
Structural Diversity and Biological Activity of Cyanopeptolins Produced by Nostoc edaphicum CCNP1411

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:
1. Introduction
2. Results and Discussion
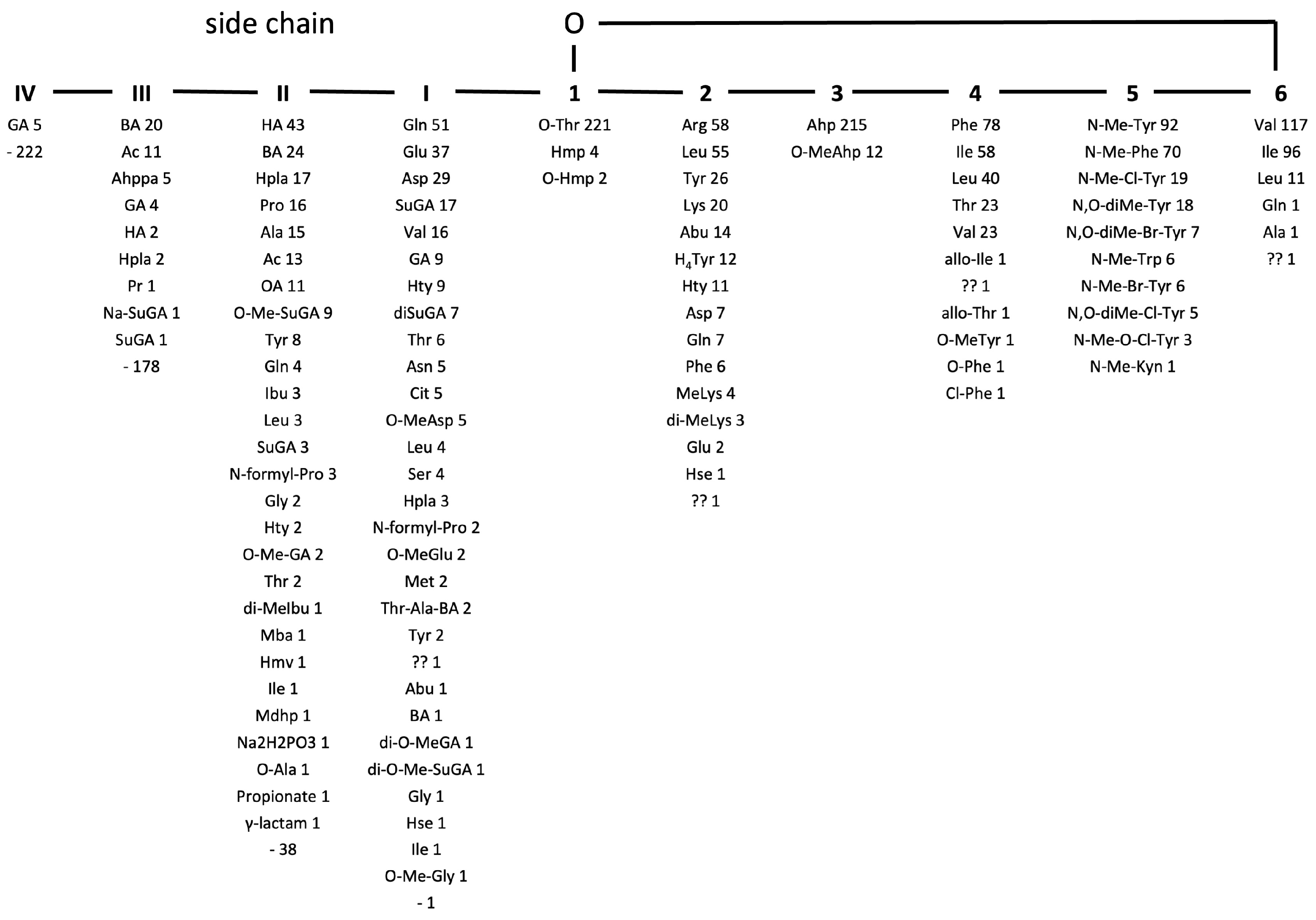
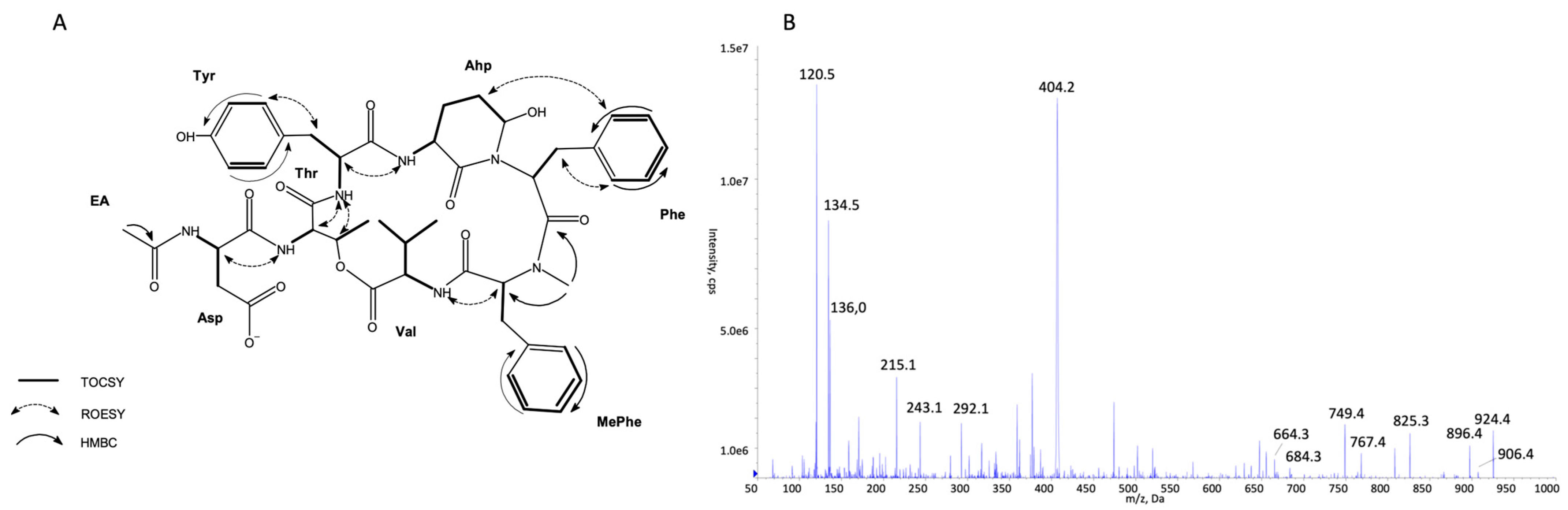
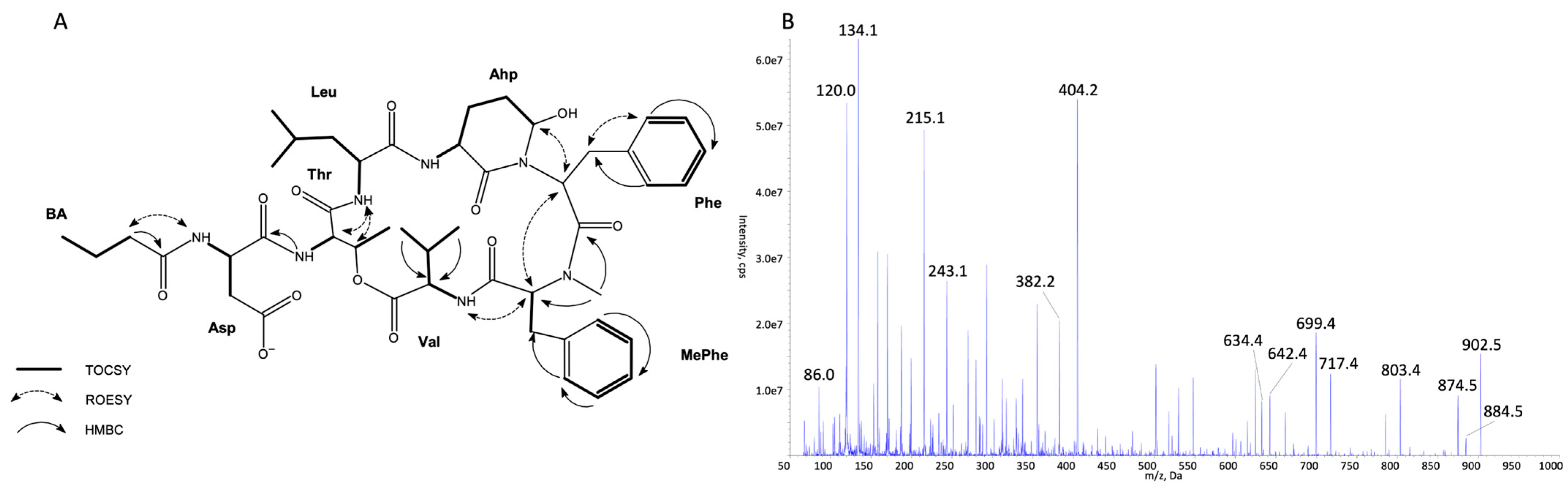
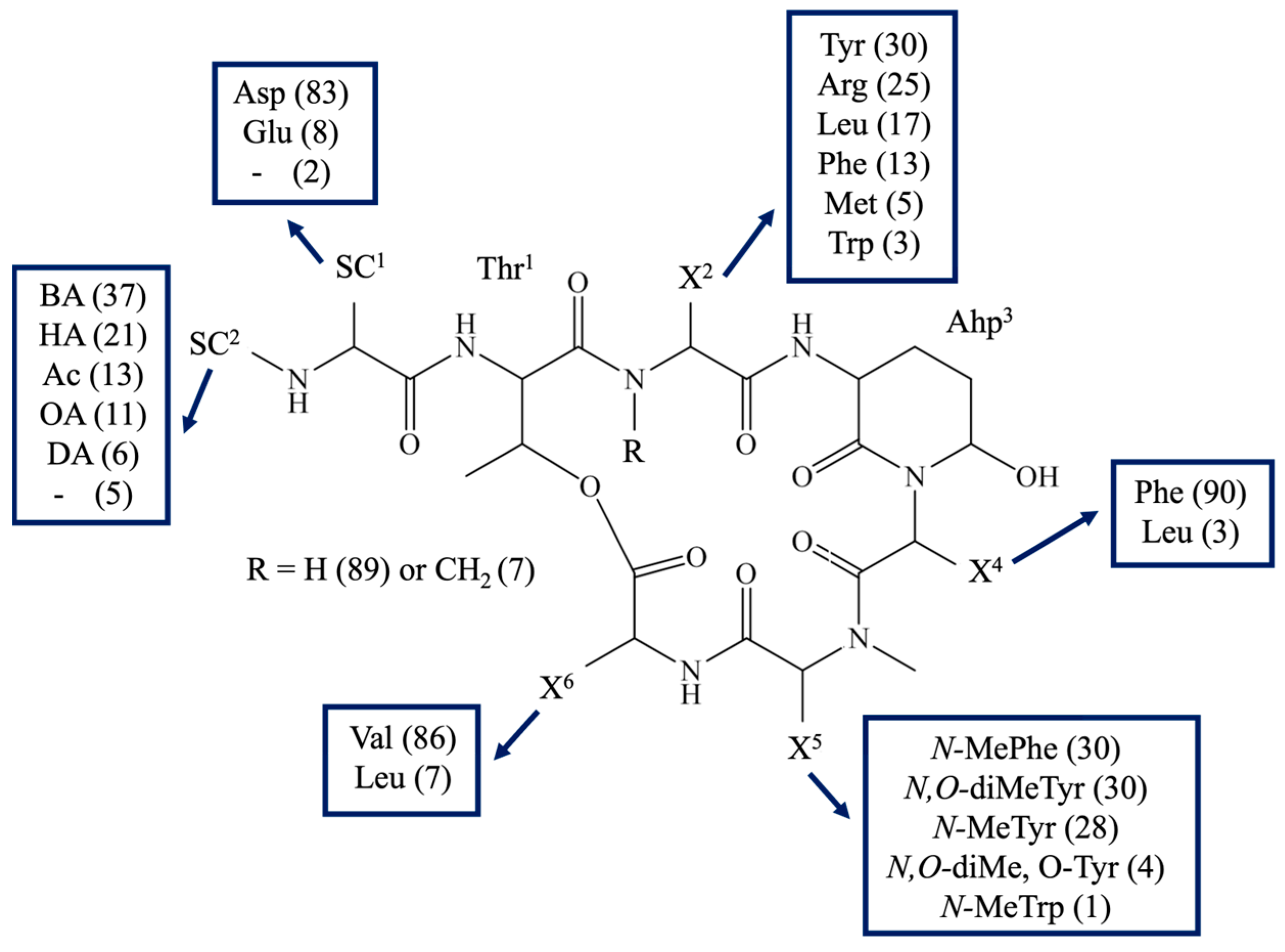
2.1. Identification of CP Structures
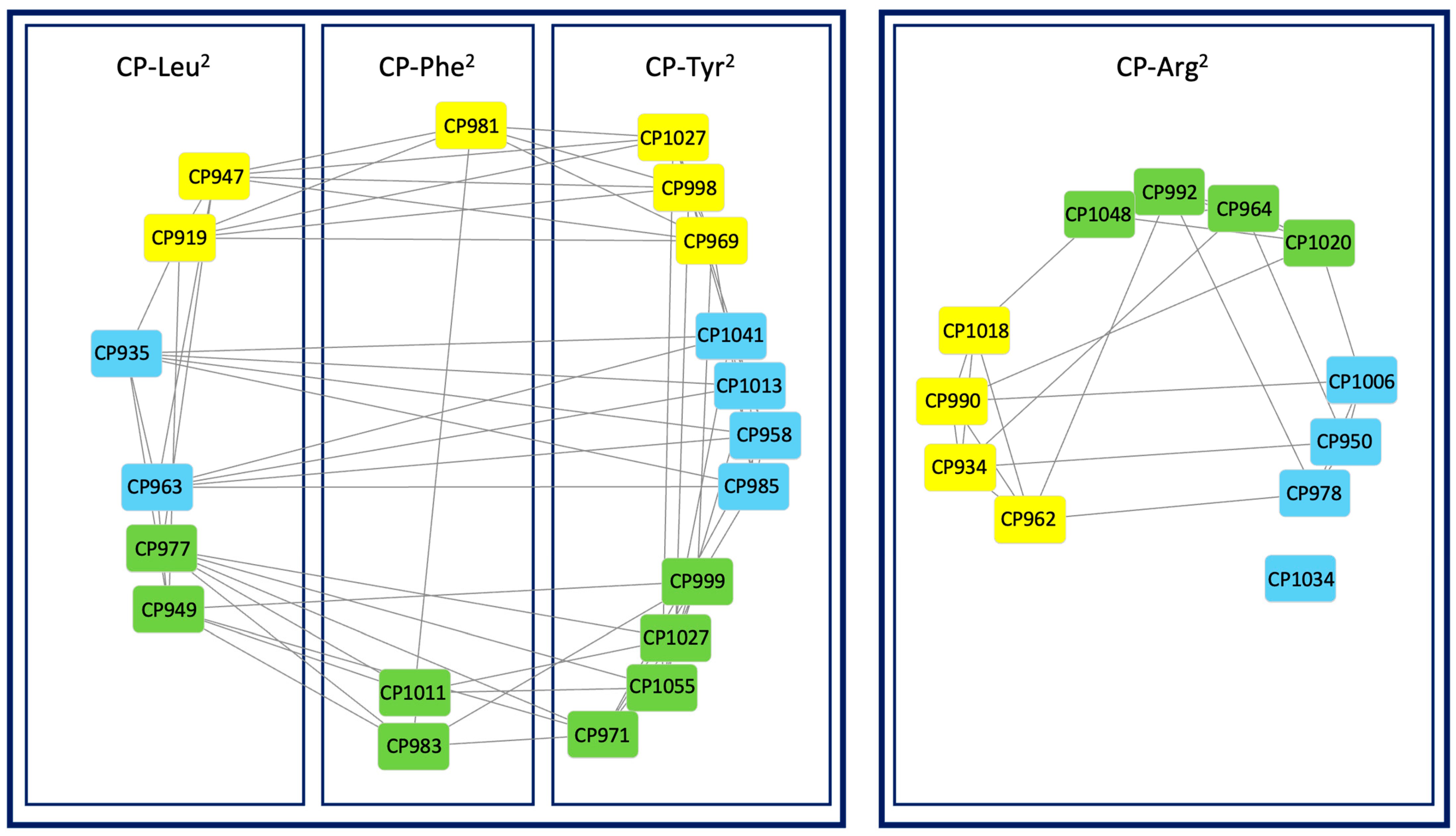
2.2. Molecular Networking of Cyanopeptolins
2.3. Enzymatic Assay
2.4. MTT Assay
3. Materials and Methods
3.1. Extraction and Isolation of Cyanopeptolins
3.2. LC-MS/MS Analysis
3.3. LC-HRMS Analysis
3.4. Molecular Networking
3.5. NMR Analysis
3.6. Enzymatic Assays
3.7. MTT Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Jones, M.R.; Pinto, M.; Torres, M.A.; Dörr, F.; Mazur-Marzec, H.; Szubert, K.; Tartaglione, L.; Dell’Aversano, C.; Miles, C.O.; Beach, D.G.; et al. CyanoMetDB, a comprehensive public database of secondary metabolites from cyanobacteria. Water Res. 2021, 196, 117017. [Google Scholar] [CrossRef]
- Janssen, E.M.-L.; Jones, M.R.; Pinto, E.; Dörr, F.; Torres, M.A.; Rios, J.F.; Mazur-Marzec, H.; Szubert, K.; Konkel, R.; Tartaglione, L.; et al. CyanoMetDB|Comprehensive Database of Secondary Metabolites from Cyanobacteria (NORMAN-SLE-S75.0.2.0); Zenodo: Geneva, Switzerland; CERN: Meyrin, Switzerland, 2023. [Google Scholar] [CrossRef]
- Martin, C.; Oberer, L.; Ino, T.; König, W.A.; Busch, M.; Weckesse, J. Cyanopeptolins, new depsipeptides from the cyanobacterium Microcystis sp. PCC 7806. J. Antibiot. 1993, 46, 1550–1556. [Google Scholar] [CrossRef]
- Ozaki, K.; Iwasaki, A.; Suenaga, K.; Teruya, T. Kyanamide, a new Ahp-containing depsipeptide from marine cyanobacterium Caldora penicillate. Tetrahedron 2019, 75, 3383–3386. [Google Scholar] [CrossRef]
- Fujii, K.; Sivonen, K.; Nakano, T.; Harada, K.-i. Structural elucidation of cyanobacterial peptides encoded by peptide synthetase gene in Anabaena species. Tetrahedron 2002, 58, 6863–6871. [Google Scholar] [CrossRef]
- Gunesekera, S.P.; Miller, M.W.; Kwan, J.C.; Luesch, H.; Paul, V.J. Molassamide, a Depsipeptide Serine Protease Inhibitor from the Marine Cyanobacterium Dichothrix utahensis. J. Nat. Prod. 2010, 73, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Bonjouklian, R.; Smitka, T.A.; Hunt, A.H.; Occolowitz, J.L.; Perun, T.J., Jr.; Doolin, L.; Stevenson, S.; Knauss, L.; Wijayaratne, R.; Szewczyk, S.; et al. A90720A, A Serine Protease Inhibitor Isolated From A Terrestrial Blue-Green Alga Microchaete loktakensis. Tetrahedron 1996, 55, 395–404. [Google Scholar] [CrossRef]
- Mazur-Marzec, H.; Fidor, A.; Cegłowska, M.; Wieczerzak, E.; Kropidłowska, M.; Goua, M.; Macaskill, J.; Edwards, C. Cyanopeptolin with Trypsin and Chymotrypsin Inhibitory Activity from the Cyanobacterium Nostoc edaphicum CCNP1411. Mar. Drugs 2018, 16, 220. [Google Scholar] [CrossRef] [PubMed]
- Okino, T.; Qi, S.; Matsuda, H.; Murakami, M.; Yamaguchi, K. Nostopeptins A and B, Elastase Inhibitors from the Cyanobacterium Nostoc minutum. J. Nat. Prod. 1997, 60, 158–161. [Google Scholar] [CrossRef]
- Ploutno, A.; Shoshan, M.; Carmeli, S. Three Novel Protease Inhibitors from a Natural Bloom of the Cyanobacterium Microcystis aeruginosa. J. Nat. Prod. 2002, 65, 973–978. [Google Scholar] [CrossRef]
- Kaya, K.; Sano, T.; Beattie, K.A.; Codd, G.A. Nostocyclin, A Novel 3-Amino-6-hydroxy-2-piperidone-containing Cyclic Depsipeptide from the Cyanobacterium Nostoc sp. Tetrahedron Lett. 1996, 37, 6725–6728. [Google Scholar] [CrossRef]
- Ploutno, A.; Carmeli, S. Modified peptides from a water bloom of the cyanobacterium Nostoc sp. Tetrahedron 2002, 58, 9949–9957. [Google Scholar] [CrossRef]
