

Discovery and Anti-Inflammatory Activity of a Cyanobacterial Fatty Acid Targeting the Keap1/Nrf2 Pathway
 , , and
, , and
Abstract
:
1. Introduction
2. Results
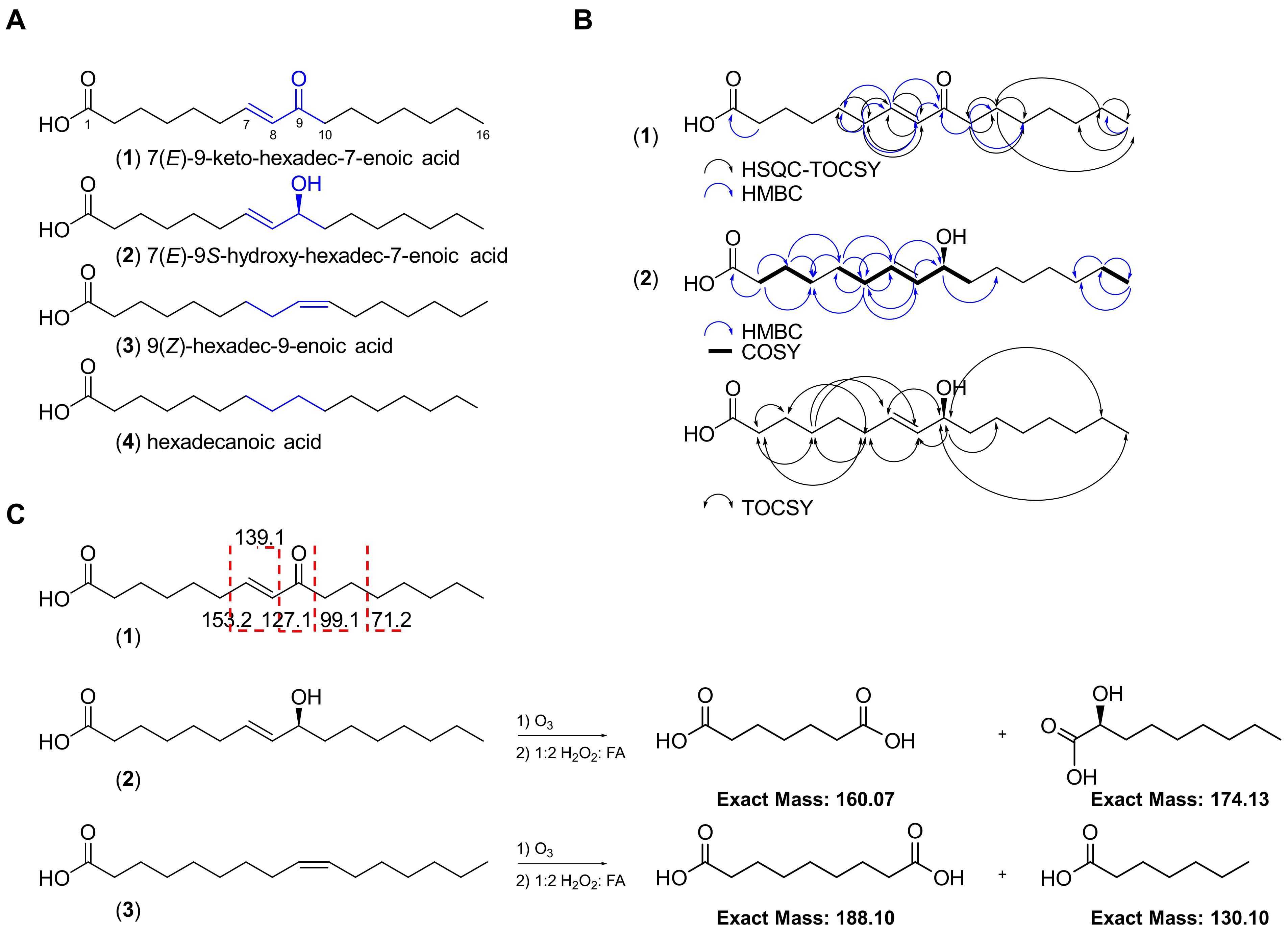
2.1. Isolation and Structure Elucidation
2.2. Biological Activity
2.2.1. ARE-Luciferase Reporter Assay
2.2.2. NO Production
2.2.3. Nqo1 and iNos Transcript Levels
2.2.4. RNA Sequencing
3. Discussion
4. Materials and Methods
4.1. Biological Material
4.2. General Experimental Procedures
4.3. Extraction and Isolation
Isolation of Compound 1 from Other Cyanobacteria Samples
4.4. Ozonolysis and Oxidative Workup
4.5. Cell Culture
4.6. ARE-Luciferase Reporter Assay
4.7. NO Assay
4.8. Cell Viability Assay
4.9. RNA Extraction and RT-qPCR Analysis of Nqo1 and iNos Transcript Levels in RAW264.7 Cells
4.10. Illumina Sequencing Library Construction
4.11. Illumina NovaSeq6000 Sequencing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic Inflammation in the Etiology of Disease across the Life Span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Lugrin, J.; Rosenblatt-Velin, N.; Parapanov, R.; Liaudet, L. The Role of Oxidative Stress during Inflammatory Processes. Biol. Chem. 2014, 395, 203–230. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting Molecular Cross-Talk between Nrf2 and NF-κB Response Pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [PubMed]
- Magesh, S.; Chen, Y.; Hu, L. Small Molecule Modulators of Keap1-Nrf2-ARE Pathway as Potential Preventive and Therapeutic Agents. Med. Res. Rev. 2012, 23, 687–726. [Google Scholar] [CrossRef]
- Egbujor, M.C.; Buttari, B.; Profumo, E.; Telkoparan-Akillilar, P.; Saso, L. An Overview of NRF2-Activating Compounds Bearing α,β-Unsaturated Moiety and Their Antioxidant Effects. Int. J. Mol. Sci. 2022, 23, 8466. [Google Scholar] [CrossRef]
- Liang, S.T.; Chen, C.; Chen, R.X.; Li, R.; Chen, W.L.; Jiang, G.H.; Du, L.L. Michael Acceptor Molecules in Natural Products and Their Mechanism of Action. Front. Pharmacol. 2022, 13, 1033003. [Google Scholar] [CrossRef]
- Hur, W.; Sun, Z.; Jiang, T.; Mason, D.E.; Peters, E.C.; Zhang, D.D.; Luesch, H.; Schultz, P.G.; Gray, N.S. A Small-Molecule Inducer of the Antioxidant Response Element. Chem. Biol. 2010, 17, 537–547. [Google Scholar] [CrossRef]
- Wang, R.; Mason, D.E.; Choe, K.P.; Lewin, A.S.; Peters, E.C.; Luesch, H. In Vitro and in Vivo Characterization of a Tunable Dual-Reactivity Probe of the Nrf2-ARE Pathway. ACS Chem. Biol. 2013, 8, 1764–1774. [Google Scholar] [CrossRef] [PubMed]
