

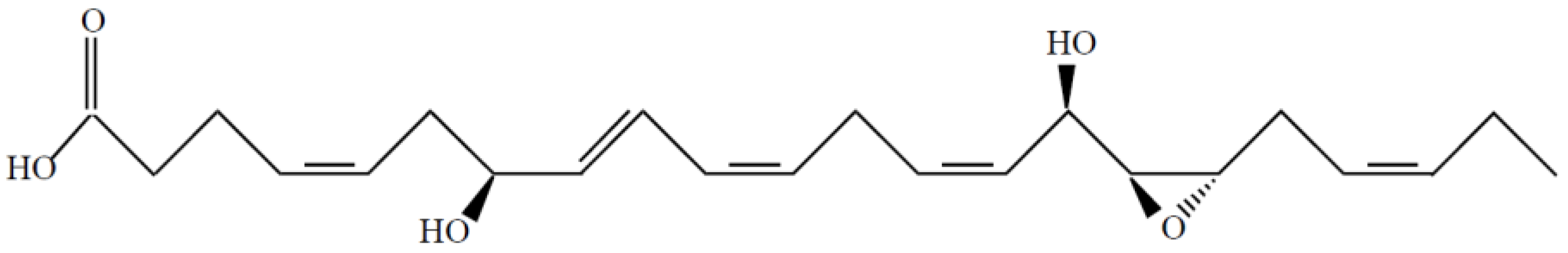
7S,15R-Dihydroxy-16S,17S-epoxy-docosapentaenoic Acid Overcomes Chemoresistance of 5-Fluorouracil by Suppressing the Infiltration of Tumor-Associated Macrophages and Inhibiting the Activation of Cancer Stem Cells in a Colorectal Cancer Xenograft Model

 and
and
Abstract
:1. Introduction
2. Results
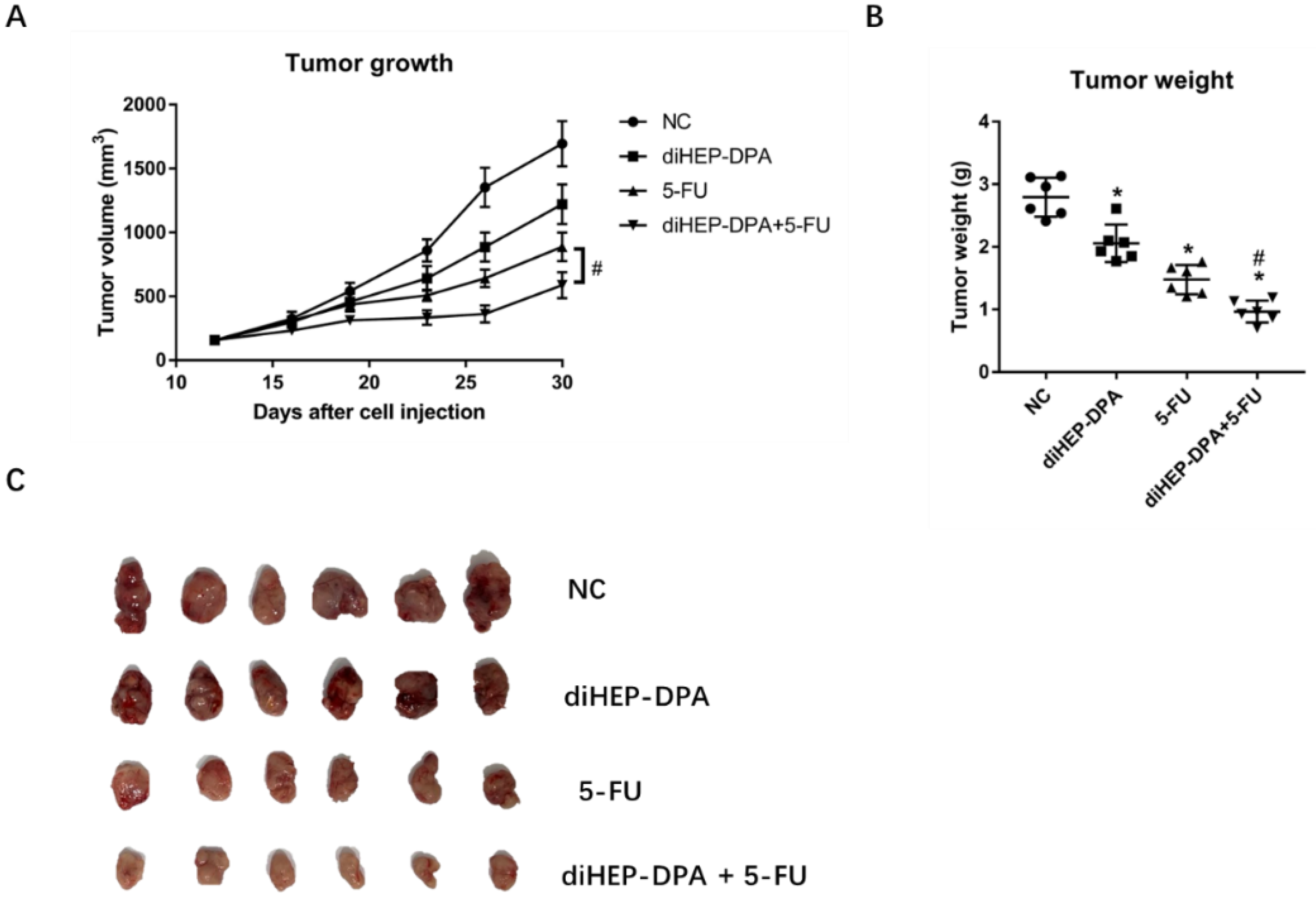
2.1. diHEP-DPA and 5-FU Inhibited Tumor Growth in the CRC Xenograft Model
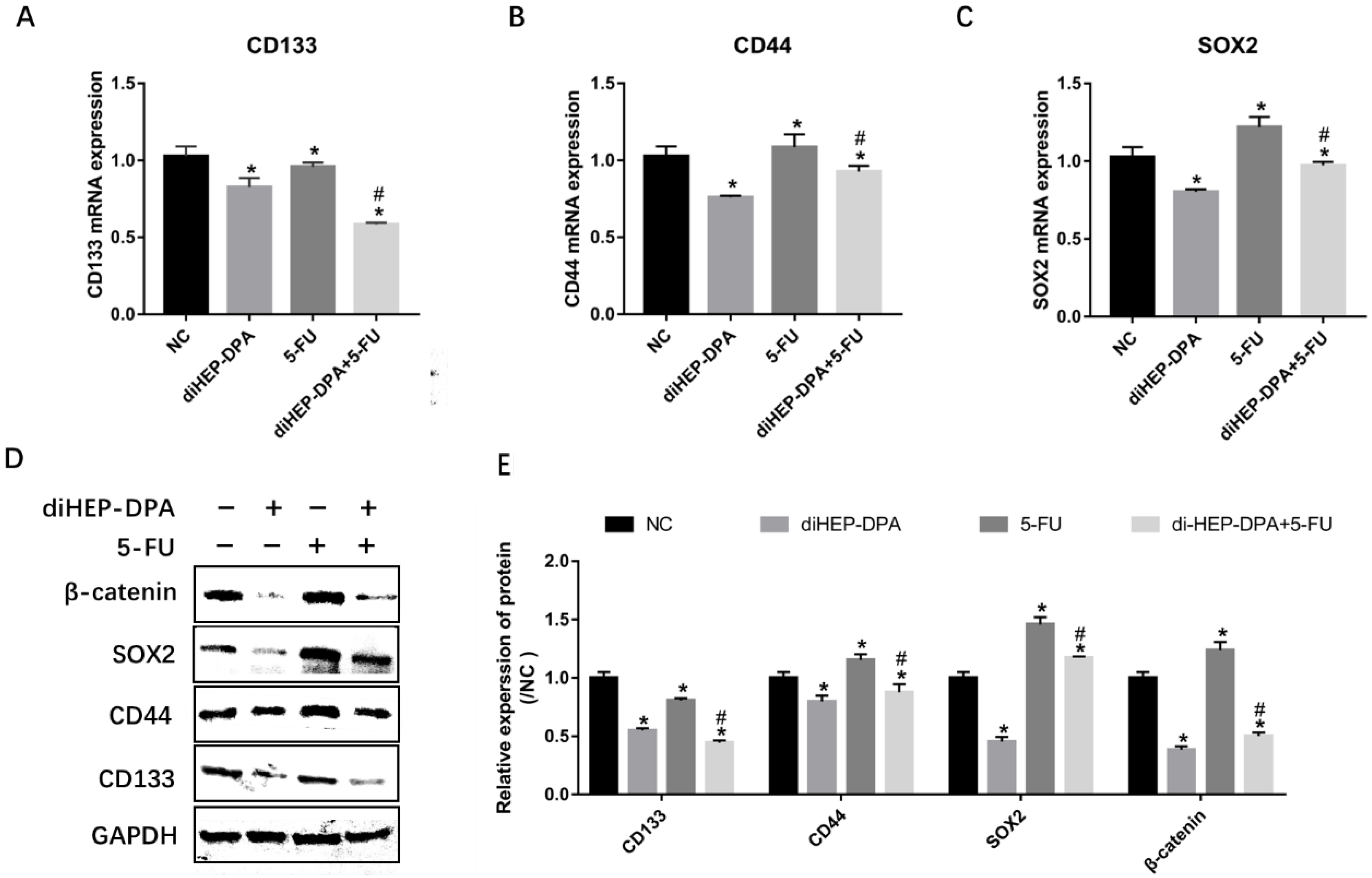
2.2. diHEP-DPA Suppressed 5-FU-Induced CSCs Activation via the WNT/β-Catenin Pathway
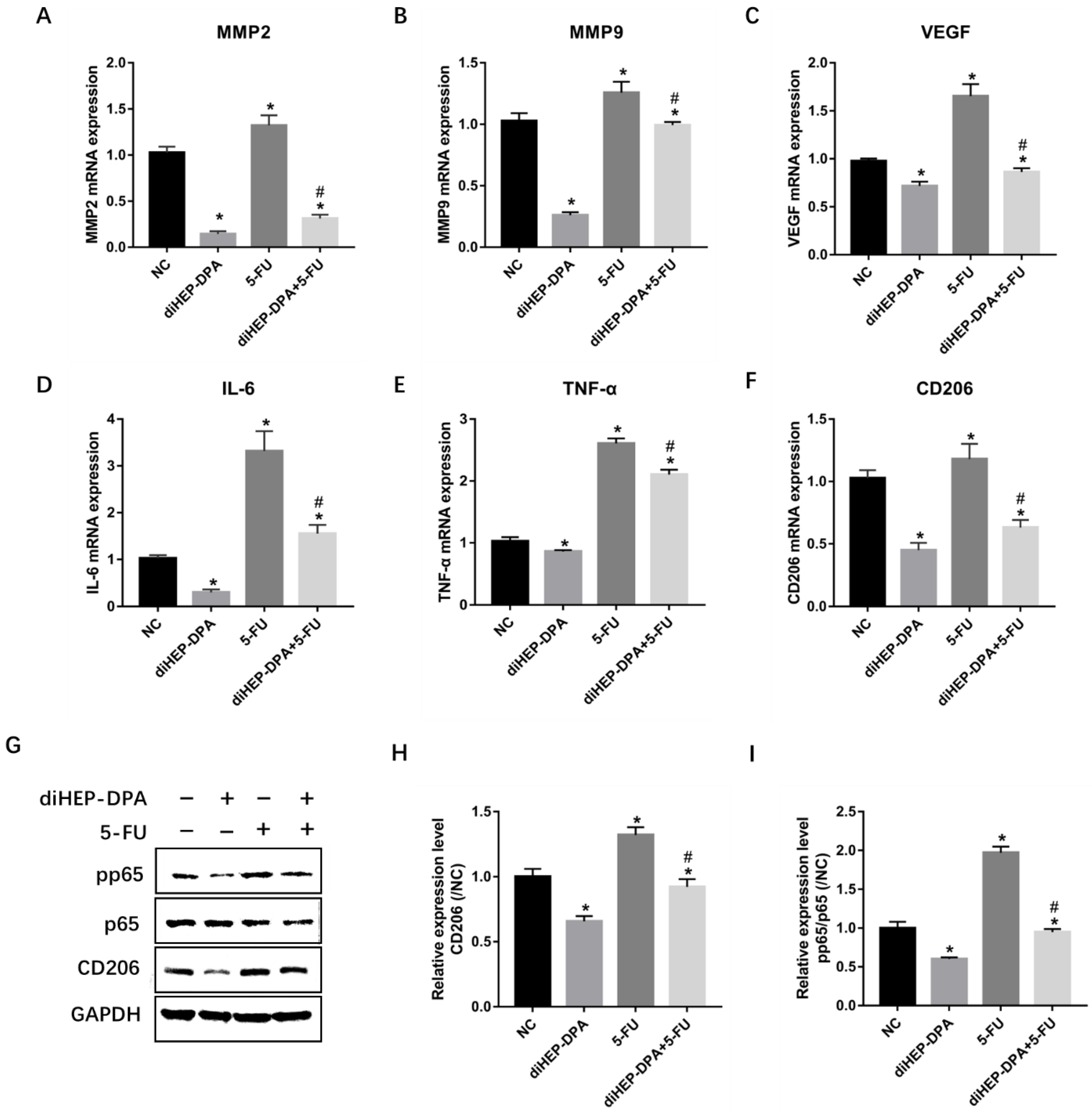
2.3. diHEP-DPA Inhibited 5-FU-Induced Infiltration of M2-likeTAMs
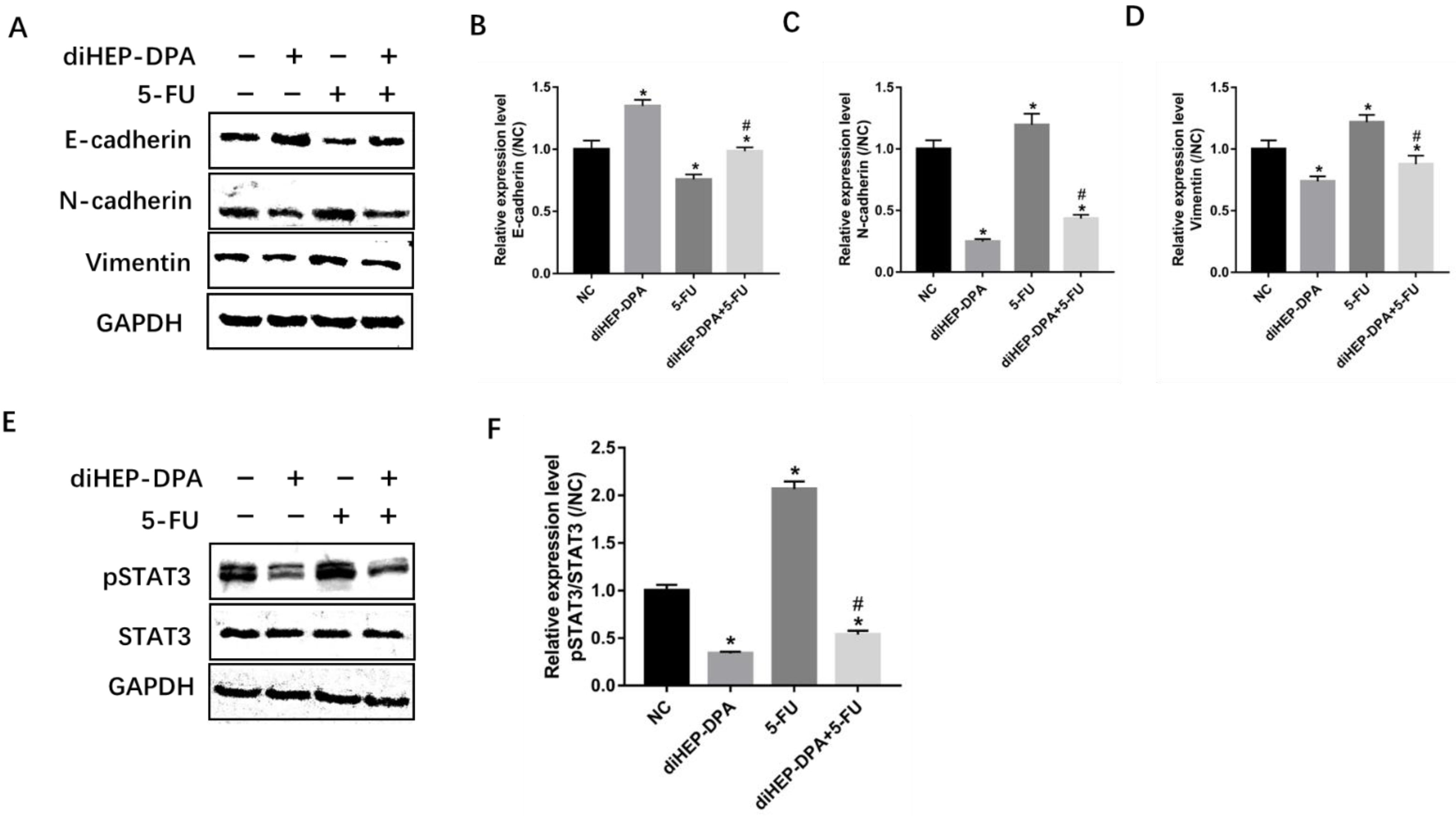
2.4. diHEP-DPA Impeded EMT via the STAT3 Signaling Pathway
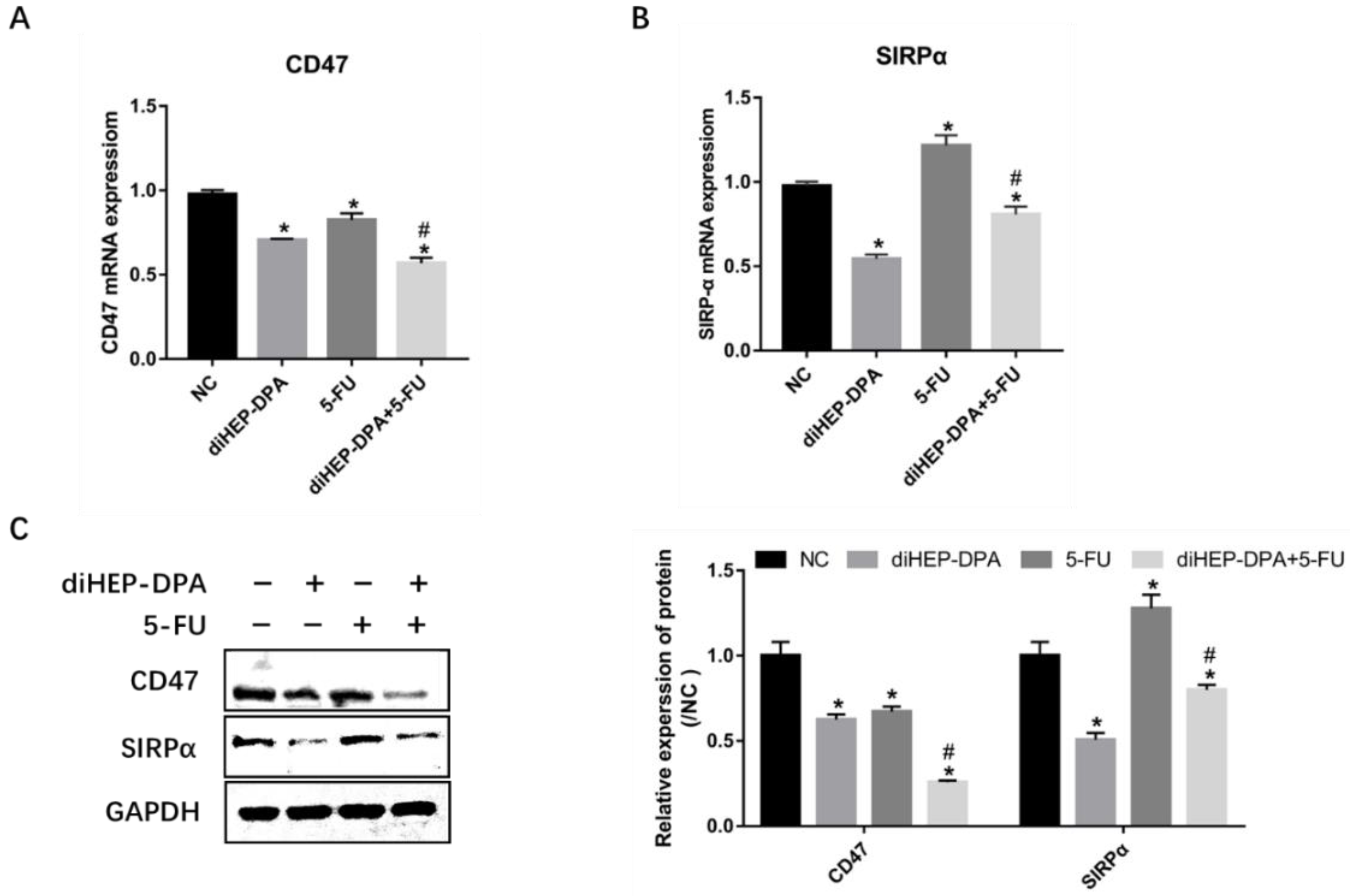
2.5. diHEP-DPA and 5-FU Enhanced Macrophage Phagocytic Activity via CD47/SIRPα
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals and Cell Culture
4.3. CRC Xenograft Model in BALB/c Mice
4.4. Quantitative Reverse Transcription PCR (qRT-PCR)
4.5. PROTEIN Preparation and Western Blotting
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Blondy, S.; David, V.; Verdier, M.; Mathonnet, M.; Perraud, A.; Christou, N. 5-Fluorouracil Resistance Mechanisms in Colorectal Cancer: From Classical Pathways to Promising Processes. Cancer Sci. 2020, 111, 3142–3154. [Google Scholar] [CrossRef]
- Dean, M.; Fojo, T.; Bates, S. Tumour Stem Cells and Drug Resistance. Nat. Rev. Cancer 2005, 5, 275–284. [Google Scholar] [CrossRef]
- Erin, N.; Grahovac, J.; Brozovic, A.; Efferth, T. Tumor Microenvironment and Epithelial Mesenchymal Transition as Targets to Overcome Tumor Multidrug Resistance. Drug Resist. Updates 2020, 53, 100715. [Google Scholar] [CrossRef]
