
Structural Characterization and Anti-Inflammatory Effects of 24-Methylcholesta-5(6), 22-Diene-3β-ol from the Cultured Marine Diatom Phaeodactylum tricornutum; Attenuate Inflammatory Signaling Pathways


Abstract
:1. Introduction
2. Results
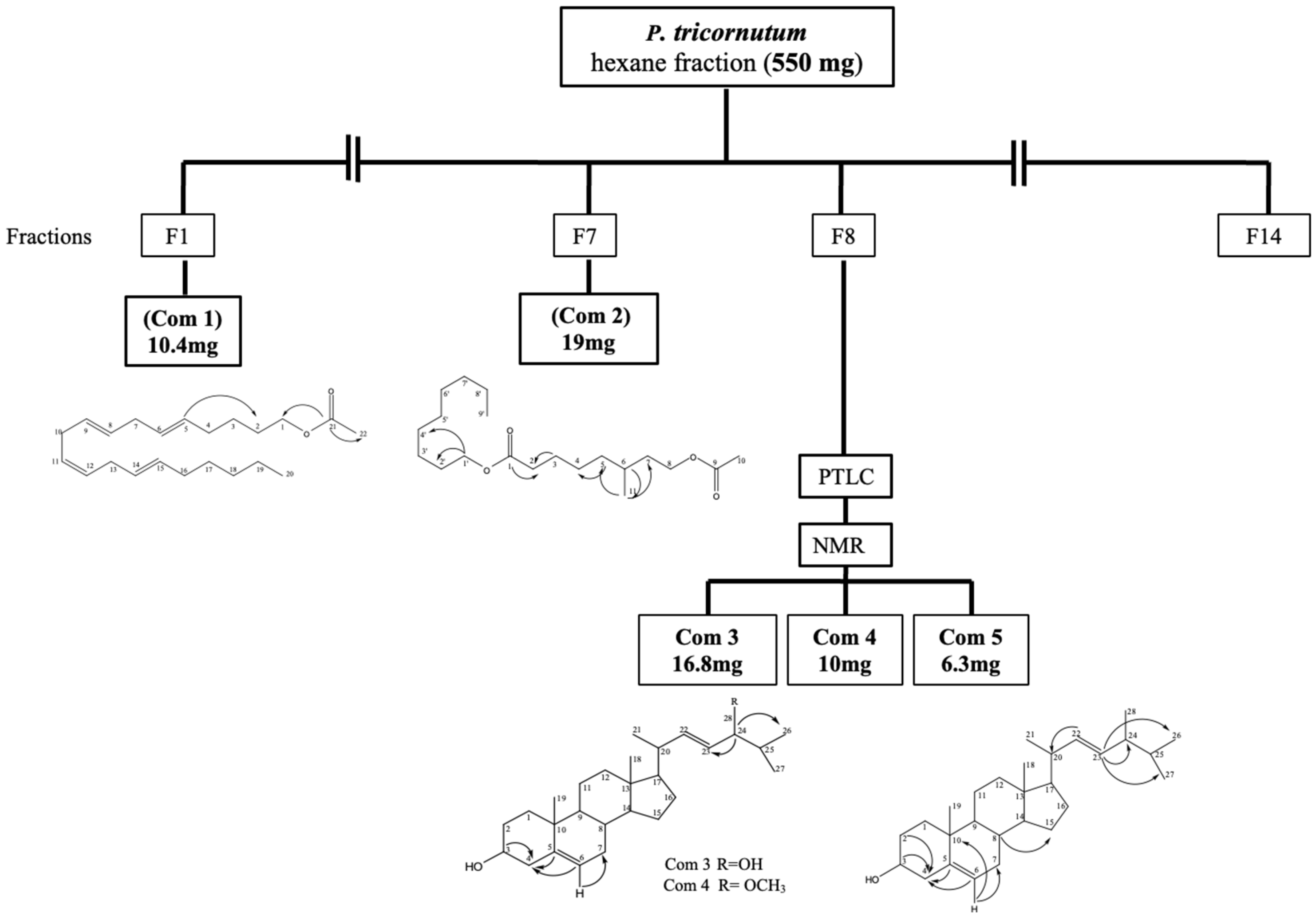
2.1. Structure Elucidation of the Compound 24-Methylcholesta-5(6), 22-Diene-3β-ol from the Cultured Marine Diatom Phaeodactylum tricornutum
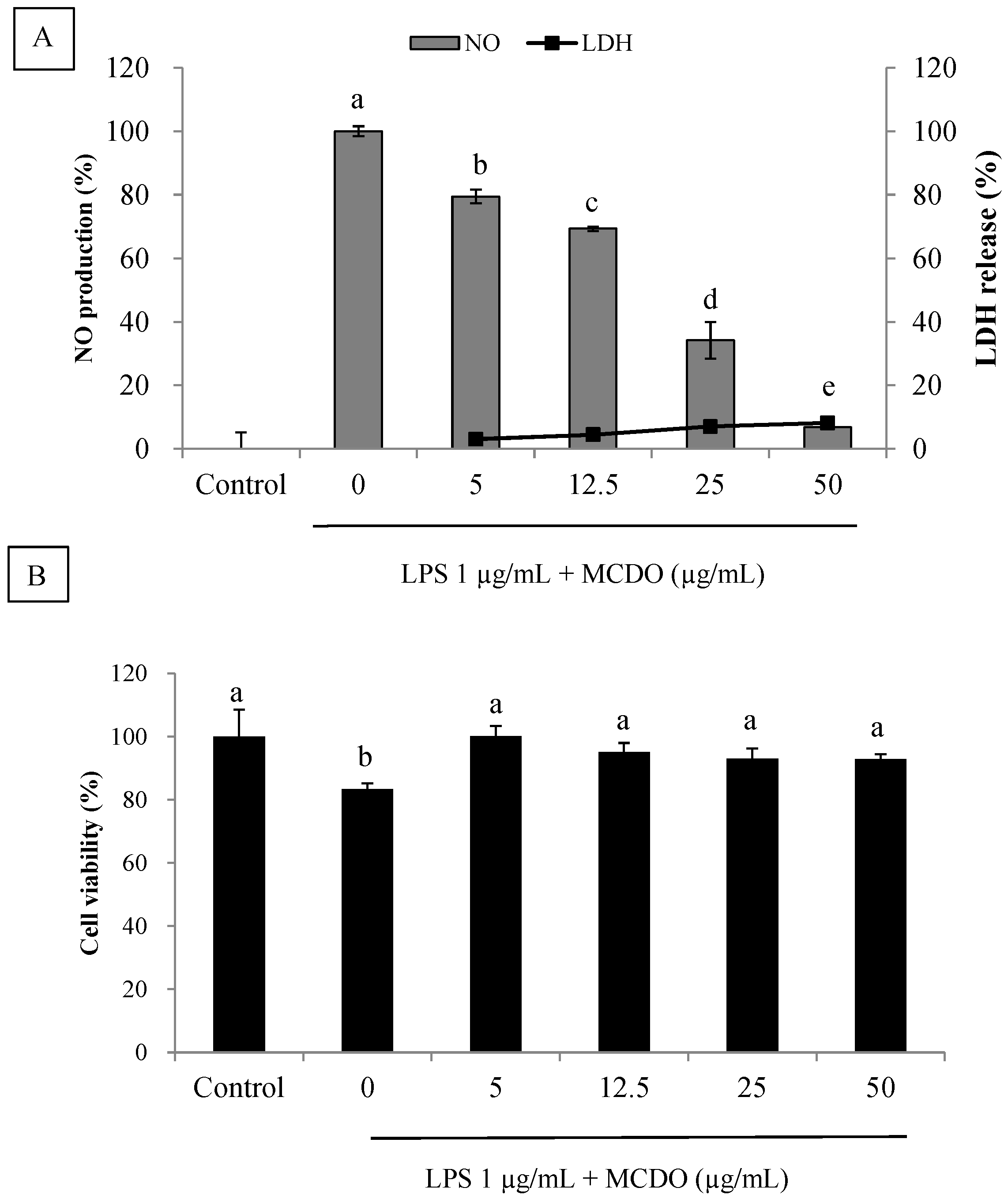
2.2. Inhibitory Effect of MCDO on LPS-Induced Nitric Oxide Production and Cytotoxicity
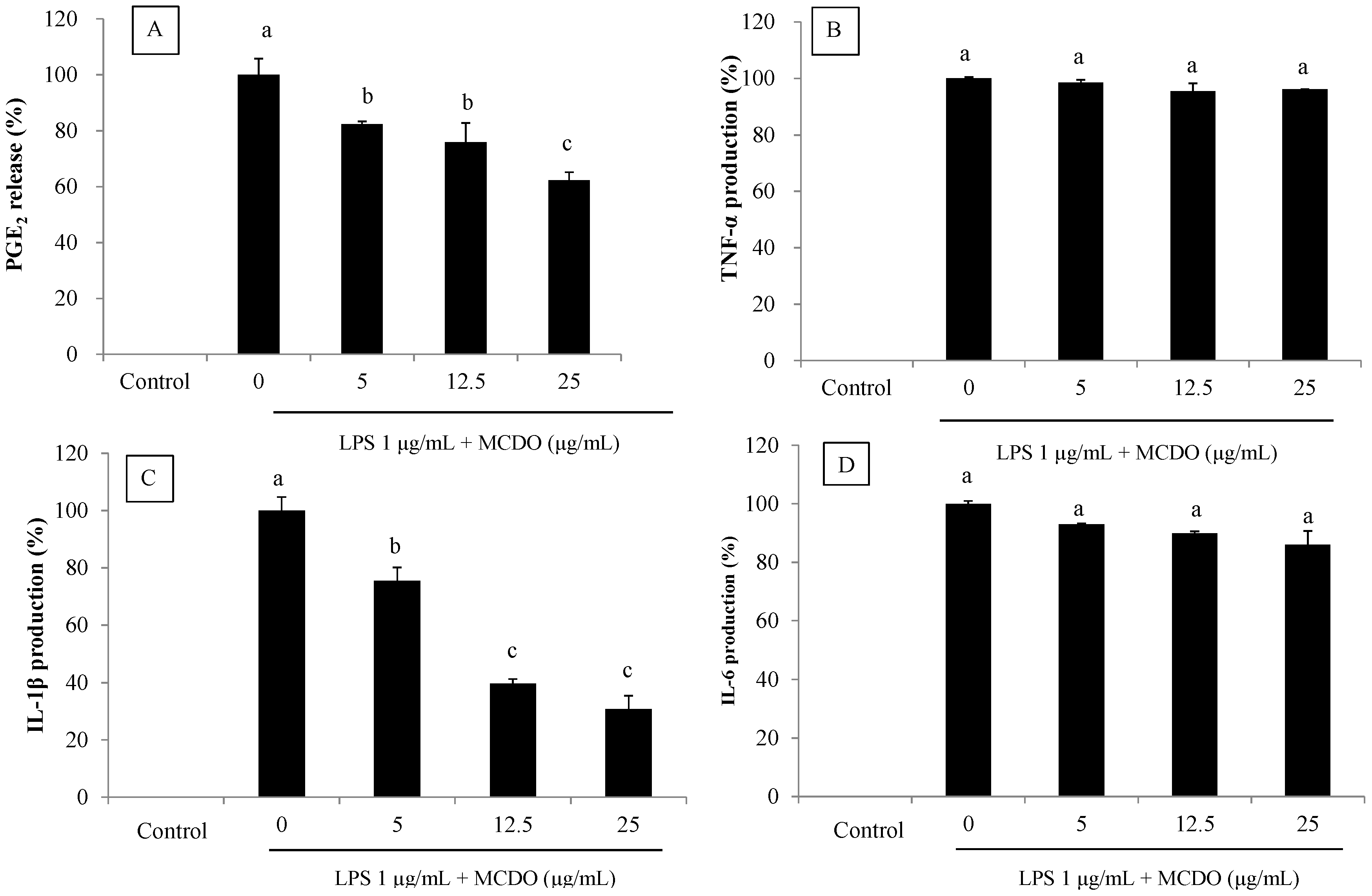
2.3. Inhibitory Effect of MCDO on LPS-Induced PGE2 Production
2.4. Inhibitory Effect of MCDO on LPS-Induced Pro-Inflammatory Cytokine (TNF-α, IL-1β, and IL-6) Production
2.5. Inhibitory Effect of MCDO on LPS-Induced iNOS and COX-2 Protein Expression
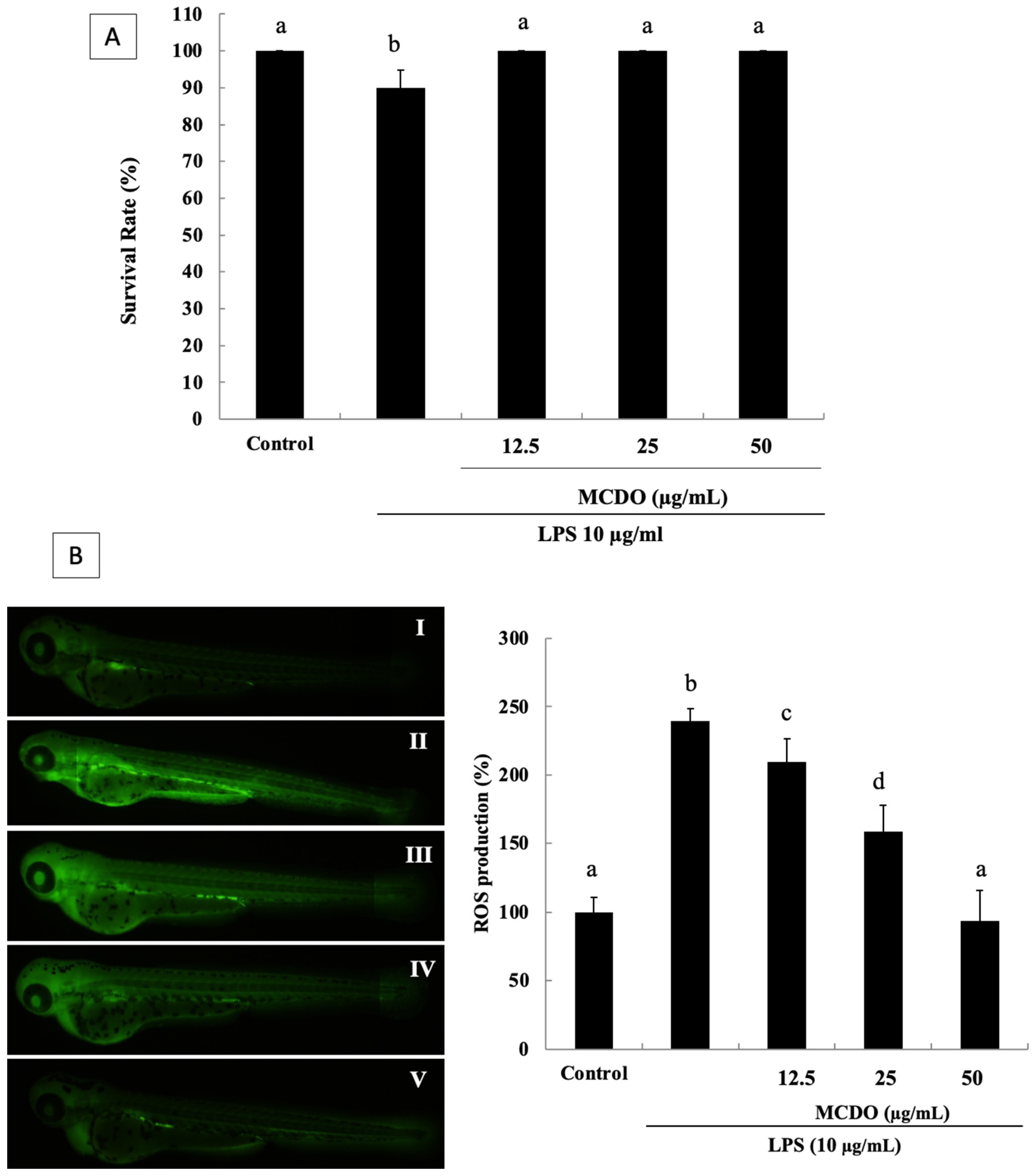
2.6. Evaluation of Toxicity and Survival Rate of MCDO in the Zebrafish Embryo
2.7. Inhibitory Effect of the MCDO on LPS-Induced ROS Generation in Zebrafish
2.8. Inhibitory Effect of MCDO on LPS-Induced Nitric Oxide (NO) Production in Zebrafish
2.9. Protective Effect of MCDO on LPS-Induced Cell Death in Live Zebrafish
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Extraction, Isolation, and Characterization of Compounds
4.3. Cell Culture Method
4.4. Detection of Nitric Oxide (NO) Production
4.5. Lactate Dehydrogenase (LDH) Cytotoxicity Assay
4.6. Detection of Pro-Inflammatory Cytokines, Tumor Necrosis Factor-α (TNF-α) Interleukin-1β (IL-1β), and Interleukin-6 (IL-6) Production
4.7. Detection of Prostaglandin –E2 (PGE2) Production
4.8. Western Blot Analysis
4.9. Origin and Maintenance of Zebrafish
4.10. Waterborne Exposure of Embryos to Samples and LPS
4.11. Estimation of Oxidative Stress-Induced Reactive Oxygen Species (ROS) Generation and Image Analysis
4.12. Estimation of Oxidative Stress-Induced Nitric Oxide (NO) Generation and Image Analysis
4.13. Estimation of Oxidative Stress-Induced Cell Death and Image Analysis