- Kanda, H.; Yamawaki, K. Bardoxolone Methyl: Drug Development for Diabetic Kidney Disease. Clin. Exp. Nephrol. 2020, 24, 857–864. [Google Scholar] [CrossRef]
- Boo, Y.C. Natural Nrf2 Modulators for Skin Protection. Antioxidants 2020, 9, 812. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, M.S.; Ratnayake, R.; Pope, J.L.; Chen, Q.Y.; Zhu, F.; Chen, S.; Carney, T.J.; Gharaibeh, R.Z.; Jobin, C.; Paul, V.J.; et al. Seaweed Natural Products Modify the Host Inflammatory Response via Nrf2 Signaling and Alter Colon Microbiota Composition and Gene Expression. Free Radic. Biol. Med. 2020, 146, 306–323. [Google Scholar] [CrossRef]
- Wang, R.; Paul, V.J.; Luesch, H. Seaweed Extracts and Unsaturated Fatty Acid Constituents from the Green Alga Ulva lactuca as Activators of the Cytoprotective Nrf2-ARE Pathway. Free Radic. Biol. Med. 2013, 57, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.; Singh, R.P.; Kumar, A.; Singh, P.K.; Gupta, R.K. Therapeutic Potential of Cyanobacteria as a Producer of Novel Bioactive Compounds. In Cyanobacterial Biotechnology in the 21st Century; Springer Nature: Singapore, 2023; pp. 237–252. [Google Scholar]
- Leão, P.N.; Martins, T.P.; Abt, K.; Reis, J.P.A.; Figueiredo, S.; Castelo-Branco, R.; Freitas, S. Incorporation and Modification of Fatty Acids in Cyanobacterial Natural Products Biosynthesis. Chem. Commun. 2023, 59, 4436–4446. [Google Scholar] [CrossRef] [PubMed]
- Shiels, K.; Tsoupras, A.; Lordan, R.; Zabetakis, I.; Murray, P.; Kumar Saha, S. Anti-Inflammatory and Antithrombotic Properties of Polar Lipid Extracts, Rich in Unsaturated Fatty Acids, from the Irish Marine Cyanobacterium Spirulina subsalsa. J. Funct. Foods 2022, 94, 105124. [Google Scholar] [CrossRef]
- Demay, J.; Bernard, C.; Reinhardt, A.; Marie, B. Natural Products from Cyanobacteria: Focus on Beneficial Activities. Mar. Drugs 2019, 17, 320. [Google Scholar] [CrossRef]
- Brumley, D.A.; Gunasekera, S.P.; Sauvage, T.; Dos Santos, L.A.H.; Chen, Q.Y.; Paul, V.J.; Luesch, H. Discovery, Synthesis, and Biological Evaluation of Anaenamides C and D from a New Marine Cyanobacterium, Hormoscilla sp. J. Nat. Prod. 2022, 85, 581–589. [Google Scholar] [CrossRef]
