
Advances in Natural Products from the Marine-Sponge-Associated Microorganisms with Antimicrobial Activity in the Last Decade

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Metabolite Sources, Structures and Activities
2.1. Marine Natural Products with Antimicrobial Activity from Sponge-Associated Fungi
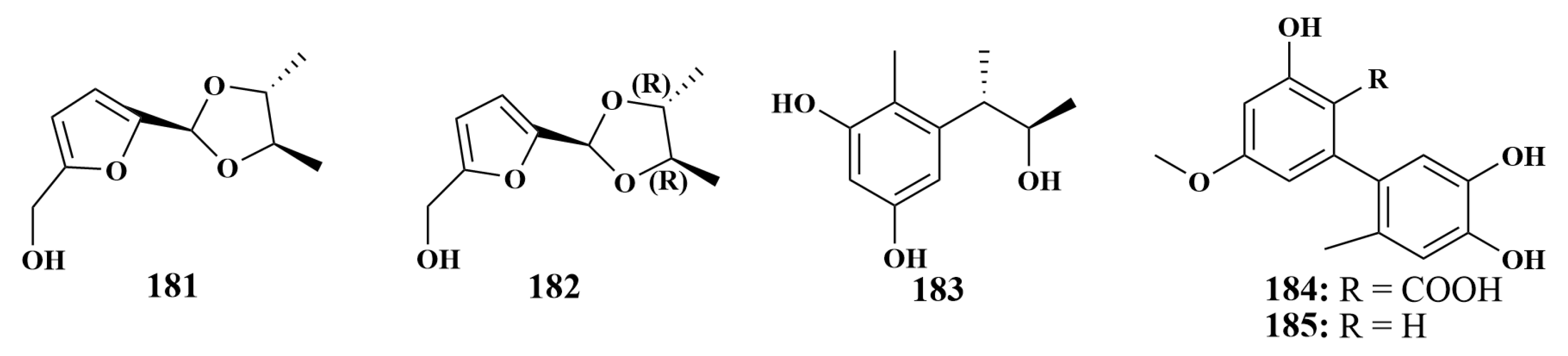
2.1.1. Terpenoids
2.1.2. Polyketides
2.1.3. Alkaloids
2.1.4. Peptides
2.1.5. Others
2.2. Marine Natural Products with Antimicrobial Activity from Sponge-Associated Actinomycetes
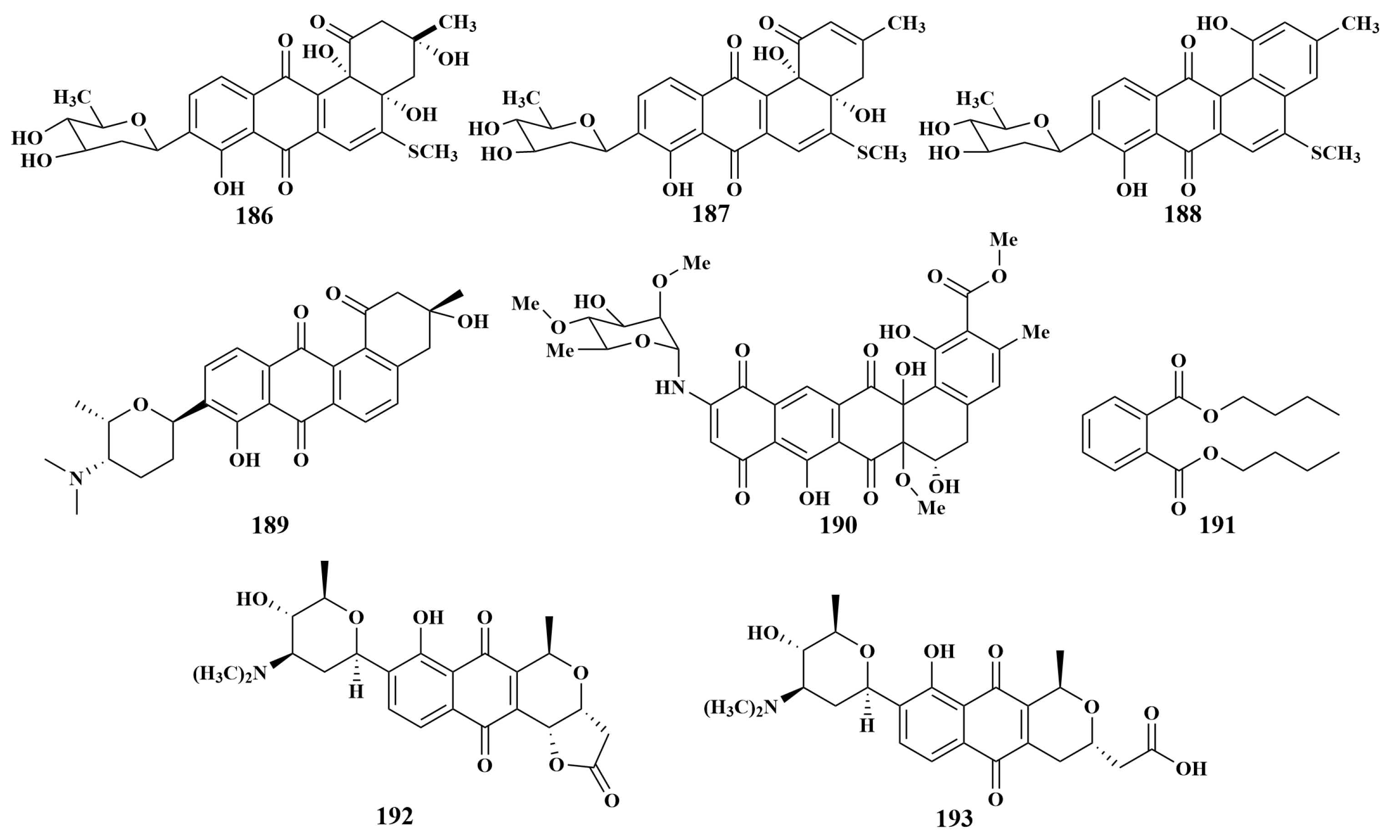
2.2.1. Polyketides
2.2.2. Alkaloids
2.2.3. Peptides
2.2.4. Other Nitrogen-Containing Metabolites
2.2.5. Glucosides
2.3. Marine Natural Products with Antimicrobial Activity from Sponge-Associated Bacterium
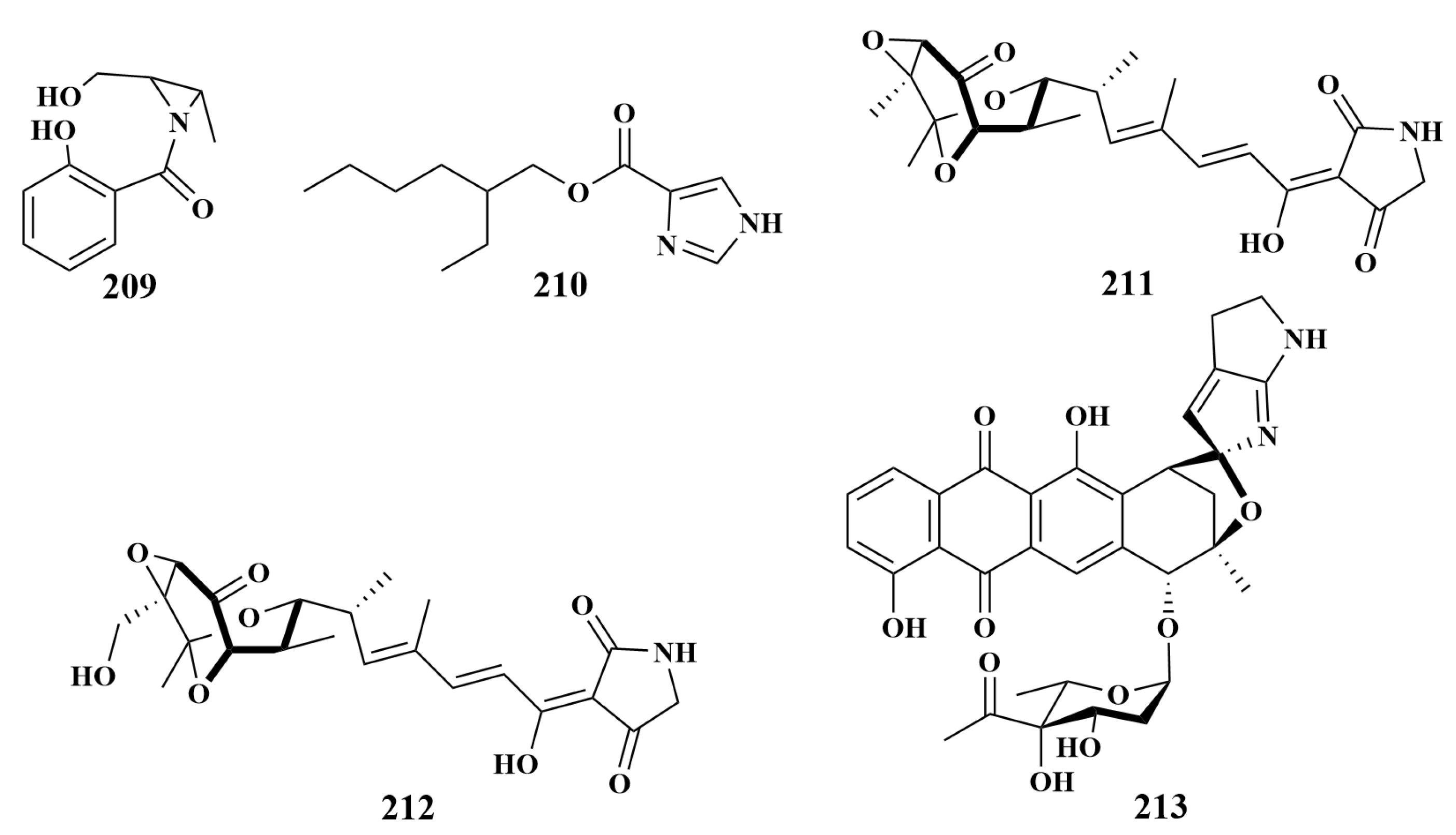
2.3.1. Polyketides
2.3.2. Alkaloids
2.3.3. Peptides
2.4. Marine Natural Products with Antimicrobial Activity from Sponge-Associated Strains through Co-Cultivation Method
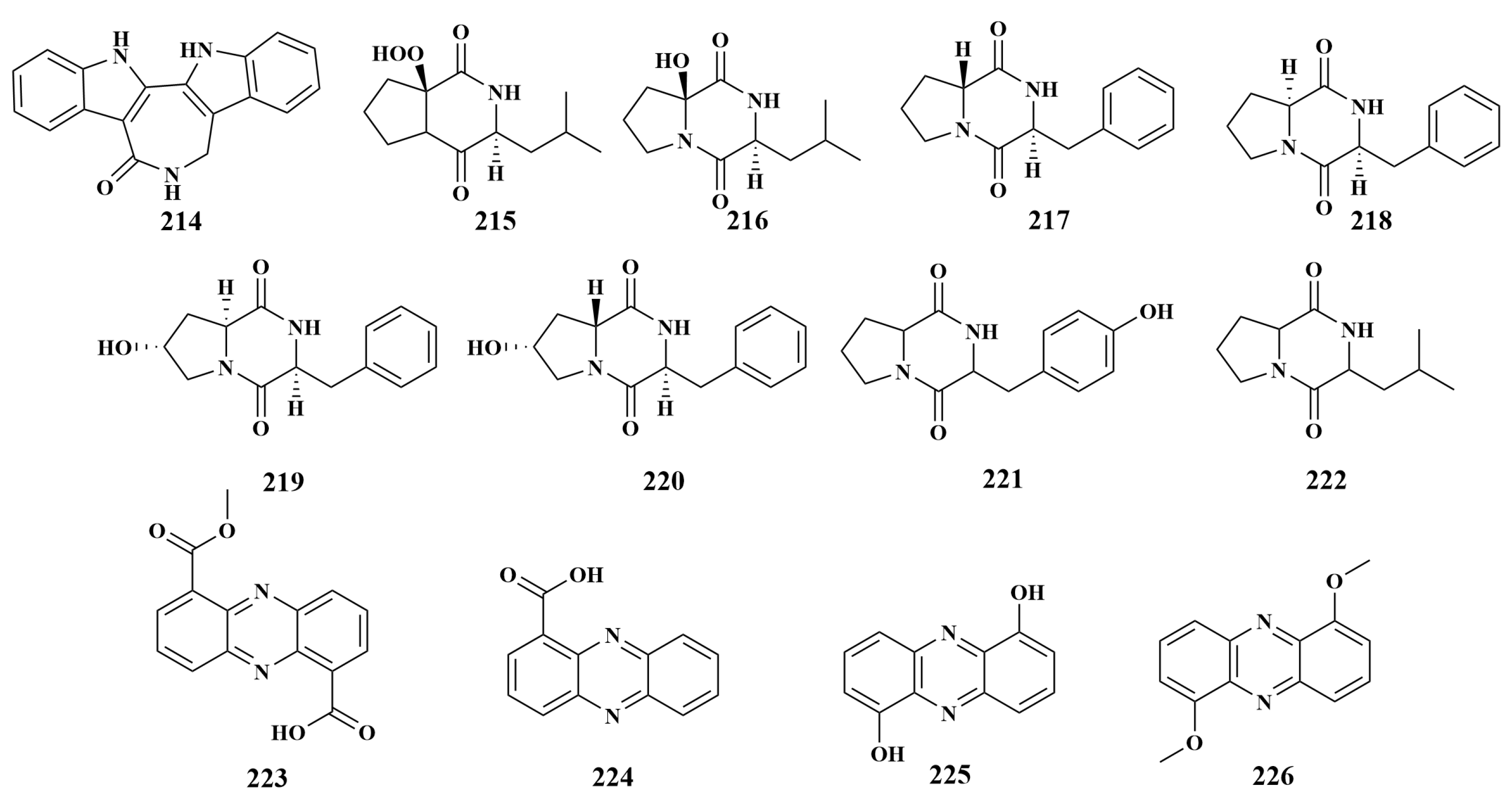
2.4.1. Polyketides
2.4.2. Alkaloids
2.4.3. Peptides
2.4.4. Other Nitrogen-Containing Metabolites
3. Discussion
4. Conclusions and Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| S. aureus | Staphylococcus aureus | E.faecalis | Enterococcus faecalis |
| E. tarda | Edwardsiella tarda | E.faecium | Enterococcus faecium |
| F. solani | Fusarium solani | E.coli | Escherichia coli |
| B. subtilis | Bacillus subtilis | C.albicans | Candida albicans |
| S. epidermidis | Staphylococcus epidermidis | MRSA | methicillin-resistant Staphylococcus aureus |
| V. parahaemolyticus | Vibrio parahaemolyticus | A.baumannii | Acinetobacter baumannii |
| V. proteolyticus | Vibrio proteolyticus | P.mirabilis | Proteus mirabilis |
| P. fluorescens | Pseudomonas fluorescens | S.flexneri | Shigella flexneri |
| S. cholerasuis | Salmonella cholerasuis | L.monocytogenes | Listeria monocytogenes |
| V. scophthalmi | Vibrio scophthalmi | V.shilonii | Vibrio shilonii |
| V. brasiliensis | Vibrio brasiliensis | VRE | vancomycin-resistant Enterococcus |
| C. neoformans | Cryptoccus neoformans | M.gypseum | Microsporum gypseum |
| C. trachamatis | Chlamydia trachamatis | B.cereus | Bacillus cereus |
| M. luteus | Micrococcus luteus | S.typhimurium | Salmonella typhimurium |
| M. tuberculosis | Mycobacterium tuberculosis | K.pneumoniae | Klebsiella pneumoniae |
| S. pneumoniae | Streptococcus pneumoniae | S.enterica | Salmonella enterica |
| V. harvey | Vibrio harvey | BCG | Bacille Calmette Guerin |
| P. lachrymans | Pseudomonas lachrymans | MSSA | methicillin-sensitive Staphycoccus aureus |
| A. tumefaciens | Agrobacterium tumefaciens | X.vesicatoria | Xanthomonas vesicatoria |
| R. solanacearum | Ralstonia solanacearum | C.parapsilosis | Candida parapsilosis |
| A. fumigatus | Aspergillus fumigatus | A.niger | Aspergillus niger |
| H. pylori | Helicobacter pylori | M.smegmatis | Mycolicibacterium smegmatis |
| S. iniae | Streptococcus iniae | V.ichthyoenteri | Vibrio ichthyoenteri |
| B. megaterium | Bacillus megaterium | S.tritici | Septoria tritici |
| T. rubrum | Trichophyton rubrum | S.lentus | Staphylococcus lentus |
| H. aquamarina | Halomonas aquamarina | P.irgensii | Polaribacter irgensii |
| P. elyakovii | Pseudoalteromonas elyakovii | R.litoralis | Roseobacter litoralis |
| S. putrefaciens | Shewanella putrefaciens | V.natriegens | Vibrio natriegens |
| V. carchariae | Vibrio carchariae | WTSA | wild-type Staphylococcus aureus |
| S. lutea | Sarcina lutea | M.tetragenus | Micrococcus tetragenus |
| V. anguillarum | Vibrio anguillarum | M.phlei | Mycobacterium phlei |
| S. albus | Streptomyces albus | C.glabrata | Candida glabrata |
| P. acnes | Propionibacterium acnes | X.campestris | Xanthomonas campestris |
| A. alternata | Alternaria alternata | A.hydrophila | Aeromonas hydrophila |
| G. gramini | Gaeumannomyces gramini | S.agalactiae | Streptococcus agalactiae |
| OSMAC | One Strain Many Compounds | MRCNS | meticillin-resistant coagulase-negative Staphylococcus |
| B. mycoides | Bacillus mycoides | R.baikonurensis | Rhodococcus baikonurensis |
| IC50 | half-inhibitory concentration | MIC | minimum inhibitory concentration |
| QM-NMR | quantum mechanical nuclear magnetic resonance | TDDFT | time-dependent density functional theory |
| MS | mass spectrometry | ECD | electronic circular dichroism |
| BTL-I | Butyrolactone I | MptpB | Mycobacterium tuberculosis protein tyrosine phosphatase B |
| KEGG | Kyoto Encylopaedia of Genes and Genomes | UHPLC-Q-TOF-MS | ultra-high-performance liquid chromatographyqua-drupole time-of-flight mass spectrometry |
| GO | Gene Ontology | ADME | adsorption, distribution, metabolism and excretion |
References
- Kiran, G.S.; Sekar, S.; Ramasamy, P.; Thinesh, T.; Hassan, S.; Lipton, A.N.; Ninawe, A.S.; Selvin, J. Marine sponge microbial association: Towards disclosing unique symbiotic interactions. Mar. Environ. Res. 2018, 140, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.R.A.; Kavlekar, D.P.; LokaBharathi, P.A. Marine drugs from sponge-microbe association—A review. Mar. Drugs 2010, 8, 1417–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slaby, B.M.; Hackl, T.; Horn, H.; Bayer, K.; Hentschel, U. Metagenomic binning of a marine sponge microbiome revealsunity in defense but metabolic specialization. ISME J. 2017, 11, 2465–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegl, A.; Hentschel, U. PKS and NRPS gene clusters from microbial symbiont cells of marine sponges by whole genome amplification. Env. Microbiol. Rep. 2010, 2, 507–513. [Google Scholar] [CrossRef]