- Sethy, C.; Kundu, C.N. 5-Fluorouracil (5-FU) Resistance and the New Strategy to Enhance the Sensitivity against Cancer: Implication of DNA Repair Inhibition. Biomed. Pharmacother. 2021, 137, 111285. [Google Scholar] [CrossRef]
- Acharyya, S.; Oskarsson, T.; Vanharanta, S.; Malladi, S.; Kim, J.; Morris, P.G.; Manova-Todorova, K.; Leversha, M.; Hogg, N.; Seshan, V.E.; et al. A CXCL1 Paracrine Network Links Cancer Chemoresistance and Metastasis. Cell 2012, 150, 165–178. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Chen, Y.; Hao, L.; Hou, A.; Chen, X.; Li, Y.; Wang, R.; Luo, P.; Ruan, Z.; Ou, J.; et al. Macrophages Induce Resistance to 5-Fluorouracil Chemotherapy in Colorectal Cancer through the Release of Putrescine. Cancer Lett. 2016, 381, 305–313. [Google Scholar] [CrossRef]
- Azwar, S.; Seow, H.F.; Abdullah, M.; Jabar, M.F.; Mohtarrudin, N. Recent Updates on Mechanisms of Resistance to 5-Fluorouracil and Reversal Strategies in Colon Cancer Treatment. Biology 2021, 10, 854. [Google Scholar] [CrossRef]
- Shih, J.-Y.; Yuan, A.; Chen, J.J.-W.; Yang, P.-C. Tumor-Associated Macrophage: Its Role in Cancer Invasion and Metastasis. J. Cancer Mol. 2006, 2, 101–106. [Google Scholar]
- Ding, L.; Liang, G.; Yao, Z.; Zhang, J.; Liu, R.; Chen, H.; Zhou, Y.; Wu, H.; Yang, B.; He, Q. Metformin Prevents Cancer Metastasis by Inhibiting M2-like Polarization of Tumor Associated Macrophages. Oncotarget 2015, 6, 36441–36455. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Liang, F.; Cheng, W.; Zhou, R.; Wu, X.; Feng, Y.; Wang, Y. The Mechanisms for Lung Cancer Risk of PM2.5: Induction of Epithelial-Mesenchymal Transition and Cancer Stem Cell Properties in Human Non-Small Cell Lung Cancer Cells. Environ. Toxicol. 2017, 32, 2341–2351. [Google Scholar] [CrossRef]