- Mascuch, S.J.; Boudreau, P.D.; Carland, T.M.; Pierce, N.T.; Olson, J.; Hensler, M.E.; Choi, H.; Campanale, J.; Hamdoun, A.; Nizet, V.; et al. Marine Natural Product Honaucin A Attenuates Inflammation by Activating the Nrf2-ARE Pathway. J. Nat. Prod. 2018, 81, 506–514. [Google Scholar] [CrossRef]
- Villa, F.A.; Lieske, K.; Gerwick, L. Selective MyD88-Dependent Pathway Inhibition by the Cyanobacterial Natural Product Malyngamide F Acetate. Eur. J. Pharmacol. 2010, 629, 140–146. [Google Scholar] [CrossRef]
- Fontana, A.; D’Ippolito, G.; Cutignano, A.; Romano, G.; Lamari, N.; Gallucci, A.M.; Cimino, G.; Miralto, A.; Lanora, A. LOX-Induced Lipid Peroxidation Mechanism Responsible for the Detrimental Effect of Marine Diatoms on Zooplankton Grazers. ChemBioChem 2007, 8, 1810–1818. [Google Scholar] [CrossRef] [PubMed]
- Moldes-Anaya, A.; Sæther, T.; Uhlig, S.; Nebb, H.I.; Larsen, T.; Eilertsen, H.C.; Paulsen, S.M. Two Isomeric C16 Oxo-Fatty Acids from the Diatom Chaetoceros karianus Show Dual Agonist Activity towards Human Peroxisome Proliferator-Activated Receptors (PPARs) α/γ. Mar. Drugs 2017, 15, 148. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.D.; Hye, J.L.; Eun, S.Y.; Shinde, P.B.; Yoon, M.L.; Hong, J.; Dong, K.K.; Jung, J.H. Anti-Inflammatory Constituents of the Red Alga Gracilaria verrucosa and Their Synthetic Analogues. J. Nat. Prod. 2008, 71, 232–240. [Google Scholar]
- D’Ippolito, G.; Cutignano, A.; Briante, R.; Febbraio, F.; Cimino, G.; Fontana, A. New C16 Fatty-Acid-Based Oxylipin Pathway in the Marine Diatom Thalassiosira rotula. Org. Biomol. Chem. 2005, 3, 4065–4070. [Google Scholar] [CrossRef] [PubMed]
- Santalova, E.A.; Denisenko, V.A. Analysis of the Configuration of an Isolated Double Bond in Some Lipids by Selective Homonuclear Decoupling. Nat. Prod. Commun. 2017, 12, 1913–1916. [Google Scholar] [CrossRef]
- Li, D.; Wu, M. Pattern Recognition Receptors in Health and Diseases. Signal Transduct. Target. Ther. 2021, 6, 291. [Google Scholar] [CrossRef]
- Moreira, L.O.; Zamboni, D.S. NOD1 and NOD2 Signaling in Infection and Inflammation. Front. Immunol. 2012, 3, 328. [Google Scholar] [CrossRef]
- Mukherjee, T.; Hovingh, E.S.; Foerster, E.G.; Abdel-Nour, M.; Philpott, D.J.; Girardin, S.E. NOD1 and NOD2 in Inflammation, Immunity and Disease. Arch. Biochem. Biophys. 2019, 670, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Kang, Z.; Liu, C.; Li, X. IL-17 Signaling in Host Defense and Inflammatory Diseases. Cell. Mol. Immunol. 2010, 7, 328–333. [Google Scholar] [CrossRef]