- Hentschel, U.; Piel, J.; Degnan, S.M.; Taylor, M.W. Genomic insights into the marine sponge microbiome. Nat. Rev. Microbiol 2012, 10, 641–654. [Google Scholar] [CrossRef]
- Graca, A.P.; Viana, F.; Bondoso, J.; Correia, M.I.; Gomes, L.; Humanes, M.; Reis, A.; Xavier, J.R.; Gaspar, H.; Lage, O.M. The antimicrobial activity of heterotrophic bacteria isolated from the marine sponge Erylus deficiens (Astrophorida, Geodiidae). Front. Microbiol. 2015, 6, 389. [Google Scholar]
- Indraningrat, A.A.G.; Smidt, H.; Sipkema, D. Bioprospecting sponge-associated microbes for antimicrobial compounds. Mar. Drugs 2016, 14, 87. [Google Scholar] [CrossRef]
- Bibi, F.; Faheem, M.; Azhar, E.I.; Yasir, M.; Alvi, S.A.; Kamal, M.A.; Ullah, I.; Naseer, M.I. Bacteria from marine sponges: A source of new drugs. Curr. Drug Metab. 2017, 18, 11–15. [Google Scholar] [CrossRef]
- Skariyachan, S.; Rao, A.G.; Patil, M.R.; Saikia, B.; Bharadwaj, K.N.V.; Rao, G.S.J. Antimicrobial potential of metabolites extracted from bacterial symbionts associated with marine sponges in coastal area of Gulf of Mannar Biosphere, India. Lett. Appl. Microbiol. 2014, 58, 231–241. [Google Scholar] [CrossRef]
- Skariyachan, S.; Acharya, A.B.; Subramaniyan, S.; Babu, S.; Kulkarni, S.; Narayanappa, R. Secondary metabolites extracted from marine sponge associated Comamonas testosteroni and Citrobacter freundii as potential antimicrobials against MDR pathogens and hypothetical leads for VP40 matrix protein of Ebola virus: An in vitro and in silico investigation. J. Biomol. Struct. Dyn. 2016, 34, 1865–1883. [Google Scholar]
- Mayer, A.M.S.; Rodriguez, A.D.; Berlinck, R.G.S.; Hamann, M.T. Marine pharmacology: Marine compounds with anthelmintic, antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Biochim. Biophys. Acta 2009, 1790, 283–308. [Google Scholar]
- Mehbub, M.F.; Perkins, M.V.; Zhang, W.; Franco, C.M.M. New marine natural products from sponges (Porifera) of the order dictyoceratida (2001 to 2012); a promising source for drug discovery, exploration and future prospects. Biotechnol. Adv. 2016, 34, 473–491. [Google Scholar] [CrossRef]
- Jin, L.M.; Quan, C.S.; Hou, X.Y.; Fan, S.D. Potential pharmacological resources: Natural bioactive compounds from marine-derived fungi. Mar. Drugs 2016, 14, 76. [Google Scholar] [CrossRef] [Green Version]
- Gulder, T.A.M.; Hong, H.; Correa, J.; Egereva, E.; Wiese, J.; Imhoff, J.F.; Gross, H. Isolation, structure elucidation and total synthesis of lajollamide from the marine fungus Asteromyces cruciatus. Mar. Drugs 2012, 10, 2912–2935. [Google Scholar] [CrossRef]
- Li, D.; Xu, Y.; Shao, C.L.; Yang, R.Y.; Zheng, C.J.; Chen, Y.Y.; Fu, X.M.; Qian, P.Y.; She, Z.G.; De, V.N.J. Antibacterial bisabolane-type sesquiterpenoids from the sponge-derived fungus Aspergillus sp. Mar. Drugs 2012, 10, 234–241. [Google Scholar] [CrossRef] [Green Version]
- Sibero, M.T.; Zhou, T.; Fukaya, K.; Urabe, D.; Radjasa, O.K.K.; Sabdono, A.; Trianto, A.; Igarashi, Y. Two new aromatic polyketides from a sponge-derived Fusarium. Beilstein J. Org. Chem. 2019, 15, 2941–2947. [Google Scholar] [CrossRef] [Green Version]
- Prompanya, C.; Dethoup, T.; Bessa, L.J.; Pinto, M.M.M.; Gales, L.; Costa, P.M.; Silva, A.M.S.; Kijjoa, A. New isocoumarin derivatives and meroterpenoids from the marine sponge-associated fungus Aspergillus similanensis sp. nov. KUFA0013. Mar. Drugs 2014, 12, 5160–5173. [Google Scholar] [CrossRef] [Green Version]
- Kumla, D.; Aung, T.S.; Buttachon, S.; Dethoup, T.; Gales, L.; Pereira, J.A.; Inácio, A.; Costa, P.M.; Lee, M.; Sekeroglu, N.; et al. A new dihydrochromone dimer and other secondary metabolites from cultures of the marine sponge-associated fungi Neosartorya fennelliae KUFA0811 and Neosartorya tsunodae KUFC9213. Mar. Drugs 2017, 15, 375. [Google Scholar] [CrossRef] [Green Version]
- Khamthong, N.; Rukachaisirikul, V.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. An antibacterial cytochalasin derivative from the marine-derived fungus Diaporthaceae sp. PSU-SP2/4. Phytochem. Lett. 2014, 10, 5–9. [Google Scholar] [CrossRef]
- Zhou, Y.M.; Debbab, A.; Wray, V.; Lin, W.H.; Schulz, B.; Trepos, R.; Pile, C.; Hellio, C.; Proksch, P.; Aly, A.H. Marine bacterial inhibitors from the sponge-derived fungus Aspergillus sp. Tetrahedron Lett. 2014, 55, 2789–2792. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.D.; Huang, X.L.; Ma, Q.Y.; Xie, Q.Y.; Wang, P.; Chen, P.W.; Zhou, L.M.; Yuan, J.Z.; Dai, H.F.; Luo, D.Q.; et al. Helvolic acid derivatives with antibacterial activities against Streptococcus agalactiae from the marine-derived fungus Aspergillus fumigatus HNMF0047. J. Nat. Prod. 2018, 81, 1869–1876. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.P.; Wang, H.K.; Wu, J.L.; Shao, P.; Zhou, W.L.; Lai, Q.L.; Lin, H.W.; Naman, C.B.; Wang, T.T.; He, S. Acremocholone, an anti-vibrio steroid from the marine mesophotic zone ciocalypta sponge-associated fungus Acremonium sp. NBUF150. Chem. Biodivers. 2022, 19, e202200028. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.F.; Lin, X.P.; Qin, C.; Liao, S.R.; Wan, J.T.; Zhang, T.Y.; Liu, J.; Fredimoses, M.; Chen, H.; Yang, B.; et al. Antimicrobial and antiviral sesquiterpenoids from sponge-associated fungus, Aspergillus sydowii ZSDS1-F6. J. Antibiot. 2014, 67, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.J.; Cheng, W.; Heydari, H.; Wang, B.; Zhu, K.; Konuklugil, B.; Lin, W.H. Sorbicillinoid-based metabolites from a sponge-derived fungus Trichoderma saturnisporum. Mar. Drugs 2018, 16, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.J.; Gu, B.B.; Jiao, W.H.; Yuan, W.; Li, Y.X.; Tang, W.Z.; Yu, H.B.; Liao, X.J.; Han, B.N.; Li, Z.Y. New furan and cyclopentenone derivatives from the sponge-associated fungus Hypocrea Koningii PF04. Mar. Drugs 2015, 13, 5579–5592. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Ding, L.J.; Wei, J.X.; Li, Q.; Gui, M.J.; He, X.P.; Su, D.Q.; He, S.; Jin, H.X. A new aquatic pathogen inhibitor produced by the marine fungus Aspergillus sp. LS116. Aquaculture 2019, 520, 734665–734670. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, L.J.; Fang, F.; He, S. Penicillilactone A, a novel antibacterial 7-membered lactone derivative from the sponge-associated fungus Penicillium sp. LS54. Nat. Prod. Res. 2019, 33, 2466–2470. [Google Scholar] [CrossRef]
- Bovio, E.; Garzoli, L.; Poli, A.; Luganini, A.; Villa, P.; Musumeci, R.; Mccormack, G.P.; Cocuzza, C.E.; Gribaudo, G.; Mehiri, M.; et al. Marine fungi from the sponge Grantia compressa: Biodiversity, chemodiversity, and biotechnological potential. Mar. Drugs 2019, 17, 220. [Google Scholar] [CrossRef] [Green Version]
- Pang, X.Y.; Cai, G.D.; Lin, X.P.; Salendra, L.; Zhou, X.F.; Yang, B.; Wang, J.J.; Wang, J.F.; Xu, S.H.; Liu, Y.H. New alkaloids and polyketides from the marine sponge-derived fungus Penicillium sp. SCSIO41015. Mar. Drugs 2019, 17, 398. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Li, Y.; Guo, X.C.; Ji, W.; Lin, W.H. Chartarlactams Q–T, dimeric phenylspirodrimanes with antibacterial and antiviral activities. Chem. Biodivers. 2020, 17, e2000170. [Google Scholar] [CrossRef]
- Yamazaki, H.; Rotinsulu, H.; Kaneko, T.; Murakami, K.; Fujiwara, H.; Ukai, K.; Namikoshi, M. A new dibenz[b,e]oxepine derivative,1-Hydroxy-10-methoxy-dibenz[b,e]oxepin-6,11-dione, from a marine-derived fungus, Beauveria bassiana TPU942. Mar. Drugs 2012, 10, 2691–2697. [Google Scholar] [CrossRef] [Green Version]
- Bunyapaiboonsri, T.; Yoiprommarat, S.; Intereya, K.; Kocharin, K. New diphenyl ethers from the insect pathogenic fungus Cordyceps sp. BBC 1861. Chem. Pharm. Bull. 2007, 55, 304–307. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.L.; Ma, X.H.; Xie, Y.Y.; Cai, S.X.; Zhu, T.J.; Gu, Q.Q.; Li, D.H. Aromatic polyketides from a sponge-derived fungus Metarhizium anisopliae MXH-99 and their antitubercular activities. Arch. Pharm. Res. 2013, 36, 739–744. [Google Scholar] [CrossRef]
- Subramani, R.; Kumar, R.; Prasad, P.; Aalbersberg, W.; Retheesh, S.T. Cytotoxic and antibacterial substances against multi-drug resistantpathogens from marine sponge symbiont: Citrinin, a secondarymetabolite of Penicillium sp. Asian. Pac. J. Trop. Biomed. 2013, 3, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Shao, C.L.; Li, Z.Y.; Gan, L.S.; Fu, X.M.; Bian, W.T.; Zhao, H.Y.; Wang, C.Y. Isocoumarin derivatives and benzofurans from a sponge-derived Penicillium sp. fungus. J. Nat. Prod. 2013, 76, 571–579. [Google Scholar] [CrossRef]
- Jansen, N.; Ohlendorf, B.; Erhard, A.; Bruhn, T.; Bringmann, G.; Imhoff, J.F. Helicusin E, isochromophilone X and isochromophilone Xl: New chloroazaphilonesproduced by the fungus Bartalinia robillardoides strain LF550. Mar. Drugs 2013, 11, 800–816. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Ohlendorf, B.; Oesker, V.; Wiese, J.; Malien, S.; Schmaljohann, R.; Imhoff, J.F. Acetylcholinesterase inhibitors from a marine fungus Talaromyces sp. strain LF458. Mar. Biotechnol. 2015, 17, 110–119. [Google Scholar] [CrossRef]
- Wu, Z.H.; Liu, D.; Huang, J.; Proksch, P.; Zhu, K.; Lin, W.H. Hansforesters A-M, polyesters from the sponge-associated fungus Hansfordia sinuosae with antibacterial activities. RSC. Adv. 2018, 8, 39756–39768. [Google Scholar] [CrossRef] [Green Version]
- Lei, H.; Lin, X.P.; Han, L.; Ma, J.; Ma, Q.J.; Zhong, J.L.; Liu, Y.H.; Sun, T.M.; Wang, J.H.; Huang, X.S. New metabolites and bioactive chlorinated benzophenone derivatives produced by a marine-derived fungus Pestalotiopsis heterocornis. Mar. Drugs 2017, 15, 68–69. [Google Scholar] [CrossRef] [Green Version]
- Lei, H.; Lin, X.P.; Han, L.; Ma, J.; Dong, K.L.; Wang, X.B.; Zhong, J.L.; Mu, Y.; Liu, Y.H.; Huang, X.S. Polyketide derivatives from a marine sponge-associated fungus Pestalotiopsis heterocornis. Phyto. Chem. 2017, 142, 51–59. [Google Scholar] [CrossRef]
- Ding, L.J.; Yuan, W.; Li, Y.X.; Liao, X.J.; Sun, H.; Peng, Q.; Han, B.N.; Lin, H.W.; Li, Z.Y.; Yang, F.; et al. Hypocrol A, a new tyrosol derivative from a sponge-derived strain of the fungus Hypocrea koningii. Nat. Prob. Res. 2016, 30, 1633–1638. [Google Scholar] [CrossRef] [PubMed]