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Serhan, C.N.; Savill, J. Resolution of inflammation: The beginning programs the end. Nat. Immunol. 2005, 6, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Lee, J.M.; Jun, S.H.; Lee, S.H.; Kim, N.W.; Lee, J.H.; Ko, N.Y.; Mun, S.H.; Kim, B.K.; Lim, B.O.; et al. The anti-inflammatory effects of Pyrolae herba extract through the inhibition of the expression of inducible nitric oxide synthase (iNOS) and NO production. J. Ethnopharmacol. 2007, 112, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Wichard, T.; Pohnert, G.J. Formation of halogenated medium chain hydrocarobons by a lipoxygenase/hydroperoxide haloyase-mediated transformation in plantonic microalgae. J. Am. Chem. Soc. 2006, 128, 7114–7115. [Google Scholar] [CrossRef]
- Veron, B.; Billard, C.; Dauguel, J.C.; Hartmann, M.A. Sterol composition of Phaeodactylim tricornutum as influenced by growth temperature and light Spectral quality. Lipids 1996, 31, 989–994. [Google Scholar] [CrossRef] [PubMed]
- Fabregas, J.; Aran, J.; Morales, E.D.; Lamela, T.; Otero, A. Modification of Sterol concentration in marine microalga. Phytochemistry 1997, 46, 1189–1191. [Google Scholar] [CrossRef]
- Sahu, A.; Panch, I.; Jain, D.; Paliwaql, C.; Ghosh, T.; Patidar, S.; Bhattachaya, S.; Mishra, S. Fatty acids as biomarker of Microalgae. Phytochemistry 2013, 89, 53–58. [Google Scholar] [CrossRef]
- Lv, X.; Zou, L.I.; Sun, B.; Wang, J.; Sun, M.Y. Variation in lipid yield and compositions of marine microalgae during cell growth and respiration, and within intracellular structure. J. Exp. Mar. Biol. Ecol. 2010, 391, 73–83. [Google Scholar] [CrossRef]
- Prestegard, S.K.; Erga, S.R.; Steinrucken, P.; Mjos, S.A.; Knutsen, G.; Rohloff, J. Specific metabolites in a Phaeodactylim tricornutum strain isolated from western Norwegian Fjord Water. Mar. Drugs 2015, 14, 9. [Google Scholar] [CrossRef]
- Libralato, G.; Gentile, E.; Ghirardini, A.V. Wastewater effects on Phaeodactylim tricornutum (Bohlin): Setting up a classification system. Ecol. Indic. 2016, 60, 31–37. [Google Scholar] [CrossRef]
- Taipale, S.J.; Hiltunen, M.; Vuorio, K.; Peltomaa, E. Suitability of Phytosterol alongside fatty acids as chemotaxonomic biomarkers for phytoplankton. Front. Plant Sci. 2016, 7, 212. [Google Scholar] [CrossRef]