- Ruiz de Morales, J.M.G.; Puig, L.; Daudén, E.; Cañete, J.D.; Pablos, J.L.; Martín, A.O.; Juanatey, C.G.; Adán, A.; Montalbán, X.; Borruel, N.; et al. Critical Role of Interleukin (IL)-17 in Inflammatory and Immune Disorders: An Updated Review of the Evidence Focusing in Controversies. Autoimmun. Rev. 2020, 19, 102429. [Google Scholar] [CrossRef]
- Yi, Y.S.; Son, Y.J.; Ryou, C.; Sung, G.H.; Kim, J.H.; Cho, J.Y. Functional Roles of P38 Mitogen-Activated Protein Kinase in Macrophage-Mediated Inflammatory Responses. Mediators Inflamm. 2014, 2014, 352371. [Google Scholar] [CrossRef]
- Magna, M.; Pisetsky, D.S. The Role of HMGB1 in the Pathogenesis of Inflammatory and Autoimmune Diseases. Mol. Med. 2014, 20, 138–146. [Google Scholar] [CrossRef]
- Tyagi, S.; Gupta, P.; Saini, A.; Kaushal, C.; Sharma, S. The Peroxisome Proliferator-Activated Receptor: A Family of Nuclear Receptors Role in Various Diseases. J. Adv. Pharm. Technol. Res. 2011, 2, 236. [Google Scholar] [CrossRef] [PubMed]
- Crinelli, R.; Antonelli, A.; Bianchi, M.; Gentilini, L.; Scaramucci, S.; Magnani, M. Selective Inhibition of NF-κB Activation and TNF-α Production in Macrophages by Red Blood Cell-Mediated Delivery of Dexamethasone. Blood Cells Mol. Dis. 2000, 26, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.J.; Han, S.H.; Lee, Y.W.; Lee, M.; Yang, K.H.; Kim, H.M. Dexamethasone Inhibits IL-1β Gene Expression in LPS-Stimulated RAW 264.7 Cells by Blocking NF-κB/Rel and AP-1 Activation. Immunopharmacology 2000, 48, 173–183. [Google Scholar] [CrossRef]
- Kent, L.M.; Smyth, L.J.C.; Plumb, J.; Clayton, C.L.; Fox, S.M.; Ray, D.W.; Farrow, S.N.; Singh, D. Inhibition of Lipopolysaccharide-Stimulated Chronic Obstructive Pulmonary Disease Macrophage Inflammatory Gene Expression by Dexamethasone and the P38 Mitogen-Activated Protein Kinase Inhibitor N-Cyano-N′-(2-{[8-(2,6- Difluorophenyl)-4-(4-Fluoro-2-Methylphenyl)-7-oxo-7,8-dihydropyrido[2,3-d] pyrimidin-2-yl]amino}ethyl)guanidine (SB706504). J. Pharmacol. Exp. Ther. 2009, 328, 458–468. [Google Scholar] [PubMed]
- Zhao, Y.; Shen, X.F.; Cao, K.; Ding, J.; Kang, X.; Guan, W.X.; Ding, Y.T.; Liu, B.R.; Du, J.F. Dexamethasone-Induced Myeloid-Derived Suppressor Cells Prolong Allo Cardiac Graft Survival through INOS- and Glucocorticoid Receptor-Dependent Mechanism. Front. Immunol. 2018, 9, 282. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, A.A.; Tan, X.; Reis, J.C.; Badr, M.Z.; Papasian, C.J.; Morrison, D.C.; Qureshi, N. Inhibition of Nitric Oxide in LPS-Stimulated Macrophages of Young and Senescent Mice by δ-Tocotrienol and Quercetin. Lipids Health Dis. 2011, 10, 239. [Google Scholar] [CrossRef]
