

Physiological Effects of Oxidative Stress Caused by Saxitoxin in the Nematode Caenorhabditis elegans


Abstract
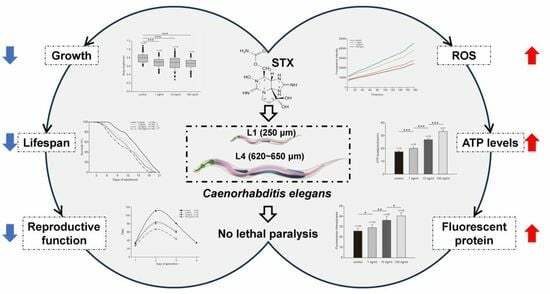
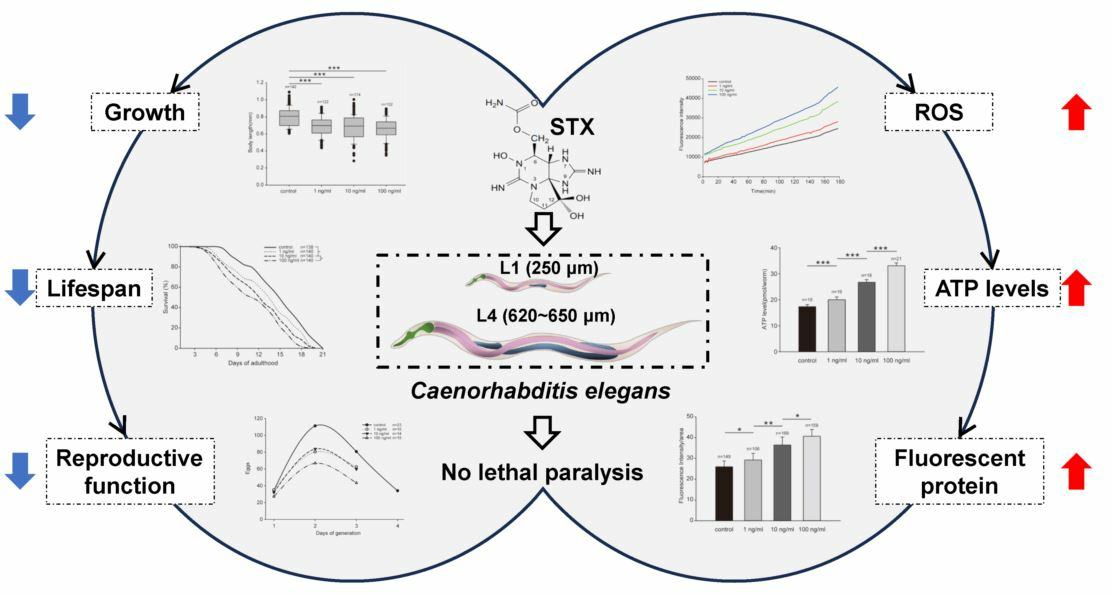
:
1. Introduction
2. Results
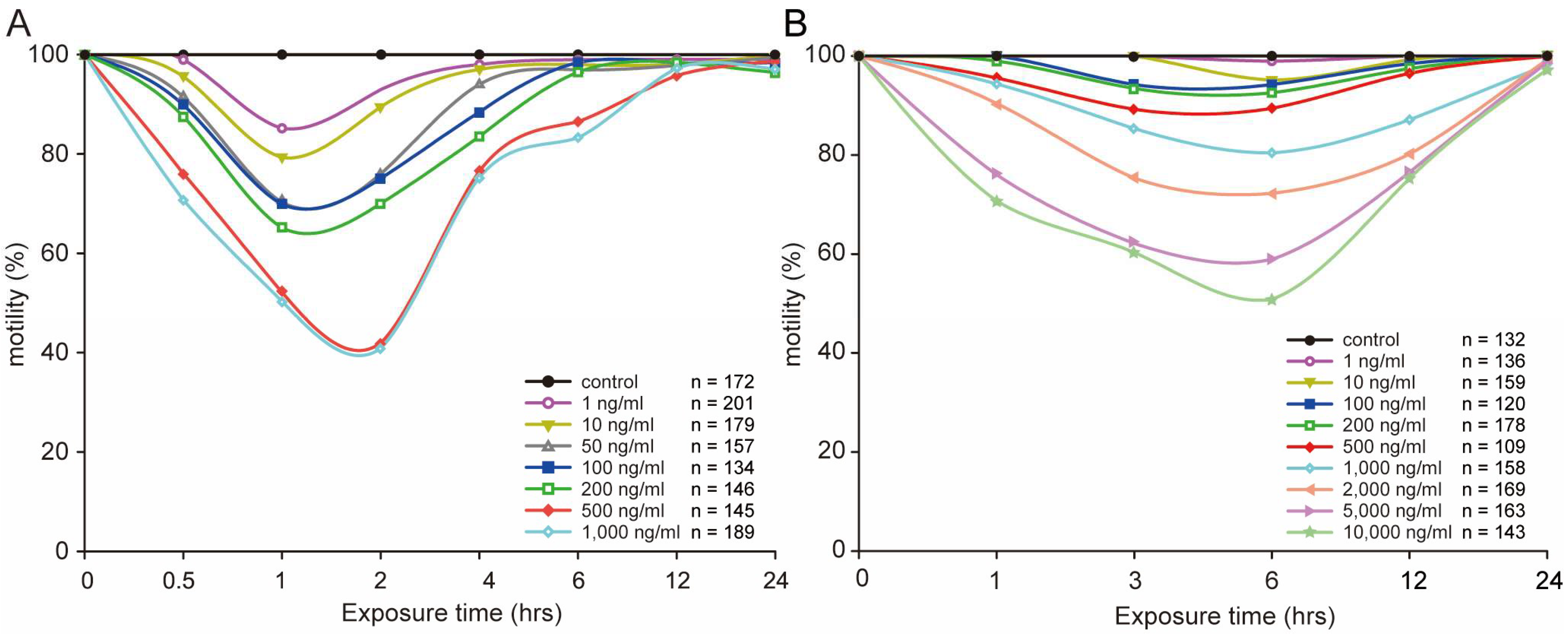
2.1. STX Caused no Lethal Paralytic Reaction in C. elegans
2.2. STX Induces the Production of ROS in Worms
2.3. STX Induced Body Size Decrease in C. elegans
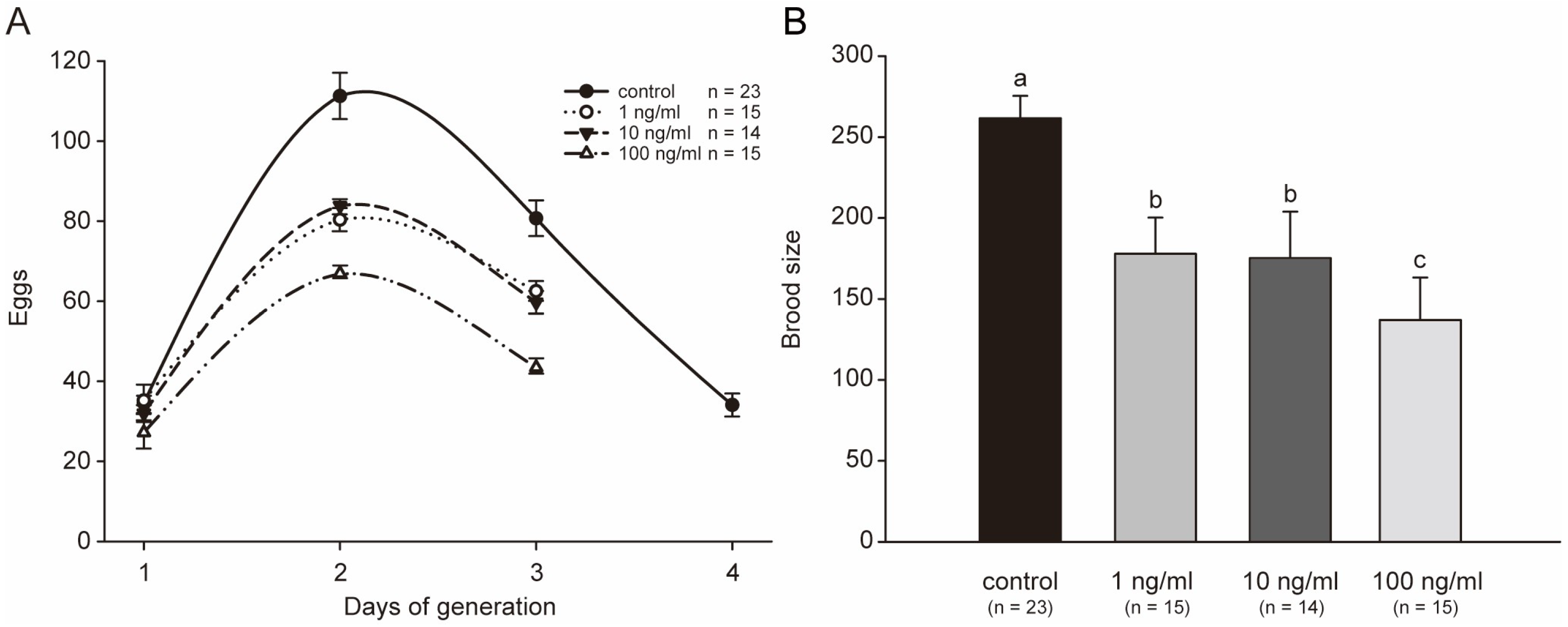
2.4. STX Decreases the Reproductive Function of C. elegans
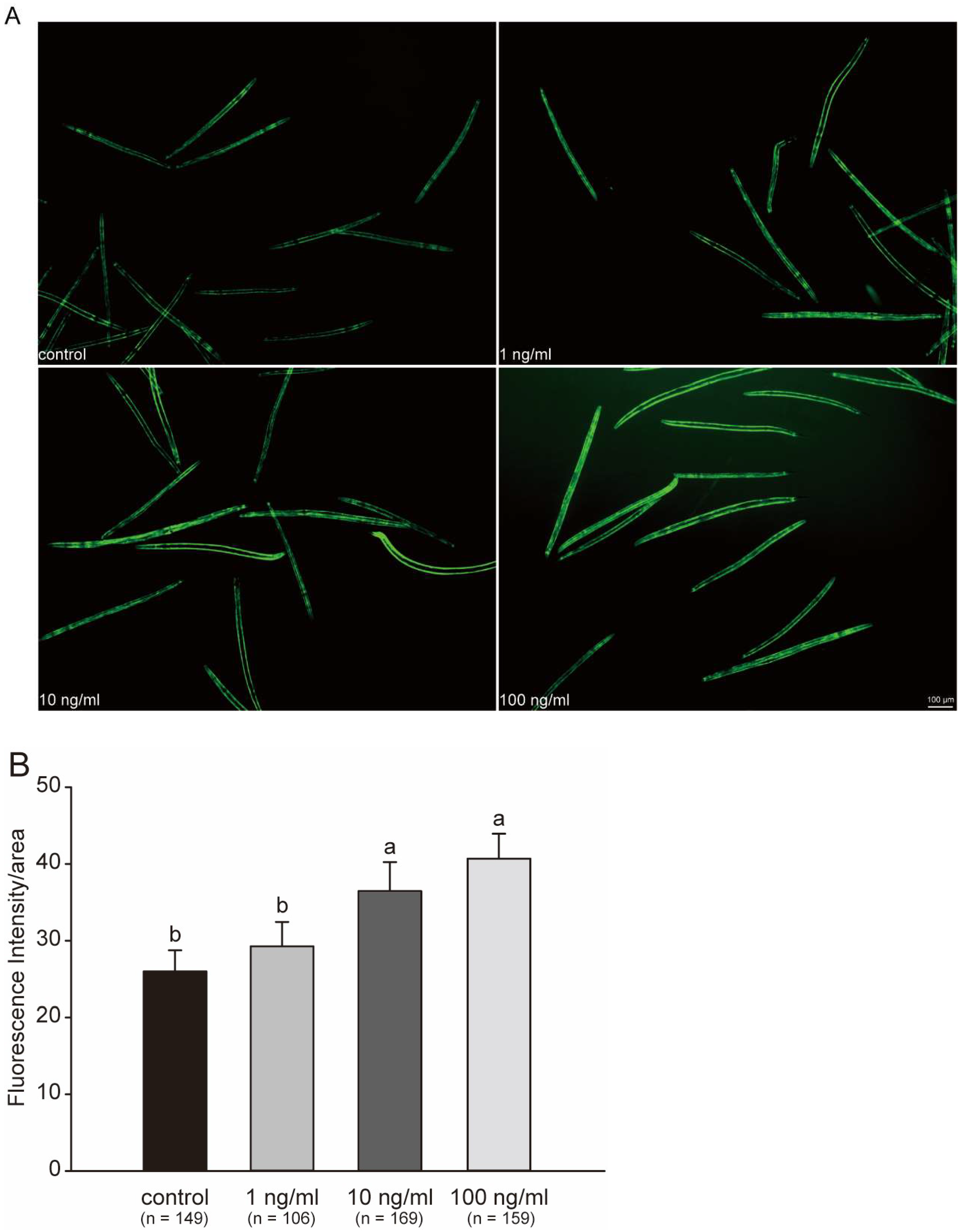
2.5. Fluorescent Protein Expression in GFP Worms
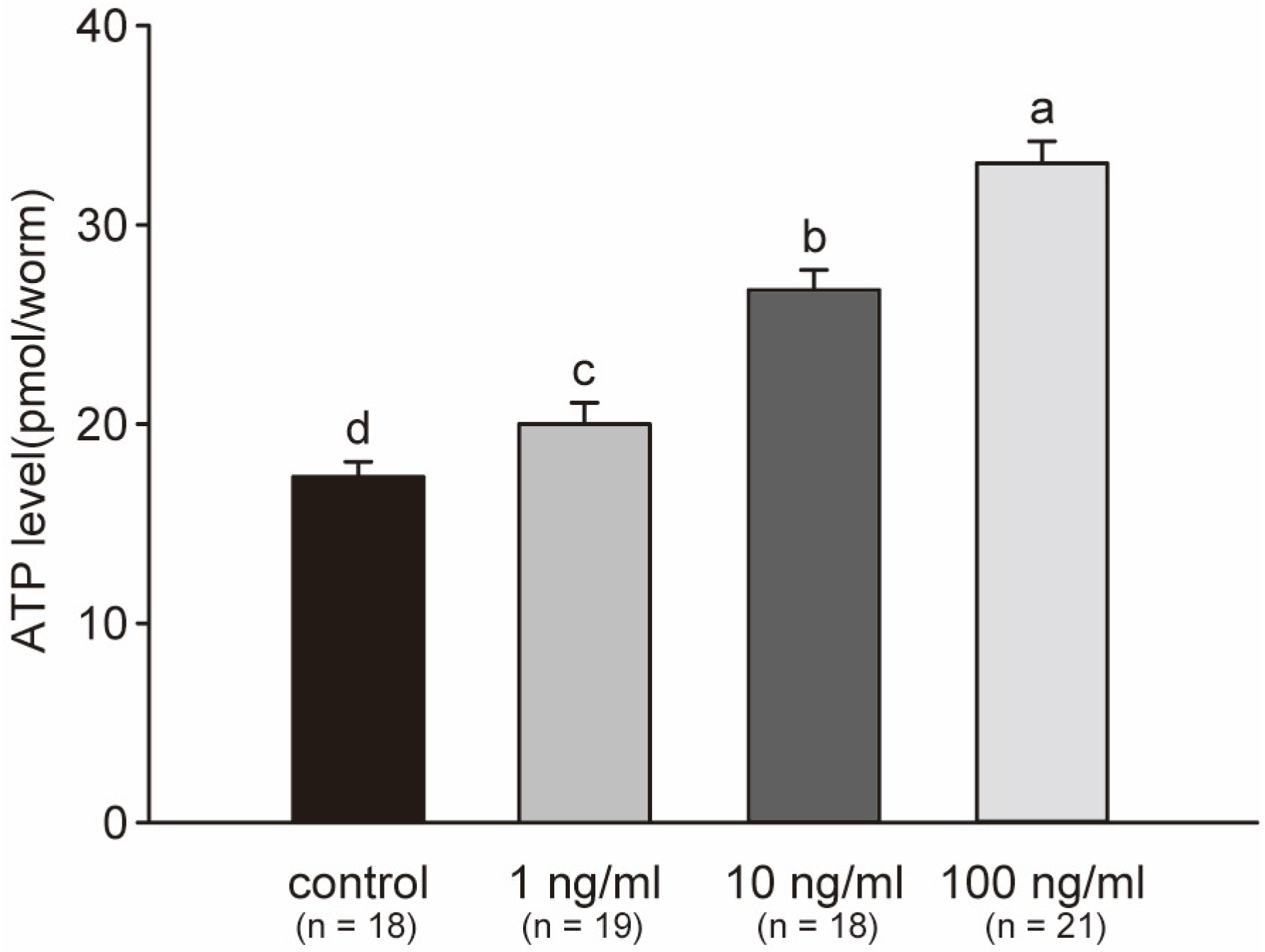
2.6. ATP Levels Significantly Increase after Exposure to STX
3. Discussion
4. Materials and Methods
4.1. Chemical and Toxin Preparation
4.2. Strains
4.3. Toxicity Assessment
4.4. Measurement of ROS Production
4.5. Growth Assays, Lifespan Assays and Brood Size
4.6. Quantification of Fluorescent Protein Intensity in GFP Worms
4.7. Adenosine Triphosphate (ATP) Assays
4.8. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- European Food Safety Authority. Marine biotoxins in shellfish—Saxitoxin group. EFSA J. 2009, 7, 1019. [Google Scholar] [CrossRef]
- Landsberg, J.H. The effects of harmful algal blooms on aquatic organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar] [CrossRef]
- Burson, A.; Matthijs, H.; Bruijne, W.D.; Talens, R.; Huisman, J. Termination of a toxic Alexandrium bloom with hydrogen peroxide. Harmful Algae 2014, 31, 125–135. [Google Scholar] [CrossRef]
- Cusick, K.D.; Minkin, S.C.; Dodani, S.C.; Chang, C.J.; Wilhelm, S.W.; Sayler, G.S. Inhibition of copper uptake in yeast reveals the copper transporter Ctr1p as a potential molecular target of saxitoxin. Environ. Sci. Technol. 2012, 46, 2959–2966. [Google Scholar] [CrossRef] [PubMed]
- Kozlowsky-Suzuki, B.; Koski, M.; Hallberg, E.; Wallen, R.; Carlsson, P. Glutathione transferase activity and oocyte development in copepods exposed to toxic phytoplankton. Harmful Algae 2009, 8, 395–406. [Google Scholar] [CrossRef]
- Tan, Z.J.; Yan, T.; Yu, R.C.; Zhou, M.J. Transfer of paralytic shellfish toxins via marine food chains: A simulated experiment. Biomed. Environ. Sci. 2007, 20, 235. [Google Scholar] [CrossRef]
- Bricelj, V.M.; Ford, S.E.; Lambert, C.; Barbou, A.; Paillard, C. Effects of toxic Alexandrium tamarense on behavior, hemocyte responses and development of brown ring disease in Manila clams. Mar. Ecol. Prog. Ser. 2011, 430, 35–48. [Google Scholar] [CrossRef]
- Navarro, J.M.; Aguila, B.L.; Machmar, F.; Chaparro, O.R.; Contreras, A.M. Dynamic of intoxication and detoxification in juveniles of Mytilus chilensis (Bivalvia: Mytilidae) exposed to paralytic shellfish toxins. Aquat. Living Resour. 2011, 24, 93–98. [Google Scholar] [CrossRef]