- Matern, U.; Oberer, L.; Falchetto, R.A.; Erhard, M.; Köning, W.A.; Herdman, M.; Weckesser, J. Scyptolin A and B, cyclic depsipeptides from axenic cultures of Scytonema hofmanni PCC 7110. Phytochemistry 2001, 58, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Matern, U.; Oberer, L.; Erhard, M.; Herdman, M.; Weckesser, J. Hofmannolin, a cyanopeptolin from Scytonema hofmanni PCC 7110. Phytochemistry 2003, 64, 1061–1067. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.-S.; Krunic, A.; Orjala, J. Stigonemapeptin, an Ahp-containing Depsipeptide with Elastase Inhibitory Activity from the Bloom-Forming Freshwater Cyanobacterium Stigonema sp. J. Nat. Prod. 2012, 75, 807–811. [Google Scholar] [CrossRef]
- Choi, H.; Oh, S.K.; Yih, W.; Chin, J.; Kang, H.; Tho, J.-R. Cyanopeptoline CB071: A Cyclic Depsipeptide Isolated from the Freshwater Cyanobacterium Aphanocapsa sp. Chem. Pharm. Bull. 2008, 56, 1191–1193. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, L.; Zhou, Y.; Guo, Y.-L. A novel pyrimidine-based stable-isotope labeling reagent and its application to quantitative analysis using matrix-assisted laser desorption/ionization mass spectrometry. J. Mass Spectrom. 2007, 42, 1514–1521. [Google Scholar] [CrossRef]
- Taori, K.; Paul, V.J.; Luesch, H. Kempopeptins A and B, Serine Protease Inhibitors with Different Selectivity Profiles from a Marine Cyanobacterium, Lyngby sp. J. Nat. Prod. 2008, 71, 1625–1629. [Google Scholar] [CrossRef]
- Rubio, B.K.; Parrish, S.M.; Yoshida, W.; Schupp, P.J.; Schils, T.; Williams, P.G. Depsipeptides from a Guamanian marine cyanobacterium Lyngbya bouillonii, with selective inhibition of serine proteases. Tetrahedron Lett. 2010, 51, 6718–6721. [Google Scholar] [CrossRef]
- Kisugi, T.; Okino, T. Micropeptins from the Freshwater Cyanobacterium Microcystis aeruginosa (NIES-100). J. Nat. Prod. 2009, 72, 777–781. [Google Scholar] [CrossRef]
- Adiv, S.; Aharonv-Nadborny, R.; Carmeli, S. Micropeptins from Microcystis aeruginosa collected in Dalton reservoir, Israel. Tetrahedron 2010, 66, 7429–7436. [Google Scholar] [CrossRef]
- Elkobi-Peer, S.; Carmeli, S. New Prenylated Aeruginosin, Microphycin, Anabaenopeptin and Micropeptin Analogues from a Microcystis Bloom Material Collected in Kibbutz Kfar Blum, Israel. Mar. Drugs 2015, 13, 2347–2375. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.-i.; Mayumi, T.; Shimada, T.; Suzuki, M. Occurrence of Four Depsipeptides, Aeruginopeptins, Together with Microcystins from Toxic Cyanobacteria. Tetrahedron Lett. 1993, 34, 6091–6094. [Google Scholar] [CrossRef]
- Beresovsky, D.; Hadas, O.; Livne, A.; Sukenik, A.; Kaplan, A.; Carmeli, S. Toxins and Biologically Active Secondary Metabolites of Microcystis sp. isolated from Lake Kinneret. Isr. J. Chem. 2006, 46, 79–87. [Google Scholar] [CrossRef]
- Okumura, H.S.; Philmus, B.; Portman, C.; Kemscheidt, T.K. Homotyrosine-containing Cyanopeptolin 880 & 960 and Anabaenopeptins 908 & 915 from Planktothrix agardhii CYA 126/8. J. Nat. Prod. 2009, 72, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Plaza, A.; Bewley, C.A. Largamides A-H, Unusual Cyclic Peptides from the Marine Cyanobacterium Oscillatoria sp. J. Org. Chem. 2006, 71, 6898–6907. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Sivonen, K.; Naganawa, E.; Harada, K.-i. Non-Toxic Peptides from Toxic Cyanobacteria, Oscillatoria agardhii. Tetrahedron 2000, 56, 725–733. [Google Scholar] [CrossRef]
- Itou, Y.; Ishida, L.; Shin, H.J.; Murakami, M. Oscillapeptins A to F, Serine Protease Inhibitors from the Three Strains of Oscillatoria agarhii. Tetrahedron 1999, 55, 6871–6882. [Google Scholar] [CrossRef]
- Harada, K.-i. Production of secondary metabolites by freshwater cyanobacteria. Chem. Pharm. Bull. 2004, 52, 889–899. [Google Scholar] [CrossRef]