- Wang, W.; Zhao, Y.; Yao, S.; Cui, X.; Pan, W.; Huang, W.; Gao, J.; Dong, T.; Zhang, S. Nigericin Inhibits Epithelial Ovarian Cancer Metastasis by Suppressing the Cell Cycle and Epithelial−mesenchymal Transition. Biochem. Mosc. 2017, 82, 933–941. [Google Scholar] [CrossRef]
- Kong, L.; Zhou, Y.; Bu, H.; Lv, T.; Shi, Y.; Yang, J. Deletion of Interleukin-6 in Monocytes/Macrophages Suppresses the Initiation of Hepatocellular Carcinoma in Mice. J. Exp. Clin. Cancer Res. 2016, 35, 131. [Google Scholar] [CrossRef] [Green Version]
- Storr, S.J.; Safuan, S.; Ahmad, N.; El-Refaee, M.; Jackson, A.M.; Martin, S.G. Macrophage-Derived Interleukin-1beta Promotes Human Breast Cancer Cell Migration and Lymphatic Adhesion in Vitro. Cancer Immunol. Immunother. 2017, 66, 1287–1294. [Google Scholar] [CrossRef] [Green Version]
- Willingham, S.B.; Volkmer, J.-P.; Gentles, A.J.; Sahoo, D.; Dalerba, P.; Mitra, S.S.; Wang, J.; Contreras-Trujillo, H.; Martin, R.; Cohen, J.D.; et al. The CD47-Signal Regulatory Protein Alpha (SIRPa) Interaction Is a Therapeutic Target for Human Solid Tumors. Proc. Natl. Acad. Sci. USA 2012, 109, 6662–6667. [Google Scholar] [CrossRef] [Green Version]
- Arrieta, O.; Aviles-Salas, A.; Orozco-Morales, M.; Hernández-Pedro, N.; Cardona, A.F.; Cabrera-Miranda, L.; Barrios-Bernal, P.; Soca-Chafre, G.; Cruz-Rico, G.; de Lourdes Peña-Torres, M.; et al. Association between CD47 Expression, Clinical Characteristics and Prognosis in Patients with Advanced Non-small Cell Lung Cancer. Cancer Med. 2020, 9, 2390–2402. [Google Scholar] [CrossRef]
- Cherciu, I.; Bărbălan, A.; Pirici, D.; Mărgăritescu, C.; Săftoiu, A. Stem Cells, Colorectal Cancer and Cancer Stem Cell Markers Correlations. Curr. Health Sci. J. 2014, 40, 153–161. [Google Scholar] [CrossRef]
- Eppert, K.; Takenaka, K.; Lechman, E.R.; Waldron, L.; Nilsson, B.; van Galen, P.; Metzeler, K.H.; Poeppl, A.; Ling, V.; Beyene, J.; et al. Stem Cell Gene Expression Programs Influence Clinical Outcome in Human Leukemia. Nat. Med. 2011, 17, 1086–1093. [Google Scholar] [CrossRef]