- Goc, N.L.; Hégaret, H.; Fabioux, C.; Miner, P.; Soudant, P. Impact of the toxic dinoflagellate Alexandrium catenella on Pacific oyster reproductive output: Application of flow cytometry assays on spermatozoa. Aquat. Living Resour. 2013, 26, 221–228. [Google Scholar] [CrossRef]
- Nielsen, M.V.; Strmgren, T. Shell growth response of mussels (Mytilus edulis) exposed to toxic microalgae. Mar. Biol. 1991, 108, 263–267. [Google Scholar] [CrossRef]
- Tran, D.; Haberkorn, H.; Soudant, P.; Ciret, P.; Massabuau, J.C. Behavioral responses of Crassostrea gigas exposed to the harmful algae Alexandrium minutum. Aquaculture 2010, 298, 338–345. [Google Scholar] [CrossRef]
- Yan, T.; Zhou, M.J.; Fu, M.; Wang, L. Study on impact of dinoflagellate Alexandrium tamarense on life activities of marine bivalves. Acta Oceanol. Sin. 2003, 22, 103–109. [Google Scholar]
- Bif, M.B.; Yunes, J.S.; Resgalla, C., Jr. Evaluation of mysids and sea urchins exposed to saxitoxins. Environ. Toxicol. Pharmacol. 2013, 36, 819–825. [Google Scholar] [CrossRef]
- Cembella, A.D.; Quilliam, M.A.; Lewis, N.I.; Bauder, A.G.; Dell’Aversano, C.; Thomas, K.; Jellett, J.; Cusack, R.R. The toxigenic marine dinoflagellate Alexandrium tamarense as the probable cause of mortality of caged salmon in Nova Scotia. Harmful Algae 2002, 1, 313–325. [Google Scholar] [CrossRef]
- Fire, S.E.; Pruden, J.; Couture, D.; Wang, Z.; Bottein, M.Y.D.; Haynes, B.L.; Knott, T.; Bouchard, D.; Lichtenwalner, A.; Wippelhauser, G. Saxitoxin exposure in an endangered fish: Association of a shortnose sturgeon mortality event with a harmful algal bloom. Mar. Ecol. Prog. Ser. 2012, 460, 145–153. [Google Scholar] [CrossRef]
- Zhang, D.; Hu, C.; Wang, G.; Li, D.; Liu, Y. Zebrafish neurotoxicity from aphantoxins-cyanobacterial paralytic shellfish poisons (PSPs) from aphanizomenon flos-aquae DC-1. Environ. Toxicol. 2013, 28, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Ferrão-Filho, A.D.S.; Kozlowsky-Suzuki, B. Cyanotoxins: Bioaccumulation and effects on aquatic animals. Mar. Drugs 2011, 9, 2729–2772. [Google Scholar] [CrossRef]
- Niedzwiadek, B.; Scott, P.M.; Lau, P.Y. Monitoring of shrimp and farmed fish sold in Canada for cyanobacterial toxins. J. Food Prot. 2012, 75, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.K.; Lacaze, J.P.; Hermann, G.; Kershaw, J.; Brownlow, A.; Turner, A.; Hall, A. Detection and effects of harmful algal toxins in Scottish harbour seals and potential links to population decline. Toxicon 2015, 97, 1–14. [Google Scholar] [CrossRef]