- Zhen, X.; Gong, T.; Wen, Y.H.; Yan, D.J.; Chen, J.J.; Zhu, P. Chrysoxanthones A–C, three new xanthone-chromanone heterdimers from sponge-associated Penicillium chrysogenum HLS111 treated with histone deacetylase lnhibitor. Mar. Drugs 2018, 16, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabdaningsih, A.; Liu, Y.; Mettal, U.; Heep, J.; Riyanti; Wang, L.; Cristianawati, O.; Nuryadi, H.; Sibero, M.T.; Marner, M.; et al. A new citrinin derivative from the indonesian marine sponge-associated fungus Penicillium citrinum. Mar. Drugs 2020, 18, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asiri, I.A.M.; Badr, J.M.; Youssef, D.T.A. Penicillivinacine, antimigratory diketopiperazine alkaloid from the marine-derived fungus Penicillium vinaceum. Phytochem. Lett. 2015, 13, 53–58. [Google Scholar] [CrossRef]
- Song, F.H.; Ren, B.; Chen, C.X.; Yu, K.; Liu, X.R.; Zhang, Y.H.; Yang, N.; He, H.T.; Liu, X.T.; Dai, H.Q.; et al. Three new sterigmatocystin analogues from marine-derived fungus Aspergillus versicolor MF359. Appl. Microbiol. Biotechnol. 2014, 98, 3753–3758. [Google Scholar] [CrossRef]
- Song, Z.J.; Liu, Y.; Gao, J.Y.; Hu, J.S.; He, H.T.; Dai, S.W.; Wang, L.Q.; Dai, H.Q.; Zhang, L.X.; Song, F.H. Antitubercular metabolites from the marine-derived fungus strain Aspergillus fumigatus MF029. Nat. Prod. Res. 2021, 35, 2647–2654. [Google Scholar] [CrossRef]
- Wu, B.; Wiese, J.; Labes, A.; Kramer, A.; Schmaljohann, R.; Imhoff, J.F. Lindgomycin, an unusual antibiotic polyketide from a marine fungus of the Lindgomycetaceae. Mar. Drugs 2015, 13, 4617–4632. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.; Sun, C.X.; Sun, Z.C.; Zhang, G.J.; Che, Q.; Gu, Q.Q.; Zhu, T.J.; Li, D.H. Antibacterial polyketides from antarctica sponge-derived fungus Penicillium sp. HDN151272. Mar. Drugs 2020, 18, 71. [Google Scholar] [CrossRef] [Green Version]
- Kumla, D.; Pereira, J.A.; Dethoup, T.; Gales, L.; Freitas-Silva, J.; Costa, P.M.; Lee, M.; Silva, A.M.S.; Sekeroglu, N.; Pinto, M.M.M.; et al. Chromone derivatives and other constituents from cultures of the marine sponge-associated fungus Penicillium erubescens KUFA0220 and their antibacterial activity. Mar. Drugs 2018, 16, 289. [Google Scholar] [CrossRef] [Green Version]
- Machado, F.P.; Kumla, D.; Pereira, J.A.; Sousa, E.; Dethoup, T.; Freitas-Silva, J.; Costa, P.M.; Mistry, S.; Silva, A.M.S.; Kijjoa, A. Prenylated phenylbutyrolactones from cultures of a marine sponge-associated fungus Aspergillus flavipes KUFA1152. Phytochemistry 2021, 185, 112709. [Google Scholar] [CrossRef]
- Sá, J.D.M.; Pereira, J.A.; Dethoup, T.; Cidade, H.; Sousa, M.E.; Rodrigues, I.C.; Costa, P.M.; Mistry, S.; Silva, A.M.S.; Kijjoa, A. Anthraquinones, diphenyl ethers, and their derivatives from the culture of the marine sponge-associated fungus Neosartorya spinosa KUFA 1047. Mar. Drugs 2021, 19, 457. [Google Scholar]
- Ozkaya, F.C.; Ebrahim, W.; El-Neketi, M.; Tanrikul, T.T.; Kalscheuer, R.; Muller, W.E.G.; Guo, Z.Y.; Zou, K.; Liu, Z.; Proksch, P. Induction of new metabolites from sponge-associated fungus Aspergillus carneus by OSMAC approach. Fitoterapia 2018, 131, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Pilevneli, A.D.; Ebada, S.S.; Kaskatepe, B.; Konuklugil, B. Penicacids H-J, three new mycophenolic acid derivatives from the marine-derived fungus Rhizopus oryzae. RSC. Adv. 2021, 11, 34938–34944. [Google Scholar] [CrossRef]
- Li, W.; Ding, L.J.; Wang, N.; Xu, J.Z.; Zhang, W.Y.; Zhang, B.; He, S.; Wu, B.; Jin, H.X. Isolation and characterization of two new metabolites from the sponge-derived fungus Aspergillus sp. LS34 by OSMAC approach. Mar. Drugs 2019, 17, 283. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ding, L.J.; Zhang, Z.M.; Yan, X.J.; He, S. New antifungal tetrahydrofuran derivatives from a marine sponge-associated fungus Aspergillus sp. LS78. Fitoterapia 2020, 146, 104672–104677. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, L.J.; He, J.X.; Zhang, Z.M.; Deng, Y.T.; He, S.; Yan, X.J. A new antibacteriail chromone from a marine sponge-associated fungus Aspergils sp. LS57. Fitoterapia 2021, 154, 105004. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, X.P.; Ju, Z.R.; Liao, X.J.; Huang, X.J.; Zhang, C.; Zhao, B.X.; Xu, S.H. Aspergchromones A and B, two new polyketides from the marine sponge-associated fungus Aspergillus sp. SCSIOXWS03F03. J. Asian Nat. Prob. Res. 2016, 19, 684–690. [Google Scholar] [CrossRef]
- Liu, J.; Yu, R.L.; Jia, J.; Gu, W.; Zhang, H.W. Assignment of absolute configurations of two promising anti-Helicobacter pylori agents from the marine sponge-derived fungus Aspergillus niger L14. Molecules 2021, 26, 5061. [Google Scholar] [CrossRef]
- Ding, L.; Ren, L.; Li, S.; Song, J.J.; Han, Z.W.; He, S.; Xu, S.H. Production of new antibacterial 4-Hydroxy-α-Pyrones by a marine fungus Aspergillus niger cultivated in solid medium. Mar. Drugs 2019, 17, 344. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.Q.; Lin, S.T.; Kumaravel, K.; Zhou, H.; Wang, S.Y.; Liu, Y.H. Polyketide-derived metabolites from the sponge-derived fungus Aspergillus sp. F40. Phytochem. Lett. 2018, 27, 74–77. [Google Scholar] [CrossRef]
- Pang, X.Y.; Lin, X.P.; Wang, P.; Zhou, X.F.; Yang, B.; Wang, J.F.; Liu, Y.H. Perylenequione derivatives with anticancer activities isolated from the marine sponge-derived fungus, Alternaria sp. SCSIO41014. Mar. Drugs 2018, 16, 280. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.M.; Luo, X.W.; Li, K.L.; Guo, C.; Li, J.X.; Lin, X.P.; Liu, Y.H. Antibacterial secondary metabolites from a marine sponge-derived fungus Fusarium equiseti SCSIO 41019. China Antibiot. J. 2019, 44, 1035–1040. [Google Scholar]