- Cho, Y.-H.; Ro, E.J.; Yoon, J.-S.; Mizutani, T.; Kang, D.-W.; Park, J.-C.; Il Kim, T.; Clevers, H.; Choi, K.-Y. 5-FU Promotes Stemness of Colorectal Cancer via P53-Mediated WNT/β-Catenin Pathway Activation. Nat. Commun. 2020, 11, 5321. [Google Scholar] [CrossRef]
- Wang, L.F.; Choi, H.S.; Su, Y.; Lee, B.; Song, J.J.; Jang, Y.-S.; Seo, J.-W. 7S,15R-Dihydroxy-16S,17S-Epoxy-Docosapentaenoic Acid, a Novel DHA Epoxy Derivative, Inhibits Colorectal Cancer Stemness through Repolarization of Tumor-Associated Macrophage Functions and the ROS/STAT3 Signaling Pathway. Antioxidants 2021, 10, 1459. [Google Scholar] [CrossRef]
- Wang, K.; Song, K.; Ma, Z.; Yao, Y.; Liu, C.; Yang, J.; Xiao, H.; Zhang, J.; Zhang, Y.; Zhao, W. Identification of EMT-Related High-Risk Stage II Colorectal Cancer and Characterisation of Metastasis-Related Genes. Br. J. Cancer 2020, 123, 410–417. [Google Scholar] [CrossRef]
- Yang, Y.; Ma, L.; Xu, Y.; Liu, Y.; Li, W.; Cai, J.; Zhang, Y. Enalapril Overcomes Chemoresistance and Potentiates Antitumor Efficacy of 5-FU in Colorectal Cancer by Suppressing Proliferation, Angiogenesis, and NF-ΚB/STAT3-Regulated Proteins. Cell Death Dis. 2020, 11, 477. [Google Scholar] [CrossRef]
- Liu, H.; Ren, G.; Wang, T.; Chen, Y.; Gong, C.; Bai, Y.; Wang, B.; Qi, H.; Shen, J.; Zhu, L.; et al. Aberrantly Expressed Fra-1 by IL-6/STAT3 Transactivation Promotes Colorectal Cancer Aggressiveness through Epithelial–Mesenchymal Transition. Carcinogenesis 2015, 36, 459–468. [Google Scholar] [CrossRef] [Green Version]
- Oldenborg, P.-A.; Zheleznyak, A.; Fang, Y.-F.; Lagenaur, C.F.; Gresham, H.D.; Lindberg, F.P. Role of CD47 as a Marker of Self on Red Blood Cells. Science 2000, 288, 2051–2054. [Google Scholar] [CrossRef]