- Pearson, L.; Mihali, T.; Moffitt, M.; Kellmann, R.; Neilan, B. On the chemistry, toxicology and genetics of the cyanobacterial toxins, microcystin, nodularin, saxitoxin and cylindrospermopsin. Mar. Drugs 2010, 8, 1650–1680. [Google Scholar] [CrossRef]
- Li, S.C.; Wang, W.X.; Hsieh, D. Effects of toxic dinoflagellate Alexandrium tamarense on the energy budgets and growth of two marine bivalves. Mar. Environ. Res. 2002, 53, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Cheng, J.; Chen, X.; Cheng, S.H.; Mak, Y.L.; Lam, P.; Chan, L.L.; Wang, M. Early developmental toxicity of saxitoxin on medaka (Oryzias melastigma) embryos. Toxicon 2014, 77, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Al-Yamani, F.Y.; Polikarpov, I.; Saburova, M. Marine life mortalities and harmful algal blooms in the Northern Arabian Gulf. Aquat. Ecosyst. Health Manag. 2020, 23, 196–209. [Google Scholar] [CrossRef]
- Burridge, L.E.; Martin, J.L.; Lyons, M.C.; Legresley, M.M. Lethality of microalgae to farmed Atlantic salmon (Salmo salar). Aquaculture 2010, 308, 101–105. [Google Scholar] [CrossRef]
- Etheridge, S.M. Paralytic shellfish poisoning: Seafood safety and human health perspectives. Toxicon 2010, 56, 108–122. [Google Scholar] [CrossRef]
- Hille, B. The receptor for tetrodotoxin and saxitoxin. A structural hypothesis. Biophys. J. 1975, 15, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Li, Z.; Jiang, Y.; Pan, X.; Wu, J.; Ben, C.A.; Smith, J.J.; Chin, Y.; Lei, J.; Zhou, Q. Structural basis for the modulation of voltage-gated sodium channels by animal toxins. Science 2018, 362, eaau2596. [Google Scholar] [CrossRef]
- César, M.; Christian, L. The voltage-gated sodium channel: A major target of marine neurotoxins. Toxicon 2014, 91, 84–85. [Google Scholar] [CrossRef]
- Jiang, T.J.; Niu, T.; Xu, Y.X. Transfer and metabolism of paralytic shellfish poisoning from scallop (Chlamys nobilis) to spiny lobster (Panulirus stimpsoni). Toxicon 2006, 48, 988–994. [Google Scholar] [CrossRef]
- Kwong, R.; Wang, W.X.; Lam, P.; Yu, P. The uptake, distribution and elimination of paralytic shellfish toxins in mussels and fish exposed to toxic dinoflagellates. Aquat. Toxicol. 2006, 80, 82–91. [Google Scholar] [CrossRef]
- Turner, A.D.; Lewis, A.M.; Hatfield, R.G.; Galloway, A.W.; Higman, W.A. Transformation of paralytic shellfish poisoning toxins in Crassostrea gigas and Pecten maximus reference materials. Toxicon 2012, 60, 1117–1134. [Google Scholar] [CrossRef] [PubMed]
- Cusick, K.D.; Wetzel, R.K.; Minkin, S.C.; Dodani, S.C.; Sayler, G.S. Paralytic shellfish toxins inhibit copper uptake in Chlamydomonas reinhardtii. Environ. Toxicol. Chem. 2013, 32, 1388–1395. [Google Scholar] [CrossRef] [PubMed]