- Rapso, M.F.J.; Morais, R.M.S.C.; Morias, A.M.M.B. Bioactivity and application of sulphated polysaccharides from marine microalgae. Mar. Drugs 2013, 11, 233–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desbois, A.P.; Meams-Spragg, A.; Smith, V.J. A fatty acid from the diatom Phaeodactylum tricornutum is antibacterial against diverse bacteria including multi-resistant Staphylococcus aureus (MRSA). Mar. Biotechnol. 2009, 11, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Flori, S.; Jouneau, P.H.; Finazzi, G.; Marechal, E.; Falconel, D. Ultrastructure of the periplastidial compartment of the diatom Phaeodactylim tricornutum. Protist 2016, 167, 254–267. [Google Scholar] [CrossRef]
- Carreto, J.I.; Catoggio, J.A. Variations in pigment contents of the diatom Phaeodactylim tricornutum during growth. Mar. Biol. 1976, 36, 105–112. [Google Scholar] [CrossRef]
- Zhukova, N.V.; Aizdaicher, N.A. Fatty acid composition of 15 species of marine microalgae. Phytochemistry 1995, 39, 351–356. [Google Scholar] [CrossRef]
- Fajardo, A.F.; Cerdan, L.E.; Medina, A.R.; Fernandaz, F.G.A.; Moreno, P.A.G.; Grima, E.M. Lipid extraction from the microalga Phaeodactylim tricornutum. Eur. J. Lipid Sci. Technol. 2007, 109, 120–126. [Google Scholar] [CrossRef]
- Rapso, M.F.J.; Morais, R.M.S.C.; Morais, A.M.M.M.B. Health applications of bioactive compounds from marine microalgae. Life Sci. 2013, 93, 479–486. [Google Scholar] [CrossRef]
- Guzman, S.; Gato, A.; Lamela, M.; Garabal, M.F.; Calleja, J.M. Anti-inflammatory and immunomodulattory activities of polysaccharide form Chlorella stigmatophora and Phaeodactylim tricornutum. Phytother. Res. 2003, 17, 665–670. [Google Scholar] [CrossRef]
- Becker, W. Microalgae in human and animal nutrition. In Handbook of Microalgal Culture; Richmond, A., Ed.; Blackwell Publishing: Oxford, UK, 2004; pp. 312–351. [Google Scholar]
- Kawaguchi, S.; Maita, Y. Amino acid sequence of cadmium-binding peptide induced in a marine diatom, Phaeodactylim tricornutum. Bull. Environ. Contam. Toxicol. 1990, 45, 893–899. [Google Scholar] [CrossRef]
- Morelli, E.; Cruz, B.H.; Somovigo, S.; Scarano, G. Speciation of cadmium-γ-glutamyl peptides complexes in cells of the marine microalga Phaeodactylim tricornutum. Plant Sci. 2002, 163, 807–813. [Google Scholar] [CrossRef]
- Guzmán-Murillo, M.A.; López-Bolaños, C.C.; Ledesma-Verdejo, T.; Roldan-Libenson, G.; Cadena-Roa, M.A.; Ascencio, F. Effects of fertilizer-based culture media on the production of exocellular polysaccharides and cellular superoxide dismutase by Phaeodactylum tricornutum (Bohlin). J. Appl. Phycol. 2007, 19, 33–40. [Google Scholar] [CrossRef]