- Golde, S.; Coles, A.; Lindquist, J.A.; Compston, A. Decreased INOS Synthesis Mediates Dexamethasone-Induced Protection of Neurons from Inflammatory Injury in Vitro. Eur. J. Neurosci. 2003, 18, 2527–2537. [Google Scholar] [CrossRef] [PubMed]
- Walker, G.; Pfeilschifter, J.; Kunz, D. Mechanisms of Suppression of Inducible Nitric-Oxide Synthase (INOS) Expression in Interferon (IFN)-γ-Stimulated RAW 264.7 Cells by Dexamethasone. Evidence for Glucocorticoid-Induced Degradation of INOS Protein by Calpain as a Key Step in Post-Transcriptional Regulation. J. Biol. Chem. 1997, 272, 16679–16687. [Google Scholar]
- Kunz, D.; Walker, G.; Eberhardt, W.; Pfeilschifter, J. Molecular Mechanisms of Dexamethasone Inhibition of Nitric Oxide Synthase Expression in Interleukin 1β-Stimulated Mesangial Cells: Evidence for the Involvement of Transcriptional and Posttranscriptional Regulation. Proc. Natl. Acad. Sci. USA 1996, 93, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, R.; Lahti, A.; Hämäläinen, M.; Kankaanranta, H.; Moilanen, E. Dexamethasone Inhibits Inducible Nitric-Oxide Synthase Expression and Nitric Oxide Production by Destabilizing MRNA in Lipopolysaccharide-Treated Macrophages. Mol. Pharmacol. 2002, 62, 698–704. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Han, S.H.; Lee, Y.W.; Yea, S.S.; Yang, K.H. Inhibition of NF-Kappa B/Rel Nuclear Translocation by Dexamethasone: Mechanism for the Inhibition of INOS Gene Expression. Biochem. Mol. Biol. Int. 1998, 45, 435–441. [Google Scholar] [PubMed]
- Nair, S.; Doh, S.T.; Chan, J.Y.; Kong, A.N.; Cai, L. Regulatory Potential for Concerted Modulation of Nrf2- and Nfkb1-Mediated Gene Expression in Inflammation and Carcinogenesis. Br. J. Cancer 2008, 99, 2070–2082. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.F.; Kuo, H.P.; Liu, M.; Chou, C.K.; Xia, W.; Du, Y.; Shen, J.; Chen, C.T.; Huo, L.; Hsu, M.C.; et al. KEAP1 E3 Ligase-Mediated Downregulation of NF-κB Signaling by Targeting IKKβ. Mol. Cell 2009, 36, 131–140. [Google Scholar] [CrossRef]