- Choi, N.; Yeung, L.; Siu, W.; So, I.; Jack, R.W.; Hsieh, D.; Wu, R.; Lam, P. Relationships between tissue concentrations of paralytic shellfish toxins and antioxidative responses of clams, Ruditapes philippinarum. Mar. Pollut. Bull. 2006, 52, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Ma, F.; Hua, F.; Li, A. Effects of feeding Alexandrium tamarense, a paralytic shellfish toxin producer, on antioxidant enzymes in scallops (Patinopecten yessoensis) and mussels (Mytilus galloprovincialis). Aquaculture 2013, 396, 76–81. [Google Scholar] [CrossRef]
- Fabioux, C.; Sulistiyani, Y.; Haberkorn, H.; Hegaret, H.; Amzil, Z.; Soudant, P. Exposure to toxic Alexandrium minutum activates the detoxifying and antioxidant systems in gills of the oyster Crassostrea gigas. Harmful Algae 2015, 48, 55–62. [Google Scholar] [CrossRef]
- Gubbins, M.J.; Eddy, F.B.; Gallacher, S.; Stagg, R.M. Paralytic shellfish poisoning toxins induce xenobiotic metabolising enzymes in Atlantic salmon (Salmo salar). Mar. Environ. Res. 2000, 50, 479–483. [Google Scholar] [CrossRef]
- Costa, P.R.; Pereira, P.; Guilherme, S.; Barata, M.; Santos, M.A.; Pacheco, M.; Pousao-Ferreira, P. Hydroxybenzoate paralytic shellfish toxins induce transient GST activity depletion and chromosomal damage in white seabream (Diplodus sargus). Mar. Environ. Res. 2012, 79, 63–69. [Google Scholar] [CrossRef]
- Painefilú, J.C.; Bianchi, V.A.; Krock, B.; De Anna, J.S.; Kristoff, G.; Luquet, C.M. Effects of paralytic shellfish toxins on the middle intestine of Oncorhynchus mykiss: Glutathione metabolism, oxidative status, lysosomal function and ATP-binding cassette class C (ABCC) proteins activity. Ecotoxicol. Environ. Saf. 2020, 204, 111069. [Google Scholar] [CrossRef]
- Durbin, E.; Teegarden, G.; Campbell, R.; Cembella, A.; Baumgartner, M.F.; Mate, B.R. North Atlantic right whales, Eubalaena glacialis, exposed to paralytic shellfish poisoning (PSP) toxins via a zooplankton vector, Calanus finmarchicus. Harmful Algae 2002, 1, 243–251. [Google Scholar] [CrossRef]
- Doucette, G.J.; Cembella, A.D.; Martin, J.L.; Michaud, J.; Cole, T.; Rolland, R.M. Paralytic shellfish poisoning (PSP) toxins in North Atlantic right whales Eubalaena glacialis and their zooplankton prey in the Bay of Fundy, Canada. Mar. Ecol. Prog. 2006, 306, 303–313. [Google Scholar] [CrossRef]
- Cusick, K.; Gary, S. An overview on the marine neurotoxin, saxitoxin: Genetics, molecular targets, methods of detection and ecological functions. Mar. Drugs 2013, 11, 911–1018. [Google Scholar] [CrossRef] [PubMed]