- Jimeno, J.; Faircloth, G.; Sousa-Fero, J.M.F.; Scheuer, P.; Rinhart, K. New marine-derived anticancer therapeutics—A journey from the sea to clinical trials. Mar. Drugs 2004, 2, 14–29. [Google Scholar] [CrossRef] [Green Version]
- Abd El Baky, H.H.; El-Baroty, G.S. Healthy benefit of microalgal bioactive substances. J. Aquat. Sci. 2013, 1, 11–23. [Google Scholar]
- Rubinstien, I.; Goad, L.J. Occurrence of (24S)-24-methylcholesta-5(6), 22-diene-3β-ol in the diatom Phaeodactylim tricornutum. Phytochemistry 1974, 13, 485–487. [Google Scholar] [CrossRef]
- Goad, L.J.; Withers, N. Identification of 27-nor-(24R)-24-methylcholesta-5,22-dien-3β-ol and brassicasterol as the major sterols of the marine dinoflagellate Gymnodinium simplex. Lipids 1982, 17, 853–858. [Google Scholar] [CrossRef]
- Echigo, S.; Castellanos, L.; Duque, C.; Uekusa, H.; Hara, N. Fujimoto. C-24 Stereochemistry of marine Sterols: (24 E)-24-Ethyl-24-methylcholesta-5(6), 22-diene-3β-ol and 24-Ethyl- 24-methylcholest-5-en-3β-ol. J. Braz. Chem. Soc. 2011, 22, 997–1004. [Google Scholar] [CrossRef] [Green Version]
- Radakovits, R.; Eduafo, P.M.; Posewitz, M.C. Genetic engineering of fatty acid chain length in Phaeodactylim tricornutum. Metab. Eng. 2011, 13, 89–95. [Google Scholar] [CrossRef]
- Hong, J.W.; Kim, S.A.; Chang, J.; Yi, J.; Jeong, J.; Kim, S.; Kim, S.H.; Yoon, S.H. Isolation and description of a Korean microalga, Asterarcys quadricellulare KNUA020, and analysis of its biotechnological potential. Algae 2012, 27, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Plaza, M.; Herrero, M.; Cifuentes, A.; Ibanez, E. Innovative Natural Functional Ingredients from Microalgae. J. Agric. Food Chem. 2009, 57, 7159–7170. [Google Scholar] [CrossRef]
- Samarakoon, K.; Jeon, Y.J. Bio-functionalities of proteins derived from marine algae—A review. Food Res. Int. 2012, 48, 948–960. [Google Scholar] [CrossRef]
- Garcia, C.M.C.; Camacho, F.G.; Miron, A.S.; Sevilla, J.M.F.; Chisti, Y.; Grima, E.M. Mixotrophic production of marine microalgae Phaeodactylim tricornutum on various carbon sources. J. Microbiol. Biotechnol. 2006, 16, 689–694. [Google Scholar]
