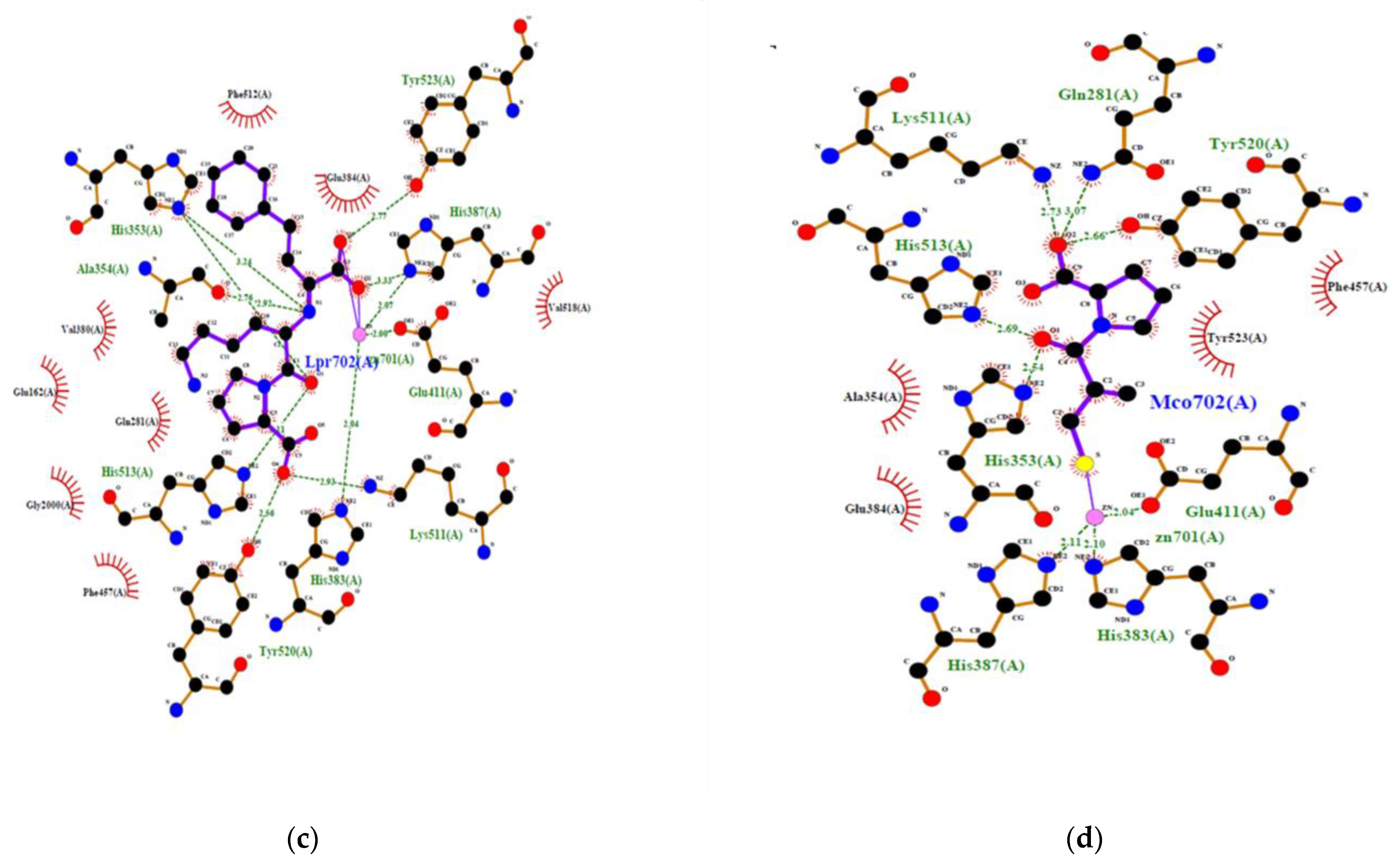
2.4. Prediction of the Interaction between New Bioactive Peptides and Human ACE
3. Materials and Methodsw
3.1. Materials
3.2. Preparation of G. chorda WSP Hydrolysate
3.3. ACE Inhibitory Assay
3.4. DPP-IV Inhibitory Assay
3.5. DPPH Radical Scavenging Assay
3.6. Separation of G. chorda WSP Hydrolysate
3.7. Statistical Analysis
3.8. Isolation and Sequencing of G. chorda DNA
3.9. In Silico Analysis
3.9.1. Building 3D Structure of G. chorda Phycobiliproteins and RuBisCo
3.9.2. Ligand Preparation
3.9.3. Protein Preparation
3.9.4. Grid Generation and Molecular Docking
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sanjeewa, K.K.; Lee, W.; Jeon, Y.J. Nutrients and bioactive potentials of edible green and red seaweed in Korea. Fish. Aquat. Sci. 2018, 21, 19. [Google Scholar] [CrossRef]
- Layse, C.; Almeida, F.; Falcão, H.D.S.; Lima, G.R.D.M.; Montenegro, C.D.A.; Lira, N.S.; de Athayde-Filho, P.F.; Rodrigues, L.C.; de Souza, M.F.V.; Barbosa-Filho, J.M.; et al. Bioactivities from marine algae of the genus Gracilaria. Int. J. Mol. Sci. 2011, 12, 4550–4573. [Google Scholar] [CrossRef] [Green Version]
- Furuta, T.; Miyabe, Y.; Yasui, H.; Kinoshita, Y.; Kishimura, H. Angiotensin I converting enzyme inhibitory peptides derived from phycobiliproteins of dulse Palmaria palmata. Mar. Drugs 2016, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Kumagai, Y.; Miyabe, Y.; Takeda, T.; Adachi, K.; Yasui, H.; Kishimura, H. In silico analysis of relationship between proteins from plastid genome of red alga Palmaria sp. (Japan) and angiotensin I converting enzyme inhibitory peptides. Mar. Drugs 2019, 17, 190. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, C.; Mora-Soler, L.; Gallagher, E.; O’Connor, P.; Prieto, J.; Soler-Vila, A.; Hayes, M. Isolation and characterization of bioactive pro-peptides with in vitro renin inhibitory activities from the macroalga Palmaria palmata. J. Agric. Food Chem. 2012, 60, 7421–7427. [Google Scholar] [CrossRef]
- Sato, N.; Furuta, T.; Takeda, T.; Miyabe, Y.; Ura, K.; Takagi, Y.; Yasui, H.; Kumagai, Y.; Kishimura, H. Antioxidant activity of proteins extracted from red alga dulse harvested in Japan. J. Food Biochem. 2018, 43, e12709. [Google Scholar] [CrossRef]
- Miyabe, Y.; Furuta, T.; Takeda, T.; Kanno, G.; Shimizu, T.; Tanaka, Y.; Gai, Z.; Yasui, H.; Kishimura, H. Structural properties of phycoerythrin from dulse Palmaria palmata. J. Food Biochem. 2017, 41, e12301. [Google Scholar] [CrossRef] [Green Version]
- Morikawa, R.; Toji, K.; Kumagai, Y.; Kishimura, H. ACE inhibitory effect of the protein hydrolysates prepared from commercially available nori product by pepsin–trypsin digestion. Eur. Food Res. Technol. 2022, 248, 243–251. [Google Scholar] [CrossRef]