- Thottumkara, A.P.; Parsons, W.H.; Du Bois, J. Saxitoxin. Angew. Chem. Int. Ed. 2014, 53, 5760–5784. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Zhen, M. Action potentials drive body wall muscle contractions in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2011, 108, 2557–2562. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Li, K.; Lu, W.; Li, S.; Chen, X.; Liu, X.; Yuan, J.; Ding, Q.; Lan, F.; Cai, S. Identification of small-molecule ion channel modulators in C. elegans channelopathy models. Nat. Commun. 2018, 9, 3912–3941. [Google Scholar] [CrossRef]
- Wang, J.; Salata, J.J.; Bennett, P.B. Saxitoxin is a gating modifier of HERG K+ channels. J. Gen. Physiol. 2003, 121, 583–598. [Google Scholar] [CrossRef]
- Wang, D.Y.; Wang, Y. Phenotypic and behavioral defects caused by barium exposure in nematode Caenorhabditis elegans. Arch. Environ. Contam. Toxicol. 2008, 54, 447–453. [Google Scholar] [CrossRef]
- Arantes, O.N.; Apfeld, J.; Dillin, A.; Kenyon, C. Regulation of life-span by germ-line stem cells in Caenorhabditis elegans. Science 2002, 295, 502–505. [Google Scholar] [CrossRef]
- Wu, Y.C.; Wang, X.; Ding, X. Methods for studying programmed cell death in C. elegans. Methods Cell Biol. 2012, 107, 295–320. [Google Scholar] [CrossRef]
- Chen, X.; Zhong, Z.; Xu, Z.; Chen, L.; Wang, Y. 2′,7′-Dichlorodihydrofluorescein as a fluorescent probe for reactive oxygen species measurement: Forty years of application and controversy. Free Radic. Res. Commun. 2010, 44, 587–604. [Google Scholar] [CrossRef]
- Meng, Q.X.; Wang, W.Y.; Lu, Q.M.; Jin, Y.; Wei, J.F.; Zhu, S.W.; Xiong, Y.L. A novel short neurotoxin, cobrotoxin c, from monocellate cobra (Naja kaouthia) venom: Isolation and purification, primary and secondary structure determination, and tertiary structure modeling. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2002, 132, 113–121. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, C.; Zhang, H.; Liu, R.; Yin, L. Integrating transcriptomics and behavior tests reveals how the C. elegans responds to copper induced aging. Ecotoxicol. Environ. Saf. 2021, 222, 112494. [Google Scholar] [CrossRef] [PubMed]
- Ros-Santaella, J.L. Impact of oxidative stress on male reproduction in domestic and wild animals. Antioxidants 2021, 10, 1154. [Google Scholar] [CrossRef]
- Tian, Y.; Zhou, M.; Meng, F.; Yu, R.; Wang, Y.; Li, J. Effects of the dinoflagellate Alexandrium tamarense on early development of the scallop Argopecten irradians concentricus. Aquaculture 2003, 217, 167–178. [Google Scholar] [CrossRef]