- Aldini, R.; Micucci, M.; Cevenini, M.; Fato, R.; Bergamini, C.; Nanni, C.; Cont, M.; Camborata, C.; Spinozzi, S.; Montagnani, M.; et al. Antiinflammatory Effect of Phytosterols in Experimental Murine Colitis Model: Prevention, Induction, Remission Study. PLoS ONE 2014, 9, e108112. [Google Scholar] [CrossRef] [PubMed]
- Samarakoon, K.W.; Ko, J.Y.; Lee, J.H.; Shah, M.M.R.; Kwon, O.N.; Kang, M.C.; Lee, J.B.; Jeon, Y.J. In vitro studies of anti-inflammatory and anticancer activities of organic solvent extracts from cultured marine microalgae. Algae 2013, 28, 111–119. [Google Scholar] [CrossRef]
- Samarakoon, K.W.; Ko, J.Y.; Lee, J.H.; Kwon, O.N.; Kim, S.W.; Jeon, Y.J. Apoptotic anticancer activity of novel fatty alchohol ester isolated from cultured marine diatom, Phaeodactylim tricornutum. J. Funct. Foods 2014, 6, 231–240. [Google Scholar] [CrossRef]
- Vanmierlo, T.; Popp, J.; Kölsch, H.; Friedrichs, S.; Jessen, F.; Stoffel-Wagner, B.; Bertsch, T.; Hartmann, T.; Maier, W.; von Bergmann, K.; et al. The plant sterol brassicasterol as an additional CSF biomarker in Alzheimer’s disease. Acta Psychiatr. Scand. 2011, 124, 184–192. [Google Scholar] [CrossRef]
- Bianconi, V.; Mannarino, M.R.; Sahebkar, A.; Cosentino, T.; Pirro, M. Cholesterol-Lowering Nutraceuticals Affecting Vascular Function and Cardiovascular Disease Risk. Curr. Cardiol. Rep. 2018, 20, 53. [Google Scholar] [CrossRef]
- Hassan, S.T.S. Brassicasterol with Dual Anti-Infective Properties against HSV-1 and Mycobacterium tuberculosis, and Cardiovascular Protective Effect: Nonclinical In Vitro and In Silico Assessments. Biomedicines 2020, 8, 132. [Google Scholar] [CrossRef]
- Wang, J.W.; Zheng, L.P.; Wu, J.Y.; Tan, R.X. Involvement of nitric oxide in oxidative burst phenylalanine ammonia-lyase activation and Taxol production induced by low-energy ultrasound in Taxus yunnanensis cell suspension cultures. Nitric Oxide 2006, 15, 351–358. [Google Scholar] [CrossRef]
- Kassim, M.; Achoui, M.; Mustafa, M.R.; Mohd, M.A.; Yusoff, K.M. Ellagic acid, phenolic acids, and flavonoids in Malaysian honey extracts demonstrate in vitro anti-inflammatory activity. Nutr. Res. 2010, 30, 650–659. [Google Scholar] [CrossRef]