- Chen, Y.P.; Liu, C.N.; Kumaravel, K.; Nan, L.H.; Tian, Y.Q. Two new sulfate-modified dibenzopyrones with anti-foodborne bacteria activity from sponge-derived fungus Alternaria sp. SCSIOS02F49. Front. Microbiol. 2022, 13, 879674. [Google Scholar] [CrossRef] [PubMed]
- Saetang, P.; Rukachaisirikul, V.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J.; Hadsadee, S.; Jungsuttiwong, S. Antibacterial and antifungal polyketides from the fungus Aspergillus unguis PSU-MF16. J. Nat. Prod. 2021, 84, 1498–1506. [Google Scholar] [CrossRef]
- Liu, S.; Wang, H.B.; Su, M.Z.; Hwang, G.J.; Hong, J.; Jung, J.H. New metabolites from the sponge-derived fungus Aspergillus sydowii J05B-7F-4. Nat. Prob. Res. 2017, 31, 1682–1686. [Google Scholar] [CrossRef] [PubMed]
- Haga, A.; Tamoto, H.; Ishino, M.; Kimura, E.; Sugita, T.; Kinoshita, K.; Takahashi, K.; Shiro, M.; Koyama, K. Pyridone alkaloids from a marine-derived fungus, Stagonosporopsis cucurbitacearum, and their activities against azole-resistant Candida albicans. J. Nat. Prod. 2013, 76, 750–754. [Google Scholar] [CrossRef]
- Shaala, L.A.; Alzughaibi, T.; Genta-Jouve, G.; Youssef, D.T.A. Fusaripyridines A and B, highly oxygenated antimicrobial alkaloid dimers featuring an unprecedented 1,4-bis(2-hydroxy-1,2-dihydropyridin-2-yl)butane-2,3-dione core from the marine fungus Fusarium sp. LY019. Mar. Drugs 2021, 19, 505. [Google Scholar] [CrossRef]
- Liu, Y.; Li, X.M.; Meng, L.Y.; Wang, B.G. N-Formyllapatin A, a new N-formylspiroquinazoline derivative from the marine-derived fungus Penicillium adametzioides AS-53. Phytochem. Lett. 2014, 10, 145–148. [Google Scholar] [CrossRef]
- Liu, Y.; Li, X.M.; Meng, L.H.; Jiang, W.L.; Xu, G.M.; Huang, C.G.; Wang, B.G. Bisthiodiketopiperazines and acorane sesquiterpenes produced by the marine-derived fungus Penicillium adametzioides AS-53 on different culture media. J. Nat. Prod. 2015, 78, 1294–1299. [Google Scholar] [CrossRef]
- Wang, C.Y.; Liu, X.H.; Zheng, Y.Y.; Ning, X.Y.; Zhang, Y.H.; Fu, X.M.; Li, X.; Shao, C.L.; Wang, C.Y. 2,5-Diketopiperazines from a sponge-derived fungus Aspergillus sclerotiorum. Front. Microbiol. 2022, 13, 808532. [Google Scholar] [CrossRef]
- Buttachon, S.; Ramos, A.A.; Inacio, A.; Dethoup, T.; Gales, L.; Lee, M.; Costa, P.M.; Silva, A.M.S.; Sekeroglu, N.; Rocha, E.; et al. Bis-Indolyl benzenoids, hydroxypyrrolidine derivatives and other constituents from cultures of the marine sponge-associated fungus Aspergillus candidus KUFA0062. Mar. Drugs 2018, 16, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Ding, L.J.; Shi, Y.T.; Yan, X.J.; Wu, B.; He, S. Molecular networking-driven discovery of antibacterial perinadines, new tetracyclic alkaloids from the marine sponge-derived fungus Aspergillus sp. ACS Omega 2022, 7, 9909–9916. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Qiu, P.P.; Zhang, Y.Y.; Ding, L.J.; He, S. Isolation and structure elucidation of antibacterial metabolites from a sponge- derived fungus Penicillium chrysogenum LS16. Mygosystema 2018, 37, 640–648. [Google Scholar]
- Scopel, M.; Abraham, W.R.; Henriques, A.T.; Macedo, A.J. Dipeptide cis-cyclo (Leucyl-Tyrosyl) produced by sponge associated Penicillium sp. F37 inhibits biofilm formation of the pathogenic Staphylococcus epidermidis. Bioorg. Med. Chem. Lett. 2013, 23, 624–626. [Google Scholar] [CrossRef]
- Liu, Y.; Mandi, A.; Li, X.M.; Meng, L.H.; Kurtan, T.; Wang, B.G. Peniciadametizine A, a dithiodiketopiperazine with a unique spiro[furan-2,7 ’-pyrazino[1,2-b][1,2]oxazine] skeleton, and a related analogue, peniciadametizine B, from the marine sponge-derived fungus Penicillium adametzioides. Mar. Drugs 2015, 13, 3640–3652. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.X.; Zhang, Z.P.; Ren, Z.L.; Yu, L.; Zhou, H.; Han, Y.X.; Shah, M.; Che, Q.; Zhang, G.J.; Li, D.H.; et al. Antibacterial cyclic tripeptides from Antarctica-sponge-derived fungus Aspergillus insulicola HDN151418. Mar. Drugs 2020, 18, 532. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z. Cyclopeptide derivatives from the sponge-derived fungus Acremonium persicinum F10. Mar. Drugs 2021, 19, 537. [Google Scholar] [CrossRef]
- Yang, C.F.; Qian, R.; Xu, Y.; Yi, J.X.; Gu, Y.W.; Liu, X.Y.; Yu, H.B.; Jiao, B.H.; Lu, X.L.; Zhang, W. Marine actinomycetes-derived natural products. Curr. Top. Med. Chem. 2019, 19, 2868–2918. [Google Scholar] [CrossRef]
- Supong, K.; Thawai, C.; Suwanborirux, K. Antimalarial and antitubercular C-glycosylaed benz[α]anthraq-uinones from the marine-derived Streypomyeces sp. BCC45596. Phytochem. Lett. 2012, 5, 651–656. [Google Scholar] [CrossRef]
- Bruntner, C.; Binder, T.; Pathom-aree, W.; Goodfellow, M.; Bull, A.T.; Potterat, O.; Puder, C.; Horer, S.; Schmid, A.; Bolek, W.; et al. Frigocyclinone, a novel angucyclinone antibiotic produced by a Streptomyces griseus strain from Antarctica. J. Antibiot. 2005, 58, 346–349. [Google Scholar] [CrossRef] [Green Version]
- Vicente, J.; Stewart, A.K.; van Wagoner, R.M.; Elliott, E.; Bourdelais, A.J.; Wright, J.L.C. Monacyclinones, new angucyclinone metabolites isolated from Streptomyces sp. M7_15 associated with the Puerto Rican Sponge Scopalina ruetzleri. Mar. Drugs 2015, 13, 4682–4700. [Google Scholar] [CrossRef] [Green Version]