- Yan, T.; Zhou, M.; Fu, M.; Wang, Y.; Yu, R.; Li, J. Inhibition of egg hatching success and larvae survival of the scallop, Chlamys farreri, associated with exposure to cells and cell fragments of the dinoflagellate Alexandrium tamarense. Toxicon 2001, 39, 1239–1244. [Google Scholar] [CrossRef]
- Yurchenko, O.V.; Radashevsky, V.I.; Hsieh, H.L.; Reunov, A.A. Ultrastructural comparison of the spermatozoa of the Pacific oyster Crassostrea gigas inhabiting polluted and relatively clean areas in Taiwan. Aquat. Ecol. 2009, 43, 513–519. [Google Scholar] [CrossRef]
- Haberkorn, H.; Lambert, C.; Goiec, N.L.; Moal, J.; Suquet, M.; Gueguen, M.; Sunila, I.; Soudant, P. Effects of Alexandrium minutum exposure on nutrition-related processes and reproductive output in oysters Crassostrea gigas. Harmful Algae 2010, 9, 427–439. [Google Scholar] [CrossRef]
- Zhijun, T.; Tian, Y.; Mingjiang, Z.; Jun, L.; Rencheng, Y.; Yunfeng, W. The effects of Alexandrium tamarense on survival, growth and reproduction of Neomysis awatschensis. Acta Ecol. Sin. 2002, 22, 1635–1639. [Google Scholar] [CrossRef]
- Piazza, V.; Ullmann, C.V.; Aberhan, M. Temperature-related body size change of marine benthic macroinvertebrates across the Early Toarcian Anoxic Event. Sci. Rep. 2020, 10, 4675. [Google Scholar] [CrossRef]
- Turner, R.E.; Plunket, J.S. Estuarine oiling increases a long-term decline in mussel growth. Environ. Pollut. 2021, 284, 117506. [Google Scholar] [CrossRef]
- Grande, F.R.D.; Granado, P.; Costa, T.M. Size-at-age or structure shift: Which hypothesis explains smaller body size of the fiddler crab Leptuca uruguayensis in northern populations? Estuar. Coast. Shelf Sci. 2021, 254, 107358. [Google Scholar] [CrossRef]
- Genner, M.J.; Sims, D.W.; Southward, A.J.; Budd, G.C.; Masterson, P.; Mchugh, M.; Rendle, P.; Southall, E.J.; Wearmouth, V.J.; Hawkins, S.J. Body size-dependent responses of a marine fish assemblage to climate change and fishing over a century-long scale. Glob. Chang. Biol. 2010, 16, 517–527. [Google Scholar] [CrossRef]
- Luckenbach, M.W.; Sellner, K.G.; Shumway, S.E.; Greene, K. Effects of two bloom-forming dinoflagellates, Prorocentrum mariae-lebouriae and Gyrodinium unctenumon on the growth and survival of the eastern oyster, Crassostrea virginica (Gmelin, 1791). J. Shellfish Res. 1993, 12, 411–415. [Google Scholar]
- Turnbull, A.; Malhi, N.; Seger, A.; Jolley, J.; Fitzgibbon, Q. Accumulation of paralytic shellfish toxins by Southern Rock Lobster Jasus edwardsii causes minimal impact on lobster health. Aquat. Toxicol. 2020, 230, 105704. [Google Scholar] [CrossRef]
- Duan, J.; Liu, H.; Zhu, J.; Lu, L.; Chang, L. A dynamic energy budget model for abalone, Haliotis discus hannai Ino. Ecol. Model. 2021, 451, 109569. [Google Scholar] [CrossRef]