- Alcaraz, M.J.; Vicente, A.M.; Araico, A.; Dominguez, J.N.; Terencio, M.C.; Ferrándiz, M.L. Role of Nuclear Factor-κB and Heme Oxygenase-1 in the Mechanism of Action of an Anti-Inflammatory Chalcone Derivative in RAW 264.7 Cells. Br. J. Pharmacol. 2004, 142, 1191–1199. [Google Scholar] [CrossRef]
- Jiménez-Villegas, J.; Ferraiuolo, L.; Mead, R.J.; Shaw, P.J.; Cuadrado, A.; Rojo, A.I. NRF2 as a Therapeutic Opportunity to Impact in the Molecular Roadmap of ALS. Free Radic. Biol. Med. 2021, 173, 125–141. [Google Scholar] [CrossRef]
- Pournajaf, S.; Dargahi, L.; Javan, M.; Pourgholami, M.H. Molecular Pharmacology and Novel Potential Therapeutic Applications of Fingolimod. Front. Pharmacol. 2022, 13, 352371. [Google Scholar] [CrossRef]
- Colombo, E.; Di Dario, M.; Capitolo, E.; Chaabane, L.; Newcombe, J.; Martino, G.; Farina, C. Fingolimod May Support Neuroprotection via Blockade of Astrocyte Nitric Oxide. Ann. Neurol. 2014, 76, 325–337. [Google Scholar] [CrossRef]
- Meng, T.; Xiao, D.; Muhammed, A.; Deng, J.; Chen, L.; He, J. Anti-Inflammatory Action and Mechanisms of Resveratrol. Molecules 2021, 26, 229. [Google Scholar] [CrossRef]
- Abou-ElWafa, G.S.E.; Shaaban, M.; Shaaban, K.A.; El-Naggar, M.E.E.; Laatsch, H. Three New Unsaturated Fatty Acids from the Marine Green Alga Ulva fasciata Delile. Z. Naturforschung B 2009, 64, 1199–1207. [Google Scholar] [CrossRef]
- Van der Horst, D.; Carter-Timofte, M.E.; van Grevenynghe, J.; Laguette, N.; Dinkova-Kostova, A.T.; Olagnier, D. Regulation of Innate Immunity by Nrf2. Curr. Opin. Immunol. 2022, 78, 102247. [Google Scholar] [CrossRef] [PubMed]
- Brandes, M.S.; Gray, N.E. NRF2 as a Therapeutic Target in Neurodegenerative Diseases. ASN Neuro 2020, 12, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Michaličková, D.; Hrnčíř, T.; Canová, N.K.; Slanař, O. Targeting Keap1/Nrf2/ARE Signaling Pathway in Multiple Sclerosis. Eur. J. Pharmacol. 2020, 873, 172973. [Google Scholar] [CrossRef]
- Maldonado, P.P.; Guevara, C.; Olesen, M.A.; Orellana, J.A.; Quintanilla, R.A.; Ortiz, F.C. Neurodegeneration in Multiple Sclerosis: The Role of Nrf2-Dependent Pathways. Antioxidants 2022, 11, 1146. [Google Scholar] [CrossRef]
- Hammer, A.; Waschbisch, A.; Kuhbandner, K.; Bayas, A.; Lee, D.H.; Duscha, A.; Haghikia, A.; Gold, R.; Linker, R.A. The NRF2 Pathway as Potential Biomarker for Dimethyl Fumarate Treatment in Multiple Sclerosis. Ann. Clin. Transl. Neurol. 2018, 5, 668–676. [Google Scholar] [CrossRef]