- Kim, H.K.; Cheon, B.S.; Kim, Y.H.; Kim, S.Y.; Kim, H.P. Effects of naturally occurring flavonoids on nitric oxide production in the macrophage cell line RAW 264.7 and their structure-activity relationship. Biochem. Pharmacol. 1999, 58, 759–765. [Google Scholar] [CrossRef]
- Suh, G.H.; Youn, Y.K.; Song, H.G.; Rhee, J.E.; Jung, S.E. The effect of glutamine on inducible nitric oxide synthase gene expression in intestinal ischemia-reperfusion. Nutr. Res. 2003, 23, 131–140. [Google Scholar]
- Nakagawa, T. Roles of prostaglandin E2 in a cochlea. Hear Res. 2011, 276, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Li, P.C.; Chen, B.C.; Chang, M.S.; Wang, J.L.; Chiu, W.T.; Lin, C.H. Lipoteichoic acid-induced nitric oxide synthase expression in RAW 264.7 macrophages is mediated by cyclooxygenase-2, prostaglandin E2, protein kinase A, p38 MAPK, and nuclear factor-kappa B pathways. Cell Signal. 2006, 18, 1235–1243. [Google Scholar] [CrossRef]
- Ahmed, N.; Chen, L.C.; Gordon, M.A.; Laskin, J.D.; Laskin, D.L. Regulation of cyclooxygenase-2 by nitric oxide in activated hepatic macrophages during acute endotoxemia. J. Leukoc. Biol. 2002, 71, 1005–1011. [Google Scholar] [CrossRef]
- Park, Y.G.; Kang, S.K.; Kim, W.J.; Lee, Y.C.; Kim, C.H. Effect of TGF-β, IL-βand IL-6 alone or in combination, and tyrosine kinase inhibitor on cyclooxygenase expression prostaglandin E2 production and bone resorption in mouse calvarial bone cells. Int. J. Biochem. Cell Biol. 2004, 36, 2270–2280. [Google Scholar] [CrossRef]
- Eisen, J.S. Zebrafish make a big splash. Cell 1996, 87, 969–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanjeewa, K.A.; Jayawardena, T.U.; Kim, S.Y.; Kim, H.S.; Ahn, G.; Kim, J.I.; Jeon, Y.J. Fucoidan isolated from invasive Sargassum horneri inhibit LPS-induced inflammation via blocking NF-κB and MAPK pathways. Algal Res. 2019, 41, 101561. [Google Scholar] [CrossRef]
- Novoa, B.; Bowman, T.V.; Zon, L.; Figueras, A. LPS response and tolerance in the Zebrafish (Danio rerio). Fish Shelfish Immunol. 2009, 26, 326–331. [Google Scholar] [CrossRef] [Green Version]
- Pichler, F.B.; Laurenson, S.; Williams, L.C.; Dodd, A.; Copp, B.R.; Love, D.R. Chemical discovery and global gene expression analysis in zebrafish. Nat. Biotechnol. 2003, 21, 879–883. [Google Scholar] [CrossRef]