- Pousse, E.; Flye-Sainte-Marie, J.; Alunno-Bruscia, M.; Hegaret, H.; Rannou, E.; Pecquerie, L.; Marques, G.M.; Thomas, Y.; Castrec, J.; Fabioux, C. Modelling paralytic shellfish toxins (PST) accumulation in Crassostrea gigas by using Dynamic Energy Budgets (DEB). J. Sea Res. 2019, 143, 152–164. [Google Scholar] [CrossRef]
- Bruynsteen, L.; Janssens, G.P.J.; Harris, P.A.; Duchateau, L.; Valle, E.; Odetti, P.; Vandevelde, K.; Buyse, J.; Hesta, M. Changes in oxidative stress in response to different levels of energy restriction in obese ponies. Br. J. Nutr. 2014, 112, 1402–1411. [Google Scholar] [CrossRef]
- Rolfe, D.F.; Brown, G.C. Cellular energy utilization and molecular origin of standard metabolic rate in mammals. Physiol. Rev. 1997, 77, 731–758. [Google Scholar] [CrossRef] [PubMed]
- Stenesen, D.; Suh, J.M.; Seo, J.; Yu, K.; Lee, K.S.; Kim, J.S.; Min, K.J.; Graff, J.M. Adenosine nucleotide biosynthesis and AMPK regulate adult life span and mediate the longevity benefit of caloric restriction in flies. Cell Metab. 2013, 17, 101–112. [Google Scholar] [CrossRef]
- Ranjan, M.; Gruber, J.; Ng, L.F.; Halliwell, B. Repression of the mitochondrial peroxiredoxin antioxidant system does not shorten life span but causes reduced fitness in Caenorhabditis elegans. Free Radic. Biol. Med. 2013, 63, 381–389. [Google Scholar] [CrossRef]
- Wu, S.B.; Wei, Y.H. AMPK-mediated increase of glycolysis as an adaptive response to oxidative stress in human cells: Implication of the cell survival in mitochondrial diseases. Biochim. Biophys. Acta-Mol. Basis Dis. 2012, 1822, 233–247. [Google Scholar] [CrossRef]
- Yanase, S.; Yasuda, K.; Ishii, N. Adaptive responses to oxidative damage in three mutants of Caenorhabditis elegans (age-1, mev-1 and daf-16) that affect life span. Mech. Ageing Dev. 2002, 123, 1579–1587. [Google Scholar] [CrossRef] [PubMed]
- Acba, B.; Vp, B.; Rm, B.; Am, B.; Sg, B.; Prca, C.; Mp, B. DNA damage and oxidative stress responses of mussels Mytilus galloprovincialis to paralytic shellfish toxins under warming and acidification conditions—Elucidation on the organ-specificity. Aquat. Toxicol. 2020, 228, 105619. [Google Scholar] [CrossRef]
- Almansa-Ordonez, A.; Bellido, R.; Vassena, R.; Barragan, M.; Zambelli, F. Oxidative stress in reproduction: A mitochondrial perspective. Biology 2020, 9, 269. [Google Scholar] [CrossRef]
- Napolitano, G.; Fasciolo, G.; Venditti, P. Mitochondrial management of reactive oxygen species. Antioxidants 2021, 10, 1824. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, Y.; Zhang, Z.; Li, Z.; Cai, P. Enhancement in the ATP level and antioxidant capacity of Caenorhabditis elegans under continuous exposure to extremely low-frequency electromagnetic field for multiple generations. Int. J. Radiat. Biol. 2020, 96, 1633–1640. [Google Scholar] [CrossRef]