- Reimer, A.; Blohm, A.; Quack, T.; Grevelding, C.G.; Kozjak-Pavlovic, V.; Rudel, T.; Hentschel, U.; Abdelmohsen, U.R. Inhibitory activities of the marine Streptomycete-derived compound SF2446A2 against Chlamydia trachomatis and Schistosoma mansoni. J. Antibiot. 2015, 68, 674–679. [Google Scholar] [CrossRef] [Green Version]
- Cui, M.Z.; Gong, F.; Zhu, Q. Study on the active metabolites of Streptomyces LS298 associated with Spongy sponge. China Med. Biotechnol. 2012, 7, 418–419. [Google Scholar]
- Lacret, R.; Oves-Costales, D.; Pérez-Victoria, I.; de la Cruz, M.; Diaz, C.; Vicente, F.; Genilloud, O.; Reyes, F. MDN-0171, a new medermycin analogue from Streptomyces albolongus CA-186053. Nat. Prod. Res. 2019, 33, 66–73. [Google Scholar] [CrossRef]
- Shaala, L.A.; Youssef, D.T.A.; Alzughaibi, T.A.; Elhady, S.S. Antimicrobial chlorinated 3-phenylpropanoic acid derivatives from the Red Sea marine actinomycete Streptomyces coelicolor LY001. Mar. Drugs 2020, 18, 450. [Google Scholar] [CrossRef]
- Cao, D.D.; Trinh, T.T.V.; Mai, H.D.T.; Vu, V.N.; Le, H.M.; Vu Thi, Q.; Nguyen, M.A.; Duong, T.T.; Tran, D.T.; Chau, V.M.; et al. Antimicrobial lavandulylated flavonoids from a sponge-derived Streptomyces sp. G248 in east vietnam sea. Mar. Drugs 2019, 17, 529. [Google Scholar] [CrossRef] [Green Version]
- Cao, D.D.; Do, T.Q.; Mai, H.D.T.; Thi, Q.V.; Nguyen, M.A.; Thi, H.M.L.; Tran, D.T.; Chau, V.M.; Thung, D.C.; Pham, V.C. Antimicrobial lavandulylated flavonoids from a sponge-derived actinomycete. Nat. Prod. Res. 2020, 34, 413–420. [Google Scholar] [CrossRef]
- Singh, S.; Prasad, P.; Subramani, R.; Aalbersberg, W. Production and purification of a bioactive substance against multi-drug resistant human pathogens from the marine-sponge-derived Salinispora sp. Asian Pac. J. Trop. Biomed. 2014, 4, 825–831. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.B.; Nepa, K.K.; Chen, J.; Harmody, D.; Zhu, H.N.; McCarthy, P.J.; Wright, A.E.; Wang, G.J. Nocardiopsistins A-C: New angucyclines with anti-MRSA activity isolated from a marine sponge-derived Nocardiopsis sp.HB-J378. Synth. Syst. Biotechnol. 2018, 3, 246–251. [Google Scholar] [CrossRef]
- Chen, M.H.; Zhang, W.L.; Chen, L.; Lin, R.; Xie, Y.; Fang, D.S.; Lian, Y.Y.; Jiang, H. Isolation, purification and identification of two new alkaloids metabolites from marine-derived Verrucosispora sp. FIM06025. Nat. Prod. Res. 2019, 33, 2897–2903. [Google Scholar] [CrossRef]
- Chen, M.H.; Lian, Y.Y.; Fang, D.S.; Chen, L.; Jia, J.; Zhang, W.L.; Lin, R.; Xie, Y.; Bi, H.K.; Jiang, H. Identification and antimicrobial properties of a new alkaloid produced by marine-derived Verrucosispora sp. FIM06-0036. Nat. Prod. Res. 2019, 35, 4211–4217. [Google Scholar] [CrossRef]
- Huang, X.L.; Kong, F.D.; Zhou, S.Q.; Huang, D.Y.; Zheng, J.P.; Zhu, W.M. Streptomyces tirandamycinicus sp. nov., a novel marine sponge-derived actinobacterium with antibacterial potential against Streptococcus agalactiae. Front. Microbiol. 2019, 10, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Back, C.R.; Stennett, H.L.; Williams, S.E.; Wang, L.Y.; Gomez, J.O.; Abdulle, O.M.; Duffy, T.; Neal, C.; Mantell, J.; Jepson, M.A.; et al. A new Micromnonospora strain with antibiotic activity isolated from the microbiome of a Mid-Atlantic deep-sea sponge. Mar. Drugs 2021, 19, 105. [Google Scholar] [CrossRef]
- Elsayed, Y.; Refaat, J.; Abdelmohsen, U.R.; Ahmed, S.; Fouad, M.A. Rhodozepinone, a new antitrypanosomal azepino-diindole alkaloid from the marine sponge-derived bacterium Rhodococcus sp. UA13. Med. Chem. Res. 2017, 26, 2751–2760. [Google Scholar] [CrossRef]
- Shaala, L.A.; Youssef, D.T.A.; Badr, J.M.; Harakeh, S.M.; Genta-Jouve, G. Bioactive diketopiperazines and nucleoside derivatives from a sponge-derived Streptomyces Species. Mar. Drugs 2019, 17, 584. [Google Scholar] [CrossRef] [Green Version]
- Alkhalifah, D.H.M. Sponge-associated sp. RM66 metabolome induction with N-acetylglucosamine: Antibacterial, antifungal and anti-trypanosomal activities. Saudi. Biol. Sci. 2021, 28, 4691–4698. [Google Scholar] [CrossRef]
- Karuppiah, V.; Li, Y.X.; Sun, W.; Feng, G.F.; Li, Z.Y. Functional gene-based discovery of phenazines from the actinobacteria associated with marine sponges in the South China Sea. Appl. Microbiol. Biotechnol. 2015, 99, 5939–5950. [Google Scholar] [CrossRef]
- Zhen, X.; Gong, T.; Liu, F.; Zhang, P.C.; Zhou, W.Q.; Li, Y.; Zhu, P. A New analogue of echinomycin and a new cyclic dipeptide from a marine-derived Streptomyces sp. LS298. Mar. Drugs 2015, 13, 6947–6961. [Google Scholar] [CrossRef] [Green Version]
- Kitani, S.; Ueguchi, T.; Igarashi, Y.; Leetanasaksakul, K.; Thamchaipenet, A.; Nihira, T. Rakicidin F, a new antibacterial cyclic depsipeptide from a marine sponge-derived Streptomyces sp. J. Antibiot. 2018, 7, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Jiao, W.H.; Yuan, W.; Li, Z.Y.; Li, J.; Li, L.; Sun, J.B.; Gui, Y.H.; Wang, J.; Ye, B.P.; Lin, H.W. Anti-MRSA actinomycins D1-D4 from the marine sponge-associated Streptomyces sp. LHW52447. Tetrahedron 2018, 74, 5914–5919. [Google Scholar] [CrossRef]
- Sekurova, O.N.; Pérez-Victoria, I.; Martín, J.; Degnes, K.F.; Sletta, H.; Reyes, F.; Zotchev, S.B. New deferoxamine glycoconjugates produced upon overexpression of pathway-specific regulatory gene in the marine sponge-derived Streptomyces albus PVA94-07. Molecules 2016, 21, 1131. [Google Scholar] [CrossRef] [Green Version]