- Liao, Y.-F.; Chiou, M.-C.; Tsai, J.-N.; Wen, C.-C.; Wang, Y.-H.; Cheng, C.-C.; Chen, Y.-H. Resveratrol treatment attenuates the wound-induced inflammation in zebrafish larvae through the suppression of myeloperoxidase expression. J. Food Drug Anal. 2011, 19, 167–173. [Google Scholar] [CrossRef]
- Park, K.H.; Cho, K.H. A zebrafish model for the rapid evaluation of prooxidative and inflammatory death by lipopolysaccharide, oxidized low-density lipoproteins and glycated high-density lipoproteins. Fish Shellfish Immunol. 2011, 31, 904–910. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Hyun, E.A.; Yoon, W.J.; Kim, B.H.; Rhee, M.H.; Kang, H.K.; Cho, J.Y.; Yoo, E.S. In vitro anti-inflammatory and anti-oxidative effects of Cinnamomum camphora extracts. J. Ethnopharmacol. 2006, 103, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Baik, K.U.; Jung, J.H.; Park, M.H. In vitro anti-inflammatory effects of cynaropicrin, a sesquiterpene lactone, from Saussurea lappa. Eur. J. Pharmacol. 2000, 398, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, A.R.; Schmaldienst, S.; Stuhlmeier, K.M.; Chen, W.; Knapp, W.; Zlabinger, G.J. A microplate assay for the detection of oxidative products using 2,7-dichlorofluorescein-diacetate. J. Immunol. Methods 1992, 156, 39–45. [Google Scholar] [CrossRef]
- Itoh, Y.; Ma, F.H.; Hoshi, H.; Oka, M.; Noda, K.; Ukai, Y.; Kojima, H.; Nagano, T.; Toda, N. Determination and bioimaging method for nitric oxide in biological specimens by diaminofluorescein fluorometry. Anal. Biochem. 2000, 287, 203–209. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Molecular Formula | IUPAC Name | Abbreviations |
|---|---|---|---|
| Com 1 | C22H36O2 | Icosa-5, 8, 11, 14-tetraenyl acetate | ITEA |
| Com 2 | C20H38O4 | Nonyl 8-acetoxy-6-methyloctanoate | NAMO |
| Com 3 | C27H44O2 | Cholestra-5(6), 22-diene-3, 24β-diol | CDDO |
| Com 4 | C28H46O2 | Cholestra-5(6), 22-dien-3, 24β-diol, methyl ether | CDDME |
| Com 5 | C28H46O | 24-methylcholesta-5(6), 22-diene-3β-ol | MCDO |
| Position (C#) | δC a (ppm) | (mult) | δH a (mult, JHH Hz) |
|---|---|---|---|
 | |||
| 1 | 37.32 | (CH2) | 1.85/1.02 m |
| 2 | 31.73 | (CH2) | 1.50/1.42 m |
| 3 | 71.9 | (CH) | 3.5 m |
| 4 | 42.38 | (CH2) | 2.27/2.19 m |
| 5 | 140.73 | (C) | |
| 6 | 121.71 | (CH) | 5.32 (1 H br d, J = 4.8 Hz) |