- Williams, P.G.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Tasipeptins A and B: New Cytotoxic Depsipeptides from the Marine Cyanobacterium Symploca sp. J. Nat. Prod. 2003, 66, 620–624. [Google Scholar] [CrossRef]
- Linington, R.G.; Edwards, D.J.; Shuman, C.F.; McPhail, K.L.; Matainaho, T.; Gerwick, W.H. Symplocamide A, a Potent Cytotoxin and Chymotrypsin Inhibitor from the Marine Cyanobacterium Symploca sp. J. Nat. Prod. 2008, 71, 22–27. [Google Scholar] [CrossRef]
- Pettit, G.R.; Kamano, Y.; Herald, C.L.; Dufresne, C.; Cerny, R.L.; Herald, D.L.; Schmidt, J.M.; Kizu, H. Isolation and Structure of the Cytostatic Depsipeptide Dolastatin 13 from the Sea Hare Dolabella auricularia. J. Am. Chem. Soc. 1989, 111, 5015–5017. [Google Scholar] [CrossRef]
- Viehrig, K.; Surup, F.; Harmrolfs, K.; Jansen, R.; Kunze, B.; Kunze, B.; Müller, R. Concerted Action of P450 Plus Helper Protein To Form the Amino-hydroxy-piperidone Moiety of the Potent Protease Inhibitor Crocapeptin. J. Am. Chem. Soc. 2013, 135, 16885–16894. [Google Scholar] [CrossRef] [PubMed]
- Kodani, S.; Komaki, H.; Hemmi, H.; Miyake, Y.; Kaweewan, I.; Dohra, H. Streptopeptolin, a Cyanopeptolin-Type Peptide from Streptomyces olivochromogenes. ACS Omega 2018, 3, 8104–8110. [Google Scholar] [CrossRef]
- Gademann, K.; Portmann, C.; Blom, J.F.; Zeder, M.; Jüttner, F. Multiple Toxin Production in the Cyanobacterium Microcystis: Isolation of the Toxic Protease Inhibitor Cyanopeptolin 1020. J. Nat. Prod. 2010, 73, 980–984. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.; Smitka, T.A.; Bonjouklian, R.; Clardy, J. Atomic structure of the trypsin-A90720A complex: A unified approach to structure and function. Chem. Biol. 1994, 1, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Okino, T.; Murakami, M.; Haraguchi, R.; Munekata, H.; Matsuda, H.; Yamaguchi, K. Micropeptins A and B, Plasmin and Trypsin Inhibitors from the Blue-Green Alga Microcystis aeruginosa. Tetrahedron Lett. 1993, 34, 8131–8134. [Google Scholar] [CrossRef]
- Kodani, S.; Suzuki, S.; Ishida, K.; Murakami, M. Five new cyanobacterial peptides from water bloom materials of lake Teganuna (Japan). FEMS Microbiol. Lett. 1999, 178, 343–348. [Google Scholar] [CrossRef]
- Okano, T.; Sano, T.; Kaya, K. Micropeptin T-20, A Novel Phospate-containing Cyclic Depsipeptide from the Cyanobacterium Microcystis aeruginosa. Tetrahedron Lett. 1999, 40, 2379–2382. [Google Scholar] [CrossRef]
- Von Elert, E.; Oberer, L.; Merkel, P.; Huhn, T.; Blom, J.F. Cyanopeptolin 954, a Chlorine-Containing Chymotrypsin Inhibitor of Microcystis aeruginosa NIVA Cya 43. J. Nat. Prod. 2005, 68, 1324–1327. [Google Scholar] [CrossRef]
- Lifshits, M.; Carmeli, S. Metabolites of Microcystis aeruginosa Bloom Material from Lake Kinneret, Israel. J. Nat. Prod. 2012, 75, 209–219. [Google Scholar] [CrossRef]
- Zafrir-Ilan, E.; Carmeli, S. Eight novel serine proteases inhibitors from a water bloom of the cyanobacterium Microcystis sp. Tetrahedron 2010, 66, 9194–9202. [Google Scholar] [CrossRef]
- Murakami, M.; Kodani, S.; Ishida, K.; Matsuda, H.; Yamaguchi, K. Micropeptin 103, a Chymotrypsin Inhibitor from the Cyanobacterium Microcystis viridis (NIES-103). Tetrahedron Lett. 1997, 38, 3035–3038. [Google Scholar] [CrossRef]
- Matthew, S.; Ross, C.; Rocca, J.R.; Paul, V.J.; Luesch, H. Lyngbyastatin 4, a Dolastatin 13 Analogue with Elastase and Chymotrypsin Inhibitory Activity from the Marine Cyanobacterium Lyngbya confervoides. J. Nat. Prod. 2007, 70, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Grach-Pogrebinsky, O.; Sedmak, B.; Carmeli, S. Protease inhibitors from a Slovenian Lake Bled toxic waterbloom of the cyanobacterium Planktothrix rubescens. Tetrahedron 2003, 59, 8329–8336. [Google Scholar] [CrossRef]