- Windarto, S.; Lee, M.C.; Nursyam, H.; Hsu, J.L. First report of screening of novel angiotensin-I converting enzyme inhibitory peptides derived from the red alga Acrochaetium sp. Mar. Biotechnol. 2022, 24, 882–894. [Google Scholar] [CrossRef]
- Joel, C.H.; Sutopo, C.C.; Prajitno, A.; Su, J.H.; Hsu, J.L. Screening of angiotensin-I converting enzyme inhibitory peptides derived from Caulerpa lentillifera. Molecules 2018, 23, 3005. [Google Scholar] [CrossRef]
- Carbonaro, M.; Maselli, P.; Nucara, A. Structural aspects of legume proteins and nutraceutical properties. Food Res. Int. 2015, 76, 19–30. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, J.; Liu, D.; Qin, S.; Sun, S.; Zhao, J.; Sui, S.F. Structure of phycobilisome from the red alga Griffithsia pacifica. Nature 2017, 551, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, Y.; Toji, K.; Katsukura, S.; Morikawa, R.; Uji, T.; Yasui, H.; Shimizu, T.; Kishimura, H. Characterization of ACE Inhibitory peptides prepared from Pyropia pseudolinearis protein. Mar. Drugs 2021, 19, 200. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.; Bogorad, L. Complementary chromatic adaptation in a filamentous blue-green alga. J. Cell Biol. 1973, 58, 419–435. [Google Scholar] [CrossRef]
- Cermeño, M.; Stack, J.; Tobin, P.R.; O’Keeffe, M.B.; Harnedy, P.A.; Stengel, D.B.; FitzGerald, R.J. Peptide identification from a Porphyra dioica protein hydrolysate with antioxidant, angiotensin converting enzyme and dipeptidyl peptidase IV inhibitory activities. Food Funct. 2019, 10, 3421–3429. [Google Scholar] [CrossRef] [PubMed]
- Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Purification and identification of dipeptidyl peptidase (DPP) IV inhibitory peptides from the macroalga Palmaria palmata. Food Chem. 2015, 172, 400–406. [Google Scholar] [CrossRef]
- Mune Mune, M.A.; Minka, S.R.; Henle, T. Investigation on antioxidant, angiotensin converting enzyme and dipeptidyl peptidase IV inhibitory activity of Bambara bean protein hydrolysates. Food Chem. 2018, 250, 162–169. [Google Scholar] [CrossRef]
- Rudolph, S.; Lunow, D.; Kaiser, S.; Henle, T. Identification and quantification of ACE-inhibiting peptides in enzymatic hydrolysates of plant proteins. Food Chem. 2017, 224, 19–25. [Google Scholar] [CrossRef]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural requirements of angiotensin I-converting enzyme inhibitory peptides: Quantitative structure−activity relationship study of di-and tripeptides. J. Agri. Food Chem. 2006, 54, 732–738. [Google Scholar] [CrossRef]
- Sumikawa, K.; Takei, K.; Kumagai, Y.; Shimizu, T.; Yasui, H.; Kishimura, H. In silico analysis of ACE inhibitory peptides from chloroplast proteins of red alga Grateloupia asiatica. Mar. Biotechnol. 2020, 22, 391–402. [Google Scholar] [CrossRef]