- der Jeught, K.V.; Xu, H.-C.; Li, Y.-J.; Lu, X.-B.; Ji, G. Drug Resistance and New Therapies in Colorectal Cancer. World J. Gastroenterol. 2018, 24, 3834–3848. [Google Scholar] [CrossRef]
- Belgiovine, C.; D’Incalci, M.; Allavena, P.; Frapolli, R. Tumor-Associated Macrophages and Anti-Tumor Therapies: Complex Links. Cell. Mol. Life Sci. 2016, 73, 2411–2424. [Google Scholar] [CrossRef]
- Shmelkov, S.V.; Butler, J.M.; Hooper, A.T.; Hormigo, A.; Kushner, J.; Milde, T.; St. Clair, R.; Baljevic, M.; White, I.; Jin, D.K.; et al. CD133 Expression Is Not Restricted to Stem Cells, and Both CD133+ and CD133– Metastatic Colon Cancer Cells Initiate Tumors. J. Clin. Invest. 2008, 118, 2111–2120. [Google Scholar] [CrossRef]
- Fan, Z.; Duan, J.; Wang, L.; Xiao, S.; Li, L.; Yan, X.; Yao, W.; Wu, L.; Zhang, S.; Zhang, Y.; et al. PTK2 Promotes Cancer Stem Cell Traits in Hepatocellular Carcinoma by Activating Wnt/β-Catenin Signaling. Cancer Lett. 2019, 450, 132–143. [Google Scholar] [CrossRef]
- Luo, F.; Li, J.B.; Liu, J.H.; Liu, K.P. Stabilizing and Upregulating Axin with Tankyrase Inhibitor Reverses 5-Fluorouracil Chemoresistance and Proliferation by Targeting the WNT/Caveolin-1 Axis in Colorectal Cancer Cells. Cancer Gene Ther. 2022, 29, 1707–1719. [Google Scholar] [CrossRef]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 Signalling Axis in Cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, L.; Lai, W.; Zeng, Y.; Xu, H.; Lan, Q.; Su, P.; Chu, Z. Interaction with Tumor-associated Macrophages Promotes PRL-3-induced Invasion of Colorectal Cancer Cells via MAPK Pathway-induced EMT and NF-κB Signaling-induced Angiogenesis. Oncol. Rep. 2019, 41, 2790–2802. [Google Scholar] [CrossRef]