- Shin, H.J.; Murakami, M.; Matsuda, H.; Ishida, K.; Yamaguchi, K. Oscillapeptin, an Elastase and Chymotrypsin Inhibitor from the Cyanobacterium Oscillatoria agardhii (NIES-204). Tetrahedron Lett. 1995, 36, 5235–5238. [Google Scholar] [CrossRef]
- Zafrir, E.; Carmeli, S. Micropeptins from an Israeli Fishpond Water Bloom of the Cyanobacterium Microcystis sp. J. Nat. Prod. 2010, 73, 352–358. [Google Scholar] [CrossRef]
- Jakobi, C.; Kenneth, L.R.; Neuber, R.; Mez, K.; Weckesser, J. Cyanopeptolin SS, a disulphated depsipeptides from a water bloom: Structural elucidation and biological activities. Phycologia 1996, 35, 111–116. [Google Scholar] [CrossRef]
- Mehner, C.; Müller, D.; Kehraus, S.; Hautmann, S.; Gütschow, M.; Köning, G.M. New Peptolides from the Cyanobacterium Nostoc insulare as Selective and Potent Inhibitors of Human Leukocyte Elastase. ChemBioChem 2008, 9, 2692–2703. [Google Scholar] [CrossRef]
- Egli, C.M.; Natumi, R.S.; Jones, M.R.; Janssen, E.M.-L. Inhibition of Extracellular Enzymes Exposed to Cyanopeptides. Environ. Chem. 2020, 74, 122–128. [Google Scholar] [CrossRef]
- Luo, D.; Chen, Q.-Y.; Luesch, H. Total Synthesis of the Potent Marine-Derived Elastase Inhibitor Lyngbyastatin 7 and in Vitro Biological Evaluation in Model Systems for Pulmonary Diseases. J. Org. Chem. 2016, 81, 532–544. [Google Scholar] [CrossRef]
- Al-Awadhi, F.H.; Paul, V.J.; Luesch, H. Structural Diversity and Anticancer Activity of Marine-Derived Elastase Inhibitors: Key Features and Mechanisms Mediating the Antimetastatic Effects in Invasive Breast Cancer. ChemBioChem 2018, 19, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Mooberry, S.L.; Leal, R.M.; Tinley, T.L.; Luesch, H.; Moore, R.E.; Corbett, T.H. The molecular pharmacology of symplostatin 1: A new antimiotic dolastatin 10 analog. Int. J. Cancer 2003, 204, 4. [Google Scholar] [CrossRef] [PubMed]
- Zainuddin, E.Z.; Mentel, R.; Wray, V.; Jansen, R.; Nimtz, M.; Lalk, M.; Mundt, S. Cyclic Depsipeptides, Ichthyopeptins A and B, from Microcystis ichthyoblabe. J. Nat. Prod. 2007, 70, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.-i.; Mayumi, T.; Shimada, T.; Fujii, K.; Kondo, F.; Park, H.-d.; Watanabe, M.F. Co-production of Microcystins and Aeruginopeptins by Natural Cyanobacterial Bloom. Environ. Toxicol. 2001, 16, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Gallegos, D.A.; Sauri, J.; Cohen, R.D.; Wan, X.; Videau, P.; Vallota-Eastkaj, A.O.; Shaala, L.A.; Youssef, D.T.A.; Williamson, R.T.; Martin, G.E.; et al. Jizanpeptins, Cyanobacterial Protease Inhibitors from a Symploca sp. Cyanobacterium Collected in the Ted Sea. J. Nat. Prod. 2018, 81, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Kwan, J.; Taori, K.; Paul, V.; Luesch, H. Lyngbyastatins 8–10, elastase inhibitors with cyclic depsipeptide scaffolds isolated from the marine cyanobacterium Lyngbya semiplena. Mar. Drugs 2009, 7, 528–538. [Google Scholar] [CrossRef]
- Taori, K.; Matthew, S.; Rocca, J.R.; Paul, V.J.; Luesch, H. Lyngbyastatins 5–7, Potent Elastase Inhibitors from Floridian Marine Cyanobacteria, Lyngbya spp. J. Nat. Prod. 2007, 70, 1593–1600. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Painuly, P.; Young, K.A.; Yang, X.; Shimizu, Y. Microcystilide A: A Novel Cell-Differentiation-Promoting Depsipeptide from Microcystis aeruginosa NO-15-1840. J. Am. Chem. Soc. 1993, 115, 11046–11047. [Google Scholar] [CrossRef]
- Isaacs, J.D.; Strangman, W.K.; Barbera, A.E.; Mallim, M.A.; McIver, M.R.; Wright, J.L.C. Microcystins and two new micropeptin cyanopeptides produced by unprecedented Microcystis aeruginosa blooms in North Carolina’s Cape Fear River. Harmful Algae 2014, 31, 82–86. [Google Scholar] [CrossRef]
- Ishida, K.; Murakami, M.; Matsuda, H.; Yamaguchi, K. Micropeptin 90, a Plasmin and Trypsin Inhibitor from the Blue0Green Alga Microcystis aeruginosa (NIES-90). Tetrahedron Lett. 1995, 36, 3535–3538. [Google Scholar] [CrossRef]