- Takasaka, N.; Kaweewan, I.; Ohnishi-Kameyama, M.; Kodani, S. Isolation of a new antibacterial peptide actinokineosin from Actinokineospora spheciospongiae based on genome mining. Lett. Appl. Microbiol. 2017, 64, 150–157. [Google Scholar] [CrossRef]
- Kiran, G.S.; Sajayan, A.; Priyadharshirni, G.; Balakrishnan, A.; Prathiviraj, R.; Sabu, A.; Selvin, J. A novel anti-infective molecule nesfactin identified from sponge associated bacteria Nesterenkonia sp. MSA31 against multidrug resistant Pseudomonas aeruginosa. Microt. Pathog. 2021, 157, 104923. [Google Scholar] [CrossRef]
- Naik, D.N.; Wahidullah, S.; Meena, R.M. Attenuation of Pseudomonas aeruginosa virulence by marine invertebrate-derived Streptomyces sp. Lett. Appl. Microbiol. 2013, 56, 197–207. [Google Scholar] [CrossRef]
- Palomo, S.; Gonzalez, I.; de la Cruz, M.; Martin, J.; Tormo, J.R.; Anderson, M.; Hill, R.T.; Vicente, F.; Reyes, F.; Genilloud, O. Sponge-derived Kocuria and Micrococcus sp. as sources of the new thiazolyl peptide antibiotic Kocurin. Mar. Drugs 2013, 11, 1071–1086. [Google Scholar] [CrossRef] [Green Version]
- Eltamany, E.E.; Abdelmohsen, U.R.; Ibrahim, A.K.; Hassanean, H.A.; Hentschel, U.; Ahmed, S.A. New antibacterial xanthone from the marine sponge-derived Micrococcus sp. EC45. Bioorg. Med. Chem. Lett. 2014, 24, 4939–4942. [Google Scholar] [CrossRef]
- Gong, T.; Zhen, X.; Li, X.L.; Chen, J.J.; Chen, T.J.; Yang, J.L.; Zhu, P. Tetrocarcin Q, a new spirotetronate with a unique glycosyl group from a marine-derived actinomycete Micromonospora carbonacea LS276. Mar. Drug 2018, 16, 74. [Google Scholar] [CrossRef] [Green Version]
- Dat, T.T.H.; Cuc, N.T.K.; Cuong, P.V.; Smidt, H.; Sipkema, D. Diversity and antimicrobial activity of Vietnamese sponge-associated bacteria. Mar. Drugs 2021, 19, 353. [Google Scholar] [CrossRef]
- Santos, O.C.S.; Soares, A.R.; Machado, F.L.S.; Romanos, M.T.V.; Muricy, G.; Giambiagi-deMarval, M.; Laport, M.S. Investigation of biotechnological potential of sponge-associated bacteria collected in Brazilian coast. Lett. Appl. Microbiol. 2015, 60, 140–147. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Lai, T.K.; Saha, A.; Selvin, J.; Mukherjee, J. Structural elucidation and antimicrobial activity of a diketopiperazine isolated from a Bacillus sp. associated with the marine sponge Spongia officinalis. Nat. Prod. Res. 2021, 35, 2315–2323. [Google Scholar] [CrossRef]
- Yan, J.X.; Wu, Q.; Helfrich, E.J.N.; Chevrette, M.G.; Braun, D.R.; Heyman, H.; Ananiev, G.E.; Rajski, S.R.; Currie, C.R.; Clardy, J.; et al. Bacillimidazoles A-F, imidazolium-containing compounds isolated from a marine Bacillus. Mar. Drugs 2022, 20, 43. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.X.; Chevrette, M.G.; Braun, D.R.; Harper, M.K.; Currie, C.R.; Bugni, T.S. Madurastatin D1 and D2, oxazoline containing siderophores isolated from an Actinomadura sp. Org. Lett. 2019, 21, 6275–6279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Alvarenga, R.F.R.; Throckmorton, K.; Chanana, S.; Braun, D.R.; Fossen, J.; Zhao, M.; McCrone, S.; Harper, M.K.; Rajski, S.R.; et al. Genome mining and metabolomics unveil pseudonochelin: A siderophore containing 5-aminosalicylate from a marine-derived Pseudonocardia sp. bacterium. Org. Lett. 2022, 24, 3998–4002. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahab, N.M.; Scharf, S.; Ozkaya, F.C.; Kurtan, T.; Mandi, A.; Fouad, M.A.; Kamel, M.S.; Muller, W.E.G.; Kalscheuer, R.; Lin, W.H.; et al. Induction of secondary metabolites from the marine-derived fungus Aspergillus versicolor through co-cultivation with Bacillus subtilis. Planta Med. 2019, 85, 503–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, X.M.; Li, S.S.; Ni, J.; Wang, G.Y.; Li, F.; Li, Q.; Chen, S.Z.; Shu, J.C.; Gan, M.L. Acremopeptaibols A-F, 16-residue peptaibols from the sponge-derived Acremonium sp. IMB18-086 cultivated with heat-killed Pseudomonas aeruginosa. J. Nat. Prod. 2021, 84, 2990–3000. [Google Scholar] [CrossRef]
- Dashti, Y.; Grkovic, T.; Abdelmohsen, U.R.; Hentschel, U.; Quinn, R.J. Production of induced secondary metabolites by a co-culture of sponge-associated actinomycetes, Actinokineospora sp. EG49 and Nocardiopsis sp. RV163. Mar. Drugs 2014, 12, 3046–3059. [Google Scholar] [CrossRef]
- Hifnawy, M.S.; Hossan, H.M.; Mohammed, R.; Fouda, M.M.; Sayed, A.M.; Hamed, A.A.; AbouZid, S.F.; Rateb, M.E.; Alhadrami, H.A.; Abdelmohsen, U.R. Induction of antibacterial metabolites by co-cultivation of two Red-Sea-sponge-associated actinomycetes Micromonospora sp. UR56 and Actinokinespora sp. EG49. Mar. Drugs 2020, 18, 243. [Google Scholar] [CrossRef]
- Malhao, F.; Ramos, A.A.; Buttachon, S.; Dethoup, T.; Kijjoa, A.; Rocha, E. Cytotoxic and antiproliferative effects of preussin, a hydroxypyrrolidine derivative from the marine sponge-associated fungus Aspergillus candidus KUFA0062, in a panel of breast cancer cell lines and using 2D and 3D cultures. Mar. Drugs 2019, 17, 448. [Google Scholar] [CrossRef] [Green Version]