- Schiavoni, G.; Gabriele, L.; Mattei, F. The Tumor Microenvironment: A Pitch for Multiple Players. Front. Oncol. 2013, 3, 90. [Google Scholar] [CrossRef] [Green Version]
- Buhrmann, C.; Kraehe, P.; Lueders, C.; Shayan, P.; Goel, A.; Shakibaei, M. Curcumin Suppresses Crosstalk between Colon Cancer Stem Cells and Stromal Fibroblasts in the Tumor Microenvironment: Potential Role of EMT. PLoS ONE 2014, 9, e107514. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Feng, F.; Ma, Y. Tumor-Associated Macrophages as Treatment Target in Colorectal Cancer. Front. Med. Sci. Res. 2022, 4, 12–18. [Google Scholar] [CrossRef]
- Pikarsky, E.; Porat, R.M.; Stein, I.; Abramovitch, R.; Amit, S.; Kasem, S.; Gutkovich-Pyest, E.; Urieli-Shoval, S.; Galun, E.; Ben-Neriah, Y. NF-ΚB Functions as a Tumour Promoter in Inflammation-Associated Cancer. Nature 2004, 431, 461–466. [Google Scholar] [CrossRef]
- Wang, B.-D.; Kline, C.L.B.; Pastor, D.M.; Olson, T.L.; Frank, B.; Luu, T.; Sharma, A.K.; Robertson, G.; Weirauch, M.T.; Patierno, S.R.; et al. Prostate Apoptosis Response Protein 4 Sensitizes Human Colon Cancer Cells to Chemotherapeutic 5-FU through Mediation of an NFκB and MicroRNA Network. Mol. Cancer 2010, 9, 98. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Xu, Y.; Yang, Y.; Liu, Y.; Ma, L.L.; Zhang, Y.Y. Aspirin Suppresses Chemoresistance and Enhances Antitumor Activity of 5-Fu in 5-Fu-Resistant Colorectal Cancer by Abolishing 5-Fu-Induced NF-κB Activation. Sci. Rep. 2019, 9, 16937. [Google Scholar] [CrossRef] [Green Version]
- Shakibaei, M.; Kraehe, P.; Popper, B.; Shayan, P.; Goel, A. Curcumin Potentiates Antitumor Activity of 5-Fluorouracil in a 3D Alginate Tumor Microenvironment of Colorectal Cancer. BMC Cancer 2015, 15, 250. [Google Scholar] [CrossRef] [Green Version]