- Ishida, K.; Matsuda, H.; Murakami, M.; Yamaguchi, K. Micropeptins 478-A and -B, Plasmin Inhibitors from the Cyanobacterium Microcystis aeruginosa. J. Nat. Prod. 1997, 60, 184–187. [Google Scholar] [CrossRef]
- Ishida, K.; Matsuda, H.; Murakami, M. Micropeptins 88-A to 88-F, chymotrypsin inhibitors from the cyanobacterium Microcystis aeruginosa (NIES-88). Tetrahedron 1998, 54, 5545–5556. [Google Scholar] [CrossRef]
- Yamaki, H.; Sitachitta, N.; Sano, T.; Kaya, K. Two New Chymotrypsin Inhibitors Isolated from the Cyanobacterium Microcystis aeruginosa NIES-88. J. Nat. Prod. 2005, 68, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, K.; Gomes, A.; Calado, L.; Yasui, G.; Assis, D.; Henry, T.; Fonseca, A.; Pinto, E. Toxicity of Cyanopeptides from Two Microcystis Strains on Larval Development of Astyanax altiparanae. Toxins 2019, 11, 220. [Google Scholar] [CrossRef] [PubMed]
- Strangman, W.K.; Stewart, A.K.; Herring, M.C.; Wright, J.L.C. Identification of the new chymotrypsin inhibitor micropeptin 996 by metabolomics-guided analysis. Tetrahedron Lett. 2018, 59, 935–937. [Google Scholar] [CrossRef]
- Blom, J.F.; Bister, B.; Bischoff, D.; Nicholson, G.; Jung, G.; Süssmuth, R.D.; Jüttner, F. Oscillapeptin J, a New Grazer Toxin of the Freshwater Cyanobacterium Planktothrix rubescens. J. Nat. Prod. 2003, 66, 431–434. [Google Scholar] [CrossRef]
- Matthew, S.; Ross, C.; Paul, V.J.; Luesch, H. Pompanopeptins A and B, new cyclic peptides from the marine cyanobacterium Lyngbya confervoides. Tetrahedron 2008, 64, 4081–4089. [Google Scholar] [CrossRef]
- Nogle, L.M.; Williamson, T.; Gerwick, W.H. Somamides A and B, Two New Depsipeptide Analogues of Dolastatin 13 from a Fijian Cyanobacterial Assemblage of Lyngbya majuscule and Schizothrix Species. J. Nat. Prod. 2001, 64, 716–719. [Google Scholar] [CrossRef]
- Walsh, C.T.; O’Brien, R.V.; Khosla, C. Nonproteinogenic amino acid building blocks for nonribosomal peptide and hybrid polyketide scaffolds. Angew. Chem. 2013, 52, 7098–7124. [Google Scholar] [CrossRef]
- Fisch, K.M. Biosynthesis of natural products by microbial iterative hybrid PKS-NRPS. RSC Adv. 2013, 3, 18228–18247. [Google Scholar] [CrossRef]
- Czarnecki, O.; Henning, M.; Lippert, I.; Welker, M. Identification of peptide metabolites of Microcystis (Cyanobacteria) that inhibit trypsin-like activity in planktonic herbivorous Daphnia (Cladocera). Environ. Microbiol. 2006, 8, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Gesner-Apter, S.; Carmeli, S. Protease Inhibitors from a Water Bloom of the Cyanobacterium Microcystis aeruginosa. J. Nat. Prod. 2009, 72, 1429–1436. [Google Scholar] [CrossRef]
- Banker, R.; Carmeli, S. Inhibitors of Serine Proteases from a Waterbloom of the Cyanobacterium Microcystis sp. Tetrahedron 1999, 55, 10835–10844. [Google Scholar] [CrossRef]
- Vegman, M.; Carmeli, S. Eight micropeptins from a Microcystis spp. Bloom collected from a fishpond near Kibbutz Lehavot HaBashan, Israel. Tetrahedron 2013, 69, 10108–10115. [Google Scholar] [CrossRef]
- Welker, M.; von Döhren, H. Cyanobacterial peptides—Nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef] [PubMed]
- Jakobi, C.; Oberer, L.; Quiquerez, C.; König, W.A.; Weckesser, J. Cyanopeptolin S, a sulfate-containing depsipeptides from a water bloom of Microcystis sp. FEMS Microbiol. Lett. 1995, 129, 129–134. [Google Scholar] [CrossRef]
- Reshef, V.; Carmeli, S. Protease inhibitors from a water bloom of the cyanobacterium Microcystis aeruginosa. Tetrahedron 2001, 57, 2885–2894. [Google Scholar] [CrossRef]
- Lodin-Friedman, A.; Carmeli, S. Metabolites from Microcystis aeruginosa Bloom Material Collected at a Water Reservoir near Kibbutz Hafetz Haim, Israel. J. Nat. Prod. 2013, 76, 1196–1200. [Google Scholar] [CrossRef]