- Apt, K.E.; Metzner, S.; Grossman, A.R. The γ subunits of phycoerythrin from a red alga: Position in phycobilisomes and sequence characterization. J. Phycol. 2001, 37, 64–70. [Google Scholar] [CrossRef]
- Vecchi, B.; Añon, M.C. ACE inhibitory tetrapeptides from Amaranthus hypochondriacus 11S globulin. Phytochemistry 2009, 70, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Liao, W.; Wu, J. Molecular interactions, bioavailability, and cellular mechanisms of angiotensin-converting enzyme inhibitory peptides. J. Food Biochem. 2019, 43, e12572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage e T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Cheung, H.S.; Cushman, D.W. Inhibition of homogeneous angiotensin-converting enzyme of rabbit lung by synthetic venom peptides of Bothrops jararaca. Biochim. Biophys. Acta (BBA)-Enzymol. 1973, 293, 451–463. [Google Scholar] [CrossRef]
- Hatanaka, T.; Inoue, Y.; Arima, J.; Kumagai, Y.; Usuki, H.; Kawakami, K.; Kimura, M.; Mukaihara, T. Production of dipeptidyl peptidase IV inhibitory peptides from defatted rice bran. Food Chem. 2012, 134, 797–802. [Google Scholar] [CrossRef]
- Sharma, O.P.; Bhat, T.K. DPPH antioxidant assay revisited. Food Chem. 2009, 113, 1202–1205. [Google Scholar] [CrossRef]
- Wilson, P.J.; Grewal, S.; McFadden, T.; Chambers, R.C.; White, B.N. Mitochondrial DNA extracted from eastern North American wolves killed in the 1800s is not of gray wolf origin. Canadian J. Zool. 2003, 81, 936–940. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucl. Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- DeLano, W.L.; Bromberg, S. PyMOL User’s Guide; DeLano Scientific LLC: South San Francisco, CA, USA, 2004. [Google Scholar]
- Laskowski, R.A. PDBsum new things. Nucl. Acids Res. 2009, 37, D355–D359. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Secondary Structure a | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sequence | Occurrence | Position | IC50 (µM) | Proteins | [M+H]+ | Solubility b | SAS in nm2 c | Peptide Ranker Score d | ||
| Detected | IDHY | 1 | 86–89 | - | PE-α | 547.25 | Good | 0.381 | 0.285 | Helix |
| LRY | 1 | 90–92 | 5.1 | PE-β | 451.26 | Good | 0.486 | 0.512 | Helix | |
| LVVER | 1 | 159–163 | - | RuBisCo-L | 615.35 | Good | 0.316 | 0.044 | Helix | |
| Potential e | AR | 1/1/1 | 36–37, 56–57, 15–16 | 96 | PE-α/ PC-β/ APC-α | 245.14 | Good | 0.125 0.700 0.865 | 0.394 | Helix Helix - |
| VR | 1/1 | 164–165, 38–39 | 52.8 | PE-γ/ APC-β | 273.17 | Good | 0.579 1.091 | 0.115 | Helix Helix | |
| LEE | 1 | 115–117 | 100 | PC-α | 389.17 | Good | 0.809 | 0.0350 | Helix | |