- Ji, M.; Li, W.; He, G.; Zhu, D.; Lv, S.; Tang, W.; Jian, M.; Zheng, P.; Yang, L.; Qi, Z.; et al. Zinc-A2-Glycoprotein 1 Promotes EMT in Colorectal Cancer by Filamin A Mediated Focal Adhesion Pathway. J. Cancer 2019, 10, 5557–5566. [Google Scholar] [CrossRef]
- Wei, C.; Yang, C.; Wang, S.; Shi, D.; Zhang, C.; Lin, X.; Xiong, B. M2 Macrophages Confer Resistance to 5-Fluorouracil in Colorectal Cancer through the Activation of CCL22/PI3K/AKT Signaling. OncoTargets Ther. 2019, 12, 3051–3063. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Zhang, R.Z.; Zhang, X.J.; Zhang, B.; Yao, Q.H. Curcumin May Reverse 5-Fluorouracil Resistance on Colonic Cancer Cells by Regulating TET1-NKD-Wnt Signal Pathway to Inhibit the EMT Progress. Biomed. Pharmacother. 2020, 129, 110381. [Google Scholar] [CrossRef]
- Koga, N.; Hu, Q.; Sakai, A.; Takada, K.; Nakanishi, R.; Hisamatsu, Y.; Ando, K.; Kimura, Y.; Oki, E.; Oda, Y.; et al. Clinical Significance of Signal Regulatory Protein Alpha (SIRPα) Expression in Esophageal Squamous Cell Carcinoma. Cancer Sci. 2021, 112, 3018–3028. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Mouse) | Sequence (5′ → 3′) |
|---|---|
| CD133 | Forward: CTGCGATAGCATCAGACCAAGC |
| Reverse: CTTTTGACGAGGCTCTCCAGATC | |
| CD44 | Forward: CGGAACCACAGCCTCCTTTCAA |
| Reverse: TGCCATCCGTTCTGAAACCACG | |
| SOX2 | Forward: AACGGCAGCTACAGCATGATGC |
| Reverse: CGAGCTGGTCATGGAGTTGTAC | |
| CD206 | Forward: GTTCACCTGGAGTGATGGTTCTC |
| Reverse: AGGACATGCCAGGGTCACCTTT | |
| VEGF | Forward: CTGCTGTAACGATGAAGCCCTG |
| Reverse: GCTGTAGGAAGCTCATCTCTCC | |
| MMP2 | Forward: CAAGGATGGACTCCTGGCACAT |
| Reverse: TACTCGCCATCAGCGTTCCCAT | |
| MMP9 | Forward: GCTGACTACGATAAGGACGGCA |
| Reverse: TAGTGGTGCAGGCAGAGTAGGA | |
| IL-6 | Forward: TACCACTTCACAAGTCGGAGGC |
| Reverse: CTGCAAGTGCATCATCGTTGTTC | |
| TNF-α | Forward: GGTGCCTATGTCTCAGCCTCTT |
| Reverse: GCCATAGAACTGATGAGAGGGAG | |
| CD47 | Forward: GGTGGGAAACTACACTTGCGAAG |