- Al-Awadhi, F.H.; Paul, V.J.; Luesch, H. Structural Diversity and Anticancer Activity of Marine-Derived Elastase Inhibitors: Key Features and Mechanisms Mediating the Antimetastatic Effects in Invasive Breast Cancer. ChemBioChem 2018, 19, 815. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C/H No. | 1 | 2 | 3 | 4 | ||
|---|---|---|---|---|---|---|
| δH (J in Hz) | δC a, mult b | δH (J in Hz) | δC a, mult b | δH (J in Hz) | δH (J in Hz) | |
| OH | − | − | − | − | − | − |
| 1 | − | 177.2, qC | − | 176.6, qC | − | − |
| 2 | 2.35, t (7.4) | 33.7, CH2 | 2.37, t (7.4) | 33.5, CH2 | 2.37, t (7.4) | 2.37, t (7.4) |
| 3 | 1.65, m | 24.8, CH2 | 1.67, m | 24.5, CH2 | 1.67, m | 1.67, m |
| 4 | 1.31, m | 31.7, CH2 | 1.37, m | 28.7, CH2 | 1.32, m | 1.28, m |
| 5 | 1.49, m | 27.9, CH2 | 1.40, m | 28.7, CH2 | 1.32, m | 1.28, m |
| 6 | 2.21, dt (7.1, 6.9) | 32.3, CH2 | 2.05, dt (7.1, 6.9) | 32.1, CH2 | 1.32, m | 1.28, m |
| 7 | 6.83, dt (15.3, 6.9) | 147.1, CH | 5.63, dt (15.3, 6.9) | 132.1, CH | 1.32, m | 1.28, m |
| 8 | 6.11, dd (15.3, 7.1) | 130.3, CH | 5.47, dd (15.3, 7.1) | 133.2, CH | 2.02, dt (6.9, 6.3) | 1.28, m |
| 9 | − | 201.0, qC | 4.06, dt (7.1, 6.8) | 73.3, CH | 5.34, m | 1.28, m |
| 9-OH | − | − | − | − | ||
| 10 | 2.54, m | 40.2, CH2 | 1.57, 1.49, m | 37.3, CH2 | 5.34, m | 1.28, m |
| 11 | 1.62, m | 24.2, CH2 | 1.33, m | 25.4, CH2 | 2.02, dt (6.9, 6.3) | 1.28, m |
| 12, 13 | 1.37, m | 28.7, CH2 | 1.30, m | 29.1, CH2 | 1.32, m | 1.28, m |
| 14 | 1.37, m | 28.7, CH2 | 1.30, m | 31.9, CH2 | 1.32, m | 1.28, m |
| 15 | 1.32, m | 22.5, CH2 | 1.31, m | 22.5, CH2 | 1.32, m | 1.28, m |
| 16 | 0.9, t (7.0) | 14.1, CH3 | 0.9, t (7.0) | 14.1, CH3 | 0.9, t (7.0) | 0.9, t (7.0) |
| Canonical Pathway | Activation z-Score | p-Value | No. of Molecules | Selected Genes |
|---|---|---|---|---|
| Role of hypercytokinemia/hyperchemokinemia in the pathogenesis of Influenza | −5.112 | 4.85 × 10−27 | 30 | IL33, CCL2, IFNB1, IL1A, IFIT3 |
| Pathogen-induced cytokine storm signaling pathway | −5.091 | 4.85 × 10−23 | 50 | IL33, CCL2, CXCL9, CSF2, NOS2 |
| Role of pattern recognition receptors in recognition of bacteria and viruses | −2.887 | 1.10 × 10−17 | 29 | IL33, CSF2, TNFSF10, IFNB1, IL1A |
| Multiple sclerosis signaling pathway | −3.402 | 1.03 × 10−12 | 28 | IL33, CSF2, TNFSF10, IL1A, IL36A |
| NOD1/2 signaling pathway | −3.138 | 8.86 × 10−13 | 26 | IL33, CCL2, CSF2, TNFSF10, NOS2 |