| FQ | 1 | 59–60 | 51.23 | APC-γ | 293.13 | Poor | 0.843 | 0.916 | Helix | |
| IW | 1 | 285–286 | 4.7 | RuBisCo-L | 317.16 | Poor | 0.453 | 0.944 | Helix | |
| VK | 1 | 134–135 | 13.0 | RuBisCo-L | 245.16 | Good | 0.662 | 0.033 | Beta strand | |
| PE | PC | APC | RuBisCo | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| α | β | γ | α | β | α | β | γ | L | S | |
| Secondary structure descriptors b | ||||||||||
| helices | 10 (74.4%) c | 10 (74.9%) | 14 (62.1%) | 9 (76.6%) | 10 (71.2%) | 9 (78.1%) | 7 (78.3%) | 9 (77.5%) | 25 (42.2%) | 3 (20.3%) |
| beta hairpins | - | - | - | - | - | - | - | - | 2 | 2 |
| beta sheets | - | - | - | - | - | - | - | - | 3 | 2 |
| beta strands | - | - | - | - | - | - | - | - | 15 (14.8%) | 4 (40.5%) |
| beta bulges | - | - | - | - | - | - | - | - | 3 | 2 |
| Helix–helix interactions | 18 | 17 | 14 | 17 | 18 | 19 | 13 | 20 | 22 | - |
| beta turns | 5 | 4 | 13 | 2 | 4 | 6 | 7 | 6 | 27 | 5 |
| gamma turns | 1 | - | 1 | - | - | - | - | - | 3 | - |
| Ligand | Number Clusters | Affinity (kcal/mol) a | RMSDi in A | Number of Bonds with Zn2+ | Hydrogen Bonds b | Number of Non-Bonded Contacts | ||
|---|---|---|---|---|---|---|---|---|
| Number | Residues | Length (Å) | ||||||
| IDHY | 10 | −9.50 | 0.0 | 2 | 7 | TYR523 GLU403 GLU403 GLU384 ALA356 SER355 ALA354 | 3.323 3.189 2.568 2.926 2.847 2.958 3.067 | 102 |
| LVVER | 10 | −6.9 | 0.0 | 0 | 6 | SER517 SER516 GLU143 GLU143 ARG124 ASP121 | 2.681 2.994 2.998 2.941 2.741 2.619 | 62 |
| Lisinopril | - | - | - | 2 | 8 | TYR523 TYR520 HIS513 LYS511 HIS387 ALA354 HIS353 HIS353 | 2.775 2.556 3.109 2.934 3.327 2.917 2.760 3.241 | 48 |
| Captopril | - | - | - | 1 | 5 | TYR520 HIS513 LYS511 HIS353 GLN281 | 2.657 2.694 2.730 2.542 3.074 | 36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mune Mune, M.A.; Miyabe, Y.; Shimizu, T.; Matsui, W.; Kumagai, Y.; Kishimura, H. Characterisation of Bioactive Peptides from Red Alga Gracilariopsis chorda. Mar. Drugs 2023, 21, 49. https://doi.org/10.3390/md21010049
Mune Mune MA, Miyabe Y, Shimizu T, Matsui W, Kumagai Y, Kishimura H. Characterisation of Bioactive Peptides from Red Alga Gracilariopsis chorda. Marine Drugs. 2023; 21(1):49. https://doi.org/10.3390/md21010049
Chicago/Turabian StyleMune Mune, Martin Alain, Yoshikatsu Miyabe, Takeshi Shimizu, Wataru Matsui, Yuya Kumagai, and Hideki Kishimura. 2023. "Characterisation of Bioactive Peptides from Red Alga Gracilariopsis chorda" Marine Drugs 21, no. 1: 49. https://doi.org/10.3390/md21010049
APA StyleMune Mune, M. A., Miyabe, Y., Shimizu, T., Matsui, W., Kumagai, Y., & Kishimura, H. (2023). Characterisation of Bioactive Peptides from Red Alga Gracilariopsis chorda. Marine Drugs, 21(1), 49. https://doi.org/10.3390/md21010049