- Kanaan, G.N.; Harper, M.E. Cellular redox dysfunction in the development of cardiovascular diseases. Biochim. Biophys Acta Gen. Subj. 2017, 1861, 2822–2829. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, R.; Li, H.; Xiang, Y.; Xiao, L.; Hu, M.; Ma, F.; Ma, C.W.; Huang, Z. Antioxidant and neuroprotective effects of Dictyophora indusiata polysaccharide in Caenorhabditis elegans. J. Ethnopharmacol. 2016, 192, 413–422. [Google Scholar] [CrossRef]
- Wen, H.; Shi, W.; Qin, J. Multiparameter evaluation of the longevity in C. elegans under stress using an integrated microfluidic device. Biomed. Microdevices 2012, 14, 721–728. [Google Scholar] [CrossRef]
- C. elegans Sequencing Consortium. Genome Sequence of the Nematode C. elegans: A Platform for Investigating Biology. Science 1998, 282, 2012–2018. [Google Scholar] [CrossRef]
- Gruber, J.; Ng, L.F.; Poovathingal, S.K.; Halliwell, B. Deceptively simple but simply deceptive—Caenorhabditis elegans lifespan studies: Considerations for aging and antioxidant effects. FEBS Lett. 2009, 583, 3377–3387. [Google Scholar] [CrossRef]
- Schulz, T.J.; Zarse, K.; Voigt, A.; Urban, N.; Birringer, M.; Ristow, M. Glucose restriction extends Caenorhabditis elegans life span by inducing mitochondrial respiration and increasing oxidative stress. Cell Metab. 2007, 6, 280–293. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Number of C. elegans | Survival Time (Days) | |
|---|---|---|---|
| Total | Censored | ||
| Control | 138 | 0 | 13.88 ± 0.34 a |
| 1 ng/mL | 140 | 0 | 12.20 ± 0.37 b |
| 10 ng/mL | 140 | 0 | 11.33 ± 0.36 c |
| 100 ng/mL | 140 | 0 | 10.38 ± 0.36 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Prithiviraj, B.; Tan, Z. Physiological Effects of Oxidative Stress Caused by Saxitoxin in the Nematode Caenorhabditis elegans. Mar. Drugs 2023, 21, 544. https://doi.org/10.3390/md21100544
Wu H, Prithiviraj B, Tan Z. Physiological Effects of Oxidative Stress Caused by Saxitoxin in the Nematode Caenorhabditis elegans. Marine Drugs. 2023; 21(10):544. https://doi.org/10.3390/md21100544
Chicago/Turabian StyleWu, Haiyan, Balakrishnan Prithiviraj, and Zhijun Tan. 2023. "Physiological Effects of Oxidative Stress Caused by Saxitoxin in the Nematode Caenorhabditis elegans" Marine Drugs 21, no. 10: 544. https://doi.org/10.3390/md21100544
APA StyleWu, H., Prithiviraj, B., & Tan, Z. (2023). Physiological Effects of Oxidative Stress Caused by Saxitoxin in the Nematode Caenorhabditis elegans. Marine Drugs, 21(10), 544. https://doi.org/10.3390/md21100544