- Bister, B.; Keller, S.; Baumann, H.I.; Nicholson, G.; Weist, S.; Jung, G.; Süssmuh, R.D.; Jüttner, F. Cyanopeptolin 963A, a Chymotrypsin Inhibitor of Microcystis PCC 7806. J. Nat. Prod. 2004, 67, 1755–1757. [Google Scholar] [CrossRef]
- Welker, M.; Brunke, M.; Preussel, K.; Lippert, I.; von Döhren, H. Diversity and distribution of Microcystis (Cyanobacteria) oligopeptide chemotypes from natural communities studied by single-colony mass spectrometry. Microbiology 2004, 150, 1785–1796. [Google Scholar] [CrossRef]
- Lifshits, M.; Zafrir-Ilan, E.; Raveh, A.; Carmeli, S. Protease inhibitors from the three fishpond water blooms of Microcysis spp. Tetrahedron 2011, 67, 4017–4024. [Google Scholar] [CrossRef]
- Tooming-Klunderud, A.; Rohrlack, T.; Shalchian-Tabrizi, K.; Kristensen, T.; Jakobsen, K. Structural analysis of non-ribosomal halogenated cyclic peptide and its putative operon from Microcystis: Implications for evolution of cyanopeptolins. Microbiology 2007, 153, 1382–1393. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.; Andreote, A.P.D.; Fiore, M.F.; Dörr, F.A.; Pinto, E. Structural Characterization of New Peptide Variants Produced by Cyanobacteria from the Brazilian Atlantic Coastal Forest Using Liquid Chromatography Coupled to Quadrupole Time-of-Flight Tandem Mass Spectrometry. Mar. Drugs 2015, 13, 3892–3919. [Google Scholar] [CrossRef] [PubMed]
- Bladt, T.T.; Kalifa-Aviv, S.; Larsen, T.O.; Carmeli, S. Micropeptins from Microcystis sp. Collected in Kabul Reservoir, Israel. Tetrahedron 2014, 70, 936–943. [Google Scholar] [CrossRef]
- Welker, M.; Maršálek, B.; Šejnohová, L.; von Döhren, H. Detection and identification of oligopeptides in Microcystis (cyanobacteria) colonies: Toward an understanding of metabolic diversity. Peptides 2006, 27, 2090–2103. [Google Scholar] [CrossRef]
- Lenz, K.A.; Miller, T.R.; Ma, H. Anabaenopeptins and cyanopeptolins induce systemic toxicity effects in a model organism the nematode Caenorhabditis elegans. Chemosphere 2019, 214, 60–69. [Google Scholar] [CrossRef]
- Hasan-Amer, R.; Carmeli, S. Inhibitors of Serine Proteases from a Microcystis sp. Bloom Material Collected from Timurim Reservoir, Israel. Mar. Drugs 2017, 15, 371. [Google Scholar] [CrossRef]
- Rounge, T.; Rohrlack, T.; Tooming-Klunderud, A.; Kristensen, T.; Jakobsen, K. Comparison of cyanopeptolin genes in Planktothrix, Microcystis and Anabaena strains: Evidence for independent evolution within each genus. Appl. Environ. Microbiol. 2007, 73, 7322–7330. [Google Scholar] [CrossRef]
- Bingnan, H.; Gross, H.; McPhail, K.L.; Goeger, D.; Maier, C.S.; Gerwick, W.H. Wewakamide A and Guineamide G, Cyclic Depsipeptides from the Marine Cyanobacteria Lyngbya semiplena and Lyngbya majuscule. J. Microbiol. Biotechnol. 2011, 21, 930–936. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Don Duy, N.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef]
- Nothias, L.-F.; Peras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Erns, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-based molecular networking in the GNPS analysis environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Jiang, S.; Li, J.; Balaprakash, P.; van Lehn, R.C.; Zavala, V.M. Capturing molecular interactions in graph neural networks: A case study in multi-component phase equilibrium. Digit. Discov. 2023, 2, 138–151. [Google Scholar] [CrossRef]
- Taha, H.M.; Aalizadeh, R.; Alygizakis, N.; Antignac, J.-P.; Arp, H.P.H.; Bade, R.; Baker, N.; Belova, L.; Bijlsma, L.; Bolton, E.E.; et al. The NORMAN Suspect List Exchange (NORMAN-SLE): Facilitating European and worldwide collaboration on suspect screening in high resolution mass spectrometry. Environ. Sci. Eur. 2022, 34, 104. [Google Scholar] [CrossRef] [PubMed]