| Reverse: CTCCTCGTAAGAACAGGCTGATC | |
| SIRPα | Forward: TCATCTGCGAGGTAGCCCACAT |
| Reverse: ACTGTTGGGTGACCTTCACGGT | |
| GAPDH | Forward: CATCACTGCCACCCAGAAGACTG |
| Reverse: ATGCCAGTGAGCTTCCCGTTCAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Y.; Choi, H.S.; Choi, J.H.; Kim, H.-S.; Jang, Y.-S.; Seo, J.-W. 7S,15R-Dihydroxy-16S,17S-epoxy-docosapentaenoic Acid Overcomes Chemoresistance of 5-Fluorouracil by Suppressing the Infiltration of Tumor-Associated Macrophages and Inhibiting the Activation of Cancer Stem Cells in a Colorectal Cancer Xenograft Model. Mar. Drugs 2023, 21, 80. https://doi.org/10.3390/md21020080
Su Y, Choi HS, Choi JH, Kim H-S, Jang Y-S, Seo J-W. 7S,15R-Dihydroxy-16S,17S-epoxy-docosapentaenoic Acid Overcomes Chemoresistance of 5-Fluorouracil by Suppressing the Infiltration of Tumor-Associated Macrophages and Inhibiting the Activation of Cancer Stem Cells in a Colorectal Cancer Xenograft Model. Marine Drugs. 2023; 21(2):80. https://doi.org/10.3390/md21020080
Chicago/Turabian StyleSu, Yan, Hack Sun Choi, Jong Hyun Choi, Hee-Sik Kim, Yong-Suk Jang, and Jeong-Woo Seo. 2023. "7S,15R-Dihydroxy-16S,17S-epoxy-docosapentaenoic Acid Overcomes Chemoresistance of 5-Fluorouracil by Suppressing the Infiltration of Tumor-Associated Macrophages and Inhibiting the Activation of Cancer Stem Cells in a Colorectal Cancer Xenograft Model" Marine Drugs 21, no. 2: 80. https://doi.org/10.3390/md21020080
APA StyleSu, Y., Choi, H. S., Choi, J. H., Kim, H. -S., Jang, Y. -S., & Seo, J. -W. (2023). 7S,15R-Dihydroxy-16S,17S-epoxy-docosapentaenoic Acid Overcomes Chemoresistance of 5-Fluorouracil by Suppressing the Infiltration of Tumor-Associated Macrophages and Inhibiting the Activation of Cancer Stem Cells in a Colorectal Cancer Xenograft Model. Marine Drugs, 21(2), 80. https://doi.org/10.3390/md21020080