| 7 | 31.98 | (CH2) | 1.98/1.85 m |
| 8 | 43.15 | (CH) | 2.24 m |
| 9 | 51.38 | (CH) | 0.89 m |
| 10 | 36.5 | (C) | |
| 11 | 21.15 | (CH2) | 1.48/1.42 m |
| 12 | 39.75 | (CH2) | 1.95/2.05 m |
| 13 | 42.47 | (C) | |
| 14 | 56.85 | (CH) | 1.16 m |
| 15 | 24.41 | (CH2) | 1.50/1.54 m |
| 16 | 28.91 | (CH2) | 1.24/1.64 m |
| 17 | 55.93 | (CH) | 1.14 m |
| 18 | 12.13 | (CH3) | 0.68 s |
| 19 | 19.49 | (CH3) | 0.99 s |
| 20 | 40.37 | (CH) | 1.98 m |
| 21 | 21.07 | (CH3) | 0.88 (3H d, J = 6.9 Hz) |
| 22 | 136.63 | (CH) | 5.15 (1 H, dd, J = 8.4, J = 15.3 Hz) |
| 23 | 131.63 | (CH) | 5.12 (1 H, dd, J = 8.4, J = 15.3 Hz) |
| 24 | 50.13 | (CH) | 0.89 m |
| 25 | 33.3 | (CH) | 1.48 m |
| 26 | 20.24 | (CH3) | 0.89 (3 H, d, J = 6.3 Hz) |
| 27 | 19.7 | (CH3) | 0.81 (3 H, d, J = 6.9 Hz) |
| 28 | 18.78 | (CH3) | 0.99 (3 H, d, J = 7.0 Hz) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samarakoon, K.W.; Kuruppu, A.I.; Ko, J.-Y.; Lee, J.-H.; Jeon, Y.-J. Structural Characterization and Anti-Inflammatory Effects of 24-Methylcholesta-5(6), 22-Diene-3β-ol from the Cultured Marine Diatom Phaeodactylum tricornutum; Attenuate Inflammatory Signaling Pathways. Mar. Drugs 2023, 21, 231. https://doi.org/10.3390/md21040231
Samarakoon KW, Kuruppu AI, Ko J-Y, Lee J-H, Jeon Y-J. Structural Characterization and Anti-Inflammatory Effects of 24-Methylcholesta-5(6), 22-Diene-3β-ol from the Cultured Marine Diatom Phaeodactylum tricornutum; Attenuate Inflammatory Signaling Pathways. Marine Drugs. 2023; 21(4):231. https://doi.org/10.3390/md21040231
Chicago/Turabian StyleSamarakoon, Kalpa W., Anchala I. Kuruppu, Ju-Young Ko, Ji-Hyeok Lee, and You-Jin Jeon. 2023. "Structural Characterization and Anti-Inflammatory Effects of 24-Methylcholesta-5(6), 22-Diene-3β-ol from the Cultured Marine Diatom Phaeodactylum tricornutum; Attenuate Inflammatory Signaling Pathways" Marine Drugs 21, no. 4: 231. https://doi.org/10.3390/md21040231
APA StyleSamarakoon, K. W., Kuruppu, A. I., Ko, J. -Y., Lee, J. -H., & Jeon, Y. -J. (2023). Structural Characterization and Anti-Inflammatory Effects of 24-Methylcholesta-5(6), 22-Diene-3β-ol from the Cultured Marine Diatom Phaeodactylum tricornutum; Attenuate Inflammatory Signaling Pathways. Marine Drugs, 21(4), 231. https://doi.org/10.3390/md21040231