- Van der Hooft, J.J.J.; Mohimani, H.; Bauermeister, A.; Dorrestein, P.C.; Duncan, K.R.; Medema, M.H. Linking genomics and genomics and metabolomics to chart specialized metabolic diversity. Chem. Soc. Rev. 2020, 49, 3297. [Google Scholar] [CrossRef] [PubMed]
- McDonald, K.; Renaud, J.B.; Pick, F.R.; Miller, J.D.; Sumarah, M.W.; McMullin, D.R. Diagnostic Fragmentation Filtering for Cyanopeptolin Detection. Environ. Toxicol. Chem. 2021, 40, 1087–1097. [Google Scholar] [CrossRef]
- Popin, R.V.; Alvarenga, D.O.; Castelo-Branco, R.; Fewer, D.P.; Sivonen, K. Mining of cyanobacterial genomes indicates natural product biosynthetic gene clusters located in conjugative plasmids. Front. Microbiol. 2021, 4, 684565. [Google Scholar] [CrossRef]
- Konkel, R.; Grabski, M.; Cegłowska, M.; Wieczerzak, E.; Węgrzyn, G.; Mazur-Marzec, H. Anabaenopeptins from Nostoc edaphicum CCNP1411. Int. J. Environ. Res. Public Health 2022, 19, 12346. [Google Scholar] [CrossRef]
- Fidor, A.; Grabski, M.; Gawor, J.; Gromadka, R.; Węgrzyn, G.; Mazur-Marzec, H. Nostoc edaphicum CCNP1411 from the Baltic Sea—A new producer of nostocyclopeptides. Mar. Drugs 2020, 18, 442. [Google Scholar] [CrossRef]
- Gathungu, R.M.; Kautz, R.; Kristal, B.S.; Bird, S.S.; Vouros, P. The integration of LC-MS and NMR for the analysis of low molecular weight trace analytes in complex matrices. Mass Spectrom. Rev. 2020, 39, 35–54. [Google Scholar] [CrossRef]
- Johnson, A.R.; Carlson, E.E. Collision-Induced Dissociation Mass Spectrometry: A Powerful Tool for Natural Product Structure Elucidation. Anal. Chem. 2015, 87, 10668–10678. [Google Scholar] [CrossRef]
- Ma, X. Recent Advances in Mass Spectrometry-Based Structural Elucidation Techniques. Molecules 2022, 27, 6466. [Google Scholar] [CrossRef] [PubMed]
- Salvador, L.A.; Taori, K.; Biggs, J.S.; Jakoncic, J.; Ostrov, D.A.; Paul, V.J.; Luesch, H. Potent Elastase Inhibitors from Cyanobacteria: Structural Basis and Mechanisms Mediating Cytoprotective and Anti-Inflammatory Effects in Bronchial Epithelial Cells. J. Med. Chem. 2013, 56, 1276–1290. [Google Scholar] [CrossRef] [PubMed]
- Schmid, R.; Heuckeroth, S.; Korf, A.; Smirnov, A.; Myers, O.; Dyrlund, T.S.; Bushuiev, R.; Murray, K.J.; Hoffmann, N.; Lu, M.; et al. Integrative analysis of multimodal mass spectrometry data in MZmine 3. Nat. Biotechnol. 2023, 41, 447–449. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Mohimani, H.; Gurevich, A.; Shlemov, A.; Mikheenko, A.; Korobeynikov, A.; Cao, L.; Shcherbin, E.; Nothias, L.-F.; Dorrestein, P.C.; Pevzner, P.A. Dereplication of microbial metabolites through database search of mass spectra. Nat. Commun. 2018, 9, 4035. [Google Scholar] [CrossRef]
- Lee, W.; Rahimi, M.; Lee, Y.; Chiu, A. POKY: A software suite for multidimensional NMR and 3D structure calculation of biomolecules. Bioinformatics 2021, 37, 3041–3042. [Google Scholar] [CrossRef]
- Pluotno, A.; Carmeli, S. Banyasin A and banyasides A and B, three novel modified peptides from a water bloom of the cyanobacterium Nostoc sp. Tetrahedron 2005, 61, 575–583. [Google Scholar] [CrossRef]
- Ocampo Bennet, X. Peptide au Seiner Cyanobakterien Wasserblütte (1998) aus dem Wannsee/Berli: Strukturen and Biologische Wirksamkeit; University Freiburg: Freiburg, Germany, 2007; 28p. [Google Scholar]
- Felczykowska, A.; Pawlik, A.; Mazur-Marzec, H.; Toruńska-Sitarz, A.; Narajczyk, M.; Richert, M.; Węgrzyn, G.; Herman-Antosiewicz, A. Selective inhibition of cancer cells’ proliferation by compounds included in extracts from Baltic Sea cyanobacteria. Toxicon 2015, 108, 1–10. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Structure | TRY | CHY | E | TRB |
|---|---|---|---|---|---|
| [µM] | |||||
| CP 1048 | [Thr + Arg + Ahp + Phe + diMeTyr + Val]Asp + OA | 7.25 | +/− | − | − |
| CP 1034 | [Thr + Arg + Ahp + Phe + MeTyr + Val]Asp + OA | 5.51 | − | − | − |