| TREM1 signaling | −3.638 | 3.42 × 10−12 | 17 | CCL2, CSF2, CD86, IL6, IL1B |
| Macrophage classical activation signaling pathway | −3.400 | 5.92 × 10−12 | 25 | IL33, CSF2, TNFSF10, NOS2 |
| IL-17 signaling | −4.146 | 6.02 × 10−09 | 21 | IL33, CCL2, MMP3, CSF2, NOS2 |
| Interferon signaling | −3.162 | 9.25 × 10−09 | 10 | IFIT1 *, IFNB1, IFIT3 *, ISG15 |
| Neuroinflammation signaling pathway | −2.449 | 1.84 × 10−08 | 27 | CCL2, MMP3, NOX1, NOS2 |
| Differential regulation of cytokine production in macrophages and T-helper cells | −2.646 | 1.19 × 10−07 | 7 | CCL2, CSF2, IL6, IL1B, CCL4 |
| LXR/RXR activation | 2.840 | 3.04 × 10−07 | 15 | IL33, CCL2, NOS2, IL1A, IL36A |
| Toll-like receptor signaling | −2.111 | 3.68 × 10−07 | 12 | IL33, IL1A, IL36A, IL1B, TLR8 |
| LPS-IL-1-mediated inhibition of RXR function | −2.000 | 1.81 × 10−05 | 19 | IL33, IL1A, IL36A, IL1B, FABP4 |
| Glutathione-mediated detoxification * | 1.633 | 2.58 × 10−05 | 7 | Gsta4, GSTA5 *, GSTA3, MGST2 |
| HMGB1 signaling | −2.646 | 5.89 × 10−05 | 14 | IL33, CCL2, CSF2, TNFSF10, IL1A |
| P38 MAPK signaling | −2.530 | 1.64 × 10−04 | 11 | IL33, IL1A, IL36A, IL1B, PLA2G4C |
| PPAR signaling | 1.265 | 2.75 × 10−05 | 10 | IL33, IL1A, IL36A, IL1B |
| NRF2-mediated oxidative stress response * | 1.414 | 2.22 × 10−03 | 14 | GSTA5, GSTA3, NQO1, PRDX1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Awadhi, F.H.; Simon, E.F.; Liu, N.; Ratnayake, R.; Paul, V.J.; Luesch, H. Discovery and Anti-Inflammatory Activity of a Cyanobacterial Fatty Acid Targeting the Keap1/Nrf2 Pathway. Mar. Drugs 2023, 21, 553. https://doi.org/10.3390/md21110553
Al-Awadhi FH, Simon EF, Liu N, Ratnayake R, Paul VJ, Luesch H. Discovery and Anti-Inflammatory Activity of a Cyanobacterial Fatty Acid Targeting the Keap1/Nrf2 Pathway. Marine Drugs. 2023; 21(11):553. https://doi.org/10.3390/md21110553
Chicago/Turabian StyleAl-Awadhi, Fatma H., Emily F. Simon, Na Liu, Ranjala Ratnayake, Valerie J. Paul, and Hendrik Luesch. 2023. "Discovery and Anti-Inflammatory Activity of a Cyanobacterial Fatty Acid Targeting the Keap1/Nrf2 Pathway" Marine Drugs 21, no. 11: 553. https://doi.org/10.3390/md21110553
APA StyleAl-Awadhi, F. H., Simon, E. F., Liu, N., Ratnayake, R., Paul, V. J., & Luesch, H. (2023). Discovery and Anti-Inflammatory Activity of a Cyanobacterial Fatty Acid Targeting the Keap1/Nrf2 Pathway. Marine Drugs, 21(11), 553. https://doi.org/10.3390/md21110553