| CP 1020b | [Thr + Arg + Ahp + Phe + diMeTyr + Val]Asp + HA | 0.39 (0.25 *) | 3.6 (3.1 *) | − | − |
| CP 1018 | [Thr + Arg + Ahp + Phe + MePhe + Val]Asp + OA | 0.28 (0.24 *) | − | − | − |
| CP 992 | [Thr + Arg + Ahp + Phe + diMeTyr + Val]Asp + BA | 0.31 (0.24 *) | 3.32 (3.5 *) | − | − |
| CP 990 | [Thr + Arg + Ahp + Phe + MePhe + Val]Asp + HA | 3.73 | − | − | − |
| CP 978 | [Thr + Arg + Ahp + Phe + MeTyr + Val]Asp + BA | 0.29 (0.26 *) | 4.2 (3.8 *) | − | − |
| CP 962 | [Thr + Arg + Ahp + Phe + MePhe + Val]Asp + BA | 3.18 | − | − | − |
| CP 950 | [Thr + Arg + Ahp + Phe + MeTyr + Val]Asp + Ac | 0.66 | − | − | − |
| CP 934 | [Thr + Arg + Ahp + Phe + MePhe + Val]Asp + Ac | 0.42 | 6.75 | − | − |
| CP 809 | [Thr + Arg + Ahp + Phe + diMeTyr + Val] | − | − | − | − |
| CP 778 | [Thr + Arg + Ahp + Phe + MePhe + Val] | − | − | − | − |
| CP 1055 | [Thr + Tyr + Ahp + Phe + diMeTyr + Val]Asp + OA | − | 3.69 | − | − |
| CP 1027 | [Thr + Tyr + Ahp + Phe + diMeTyr + Val]Asp + HA | − | 0.38 (0.26 *) | − | − |
| CP 1025 | [Thr + Tyr + Ahp + Phe + MePhe + Val]Asp + OA | − | +/− | − | − |
| CP 1013 | [Thr + Tyr + Ahp + Phe + MeTyr + Val]Asp + HA | − | 3.97 | − | − |
| CP 999 | [Thr + Tyr + Ahp + Phe + diMeTyr + Val]Asp + BA | − | + | − | − |
| CP 997b | [Thr + Tyr + Ahp + Phe + MePhe + Val]Asp + HA | − | 7.10 | − | − |
| CP 985 | [Thr + Tyr + Ahp + Phe + MeTyr + Val]Asp + BA | − | 0.49 (0.26 *) | − | − |
| CP 972 | [Thr + Tyr + Ahp + Phe + diMeTyr + Val]Asp + Ac | − | 5.19 | − | − |
| CP 969 | [Thr + Tyr + Ahp + Phe + MePhe + Val]Asp + BA | − | 1.94 | − | − |
| CP 958 | [Thr + Tyr + Ahp + Phe + MeTyr + Val]Asp + Ac | − | 0.38 | − | − |
| CP 941 | [Thr + Tyr + Ahp + Phe + MePhe + Val]Asp + Ac | − | 0.7 | − | − |
| CP 983b | [Thr + Tyr + Ahp + Phe + MePhe + Leu]Asp + BA | − | 2.49 | − | − |
| CP 949 | [Thr + Leu + Ahp + Phe + diMeTyr + Val]Asp + BA | − | 1.59 | 3.32 | − |
| CP 935 | [Thr + Leu + Ahp + Phe + MeTyr + Val]Asp + BA | − | 4.92 | − | − |
| CP 919 | [Thr + Leu + Ahp + Phe + MePhe + Val]Asp + BA | − | 1.45 | 5.71 | − |
| CP 1011 | [Thr + Phe + Ahp + Phe + diMeTyr + Val]Asp + HA | − | − | − | − |
| CP 997 | [Thr + Phe + Ahp + Phe + MeTyr + Val]Asp + HA | − | 4.64 | − | − |
| CP 981 | [Thr + Phe + Ahp + Phe + MePhe + Val]Asp + HA | − | 3.92 | − | − |
| CP 983 | [Thr + Phe + Ahp + Phe + diMeTyr + Val]Asp + BA | − | 0.99 | − | − |
| CP 969b | [Thr + Phe + Ahp + Phe + MeTyr + Val]Asp + BA | − | 4.78 | − | − |
| CP 953 | [Thr + Phe + Ahp + Phe + MePhe + Val]Asp + BA | − | 5.95 | − | − |
| CP 925 | [Thr + Phe + Ahp + Phe + MePhe + Val]Asp + Ac | − | 5.45 | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konkel, R.; Cegłowska, M.; Szubert, K.; Wieczerzak, E.; Iliakopoulou, S.; Kaloudis, T.; Mazur-Marzec, H. Structural Diversity and Biological Activity of Cyanopeptolins Produced by Nostoc edaphicum CCNP1411. Mar. Drugs 2023, 21, 508. https://doi.org/10.3390/md21100508
Konkel R, Cegłowska M, Szubert K, Wieczerzak E, Iliakopoulou S, Kaloudis T, Mazur-Marzec H. Structural Diversity and Biological Activity of Cyanopeptolins Produced by Nostoc edaphicum CCNP1411. Marine Drugs. 2023; 21(10):508. https://doi.org/10.3390/md21100508
Chicago/Turabian StyleKonkel, Robert, Marta Cegłowska, Karolina Szubert, Ewa Wieczerzak, Sofia Iliakopoulou, Triantafyllos Kaloudis, and Hanna Mazur-Marzec. 2023. "Structural Diversity and Biological Activity of Cyanopeptolins Produced by Nostoc edaphicum CCNP1411" Marine Drugs 21, no. 10: 508. https://doi.org/10.3390/md21100508
APA StyleKonkel, R., Cegłowska, M., Szubert, K., Wieczerzak, E., Iliakopoulou, S., Kaloudis, T., & Mazur-Marzec, H. (2023). Structural Diversity and Biological Activity of Cyanopeptolins Produced by Nostoc edaphicum CCNP1411. Marine Drugs, 21(10), 508. https://doi.org/10.3390/md21100508