- Pang, M.J.; Yang, Z.; Zhang, X.L.; Liu, Z.F.; Fan, J.; Zhang, H.Y. Physcion, a naturally occurring anthraquinone derivative, induces apoptosis and autophagy in human nasopharyngeal carcinoma. Acta. Pharmacol. Sin. 2016, 37, 1623–1640. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Y.; Hu, H.L.; Liu, J.Q.; Chen, J.Q.; Chu, J.; Cheng, H. Physcion, a novel anthraquinone derivative against Chlamydia psittaci infection. Vet. Microbiol. 2023, 279, 109664. [Google Scholar] [CrossRef]
- Oliveira, K.D.; Queiroz, P.R.M.; Fensterseifer, I.C.M.; Migliolo, L.; Oliveira, A.L.; Franco, O.L. Purified citritin in combination with vancomycin inhibits VRE in vitro and in vivo. Microbiology 2017, 163, 1525–1531. [Google Scholar] [CrossRef]
- Luo, X.W.; Lin, Y.; Lu, Y.J.; Zhou, X.F.; Liu, Y.H. Peptides and polyketides isolated from the marine sponge-derived fungus Aspergillus terreus SCSIO 41008. Chin. J. Nat. Med. 2019, 17, 149–154. [Google Scholar] [CrossRef]
- Wu, L.; Xie, C.L.; Yang, X.W.; Chen, G. Pharmacokinetics and metabolism study of deep-sea-derived butyrolactone I in rats by UHPLC-MS/MS and UHPLC-Q-TOF-MS. Mar. Drugs 2022, 20, 11. [Google Scholar] [CrossRef]
- Gu, T.W.; Chen, W.H.; She, J.L.; Yang, B.; Tang, L.; Tao, H.M.; Huang, J.X.; Zhou, X.F. Lipid metabolism regulatory activity and adverse effects of fungi-derived butyrolactone I. Nat. Prod. Res. 2022. [Google Scholar] [CrossRef]









































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sponge Origin | Compounds | Antimicrobial Activity |
|---|---|---|
| Agelas oroides | 102, 103, 104 | Enterococcus faecalis (E. faecalis), Staphylococcus aureus (S. aureus), Enterococcus faecium (E. faecium) |
| 259, 260, 261, 262 | E. faecalis, S. aureus, E. faecium, Fusarium solani (F. solani), Escherichia coli (E. coli), Bacillus subtilis (B. subtilis) | |
| Amphimedon sp. | 223, 224 | B. subtilis, E. coli, Candida albicans (C.albicans) |
| Axinella sp. | 168 | Staphylococcus epidermidis (S. epidermidis) |
| 48, 49, 50 | S. epidermidis, methicillin-resistant Staphylococcus aureus (MRSA) | |
| Callyspongia sp. | 122, 123, 124 | S. aureus, Vibrio parahaemolyticus (V. parahaemolyticus) |
| 125, 214 | S. aureus | |
| 30, 31, 183 | S. aureus, Acinetobacter baumannii (A. baumannii) | |
| 126, 146, 147 | S. aureus, MRSA | |
| 127-136, 184, 185 | E. coli, Vibrio proteolyticus (V. proteolyticus), Proteus mirabilis (P. mirabilis), Pseudomonas fluorescens (P. fluorescens), Shigella flexneri (S. flexneri), Salmonella cholerasuis (S. cholerasuis), Listeria monocytogenes (L. monocytogenes), S. aureus | |
| 215, 216, 217, 218 | S. aureus, C. albicans | |
| 194–197, 219, 220 | S. aureus, E. coli, C. albicans | |
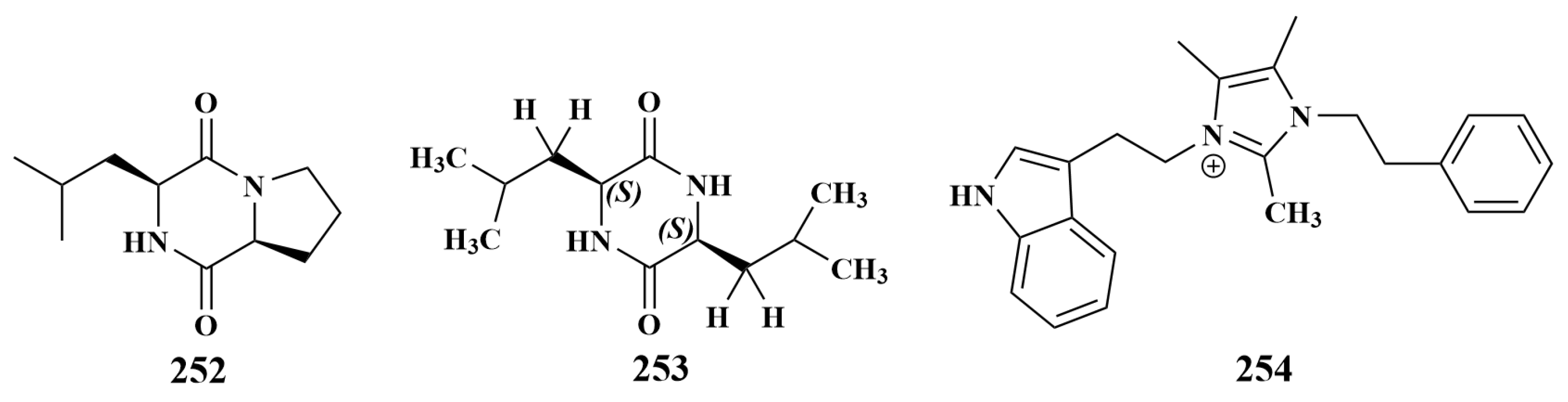
| Cinachyrella apion | 254 | MRSA |
| Ciocalypta sp. | 17, 18, 19 | Vibrio scophthalmi (V. scophthalmi), Vibrio shilonii (V. shilonii), Vibrio brasiliensis (V. brasiliensis) |
| Clathria reinwardtii | 9, 143 | S. aureus, vancomycin-resistant Enterococcus (VRE) E. faecalis, E. faecalis |
| Dictyonella incisa | 22, 23 | VRE, B. subtilis |
| Dendrectilla tremitersis | 105–109 | S. aureus, MRSA, E. coli, P. aeruginosa |
| Dysidea sp. | 137, 138 | S. aureus, MRSA, Cryptoccus neoformans (C. neoformans), Microsporum gypseum (M. gypseum) |
| 190 | Chlamydia trachamatis (C. trachamatis) | |
| Epipolasis sp. | 163 | S. aureus, E. faecalis, MRSA, VRE E. faecium |
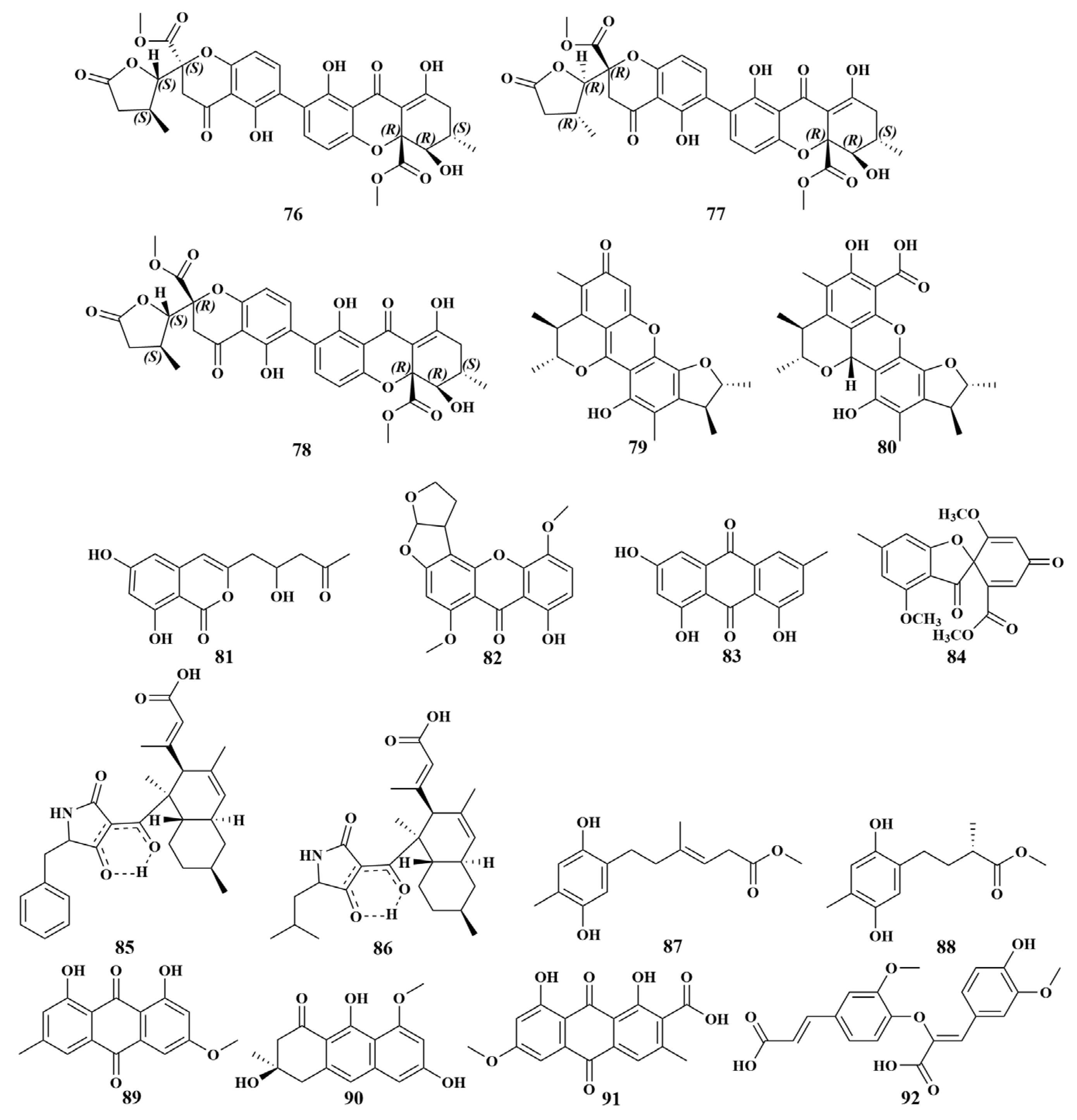
| Gelliodes carnosa | 76, 77, 78 | S. epidermidis, S. aureus, B. subtilis |
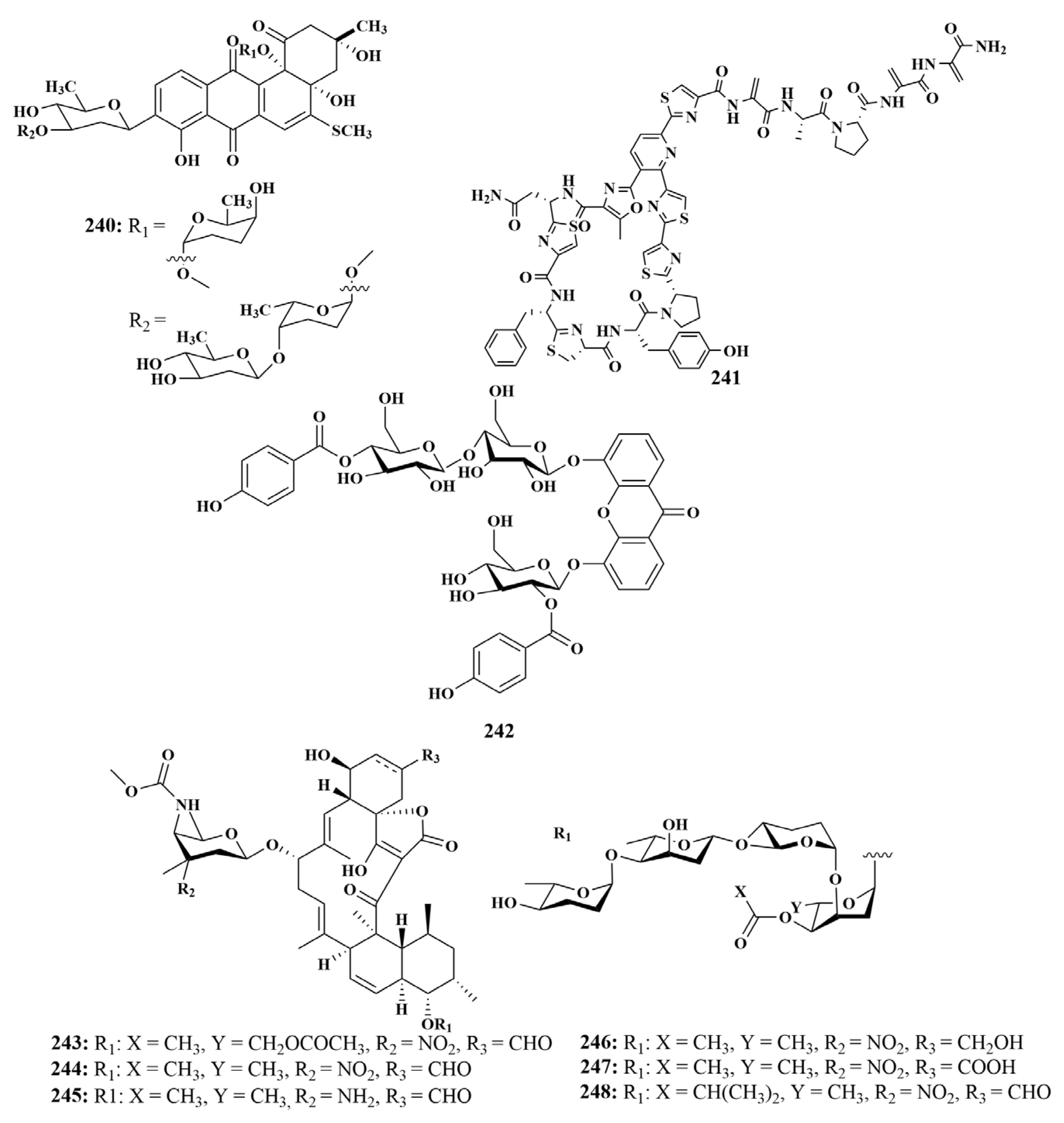
| 191, 221, 222, 227, 243–248 | B. subtilis | |
| 228 | S. epidermidis, S. aureus, E. faecium, E. faecalis | |
| Grantia compressa | 28, 29, 89, 90, 101, 167 | S. aureus, E. faecalis, E. coli, P. aeruginosa, Bacillus cereus (B. cereus), Micrococcus luteus (M. luteus), Salmonella typhimurium (S. typhimurium), Mycobacterium tuberculosis (M. tuberculosis), Klebsiella pneumoniae (K. pneumoniae), Streptococcus pneumoniae (S. pneumoniae) |
| Halichondria panicea | 198–202 | E. faecalis, S. aureus, E. coli, B. cereus, P. aeruginosa, Salmonella enterica (S. enterica), C. albicans, M. tuberculosis |
| 203, 204 | E. faecalis, S. aureus, E. coli, B. cereus, P. aeruginosa, S. enterica, C. albicans | |
| Haliclona sp. | 25, 26, 27 | Vibrio harvey (V. harvey) |
| 110, 111 | S. aureus, E. coli | |
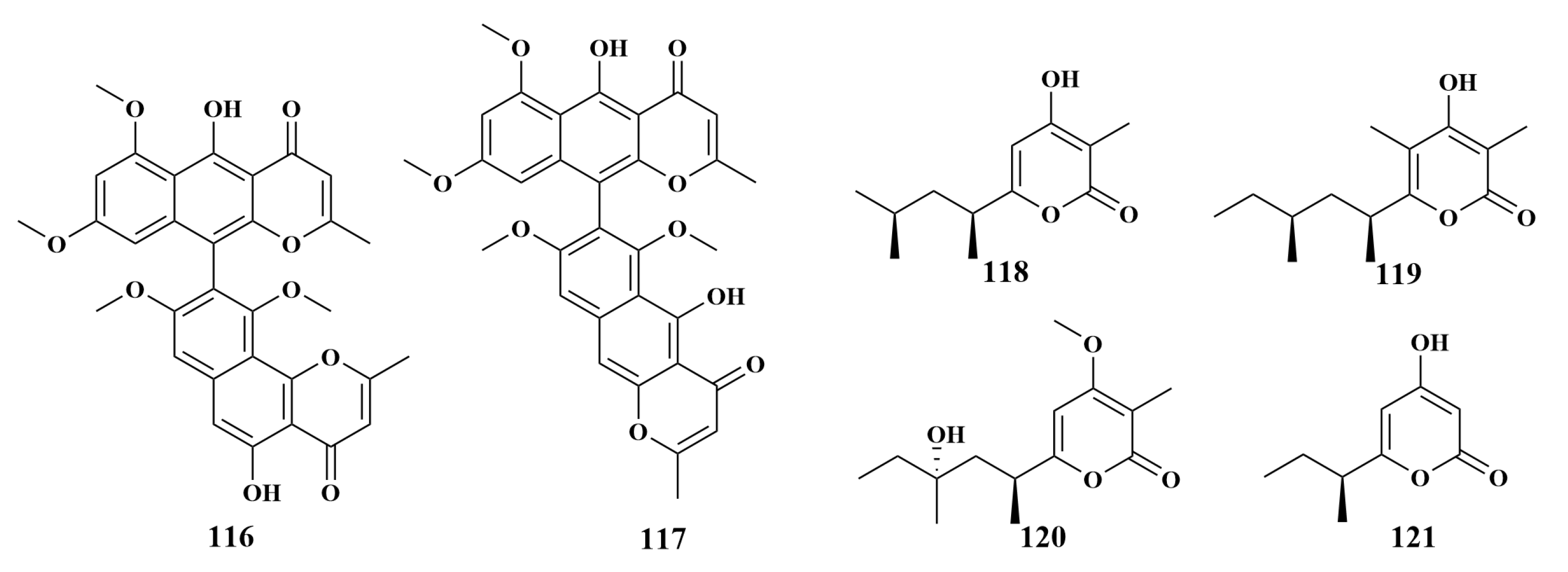
| 118, 119, 120, 121 | S. aureus, E. coli, B. subtilis, MRSA, M. tuberculosis | |
| 112, 113 | C. albicans, C. neoformans | |
| 114 | M. tuberculosis, S. aureus, E. coli, B. subtilis | |
| 164, 165 | B. subtilis | |
| Hymeniacidon perleve | 82 | S. aureus, B. subtilis |
| 83, 84 | MRSA, S. aureus, B. subtilis, Bacille Calmette Guerin (BCG) | |
| Hyrtios erectus | 81, 156, 157, 158 | S. aureus, E. coli, C. albicans |
| Isodictya setifera | 252 | S. aureus, P. aeruginosa |
| Lissodendoryx stigmata | 258 | MRSA, methicillin-sensitive Staphycoccus aureus (MSSA) |
| Melophlus sp. | 42 | MRSA, S. aureus, VRE E. faecium, C. neoformans |
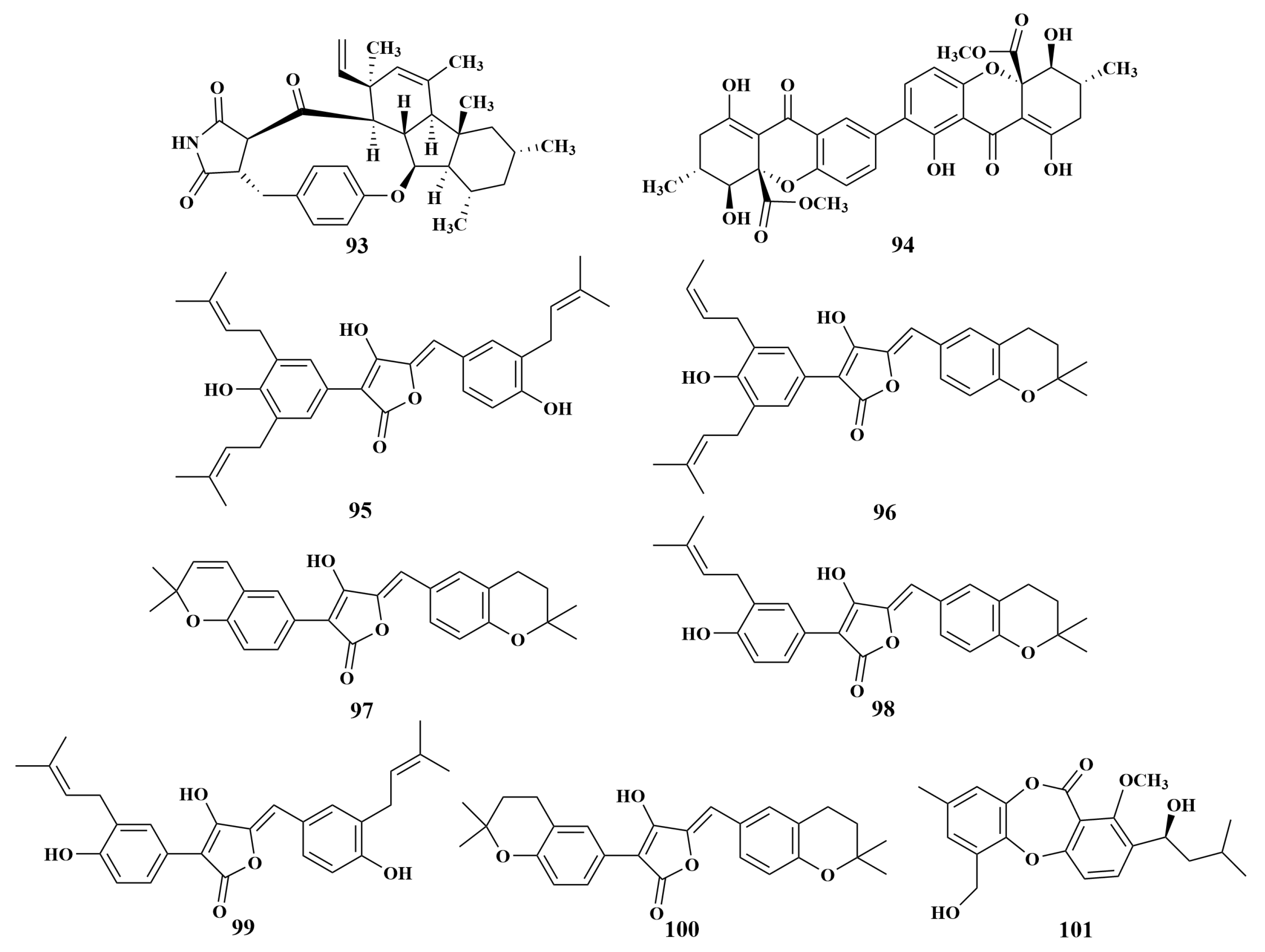
| Mycale sp. | 95–100 | S. aureus, E. faecalis, VRE E. faecalis, MRSA |
| 101 | E. faecalis | |
| Neopetrosia sp. | 93, 94 | VRE E. faecalis, S. aureus, E. faecalis, MRSA |
| Niphates sp. | 51, 52 | Pseudomonas lachrymans (P. lachrymans), Agrobacterium tumefaciens (A. tumefaciens), Xanthomonas vesicatoria (X. vesicatoria), Ralstonia solanacearum (R. solanacearum), S. aureus |
| 32, 33, 34, 35 | S. aureus | |
| Phakellia fusca | 24, 180, 181 | S. aureus |
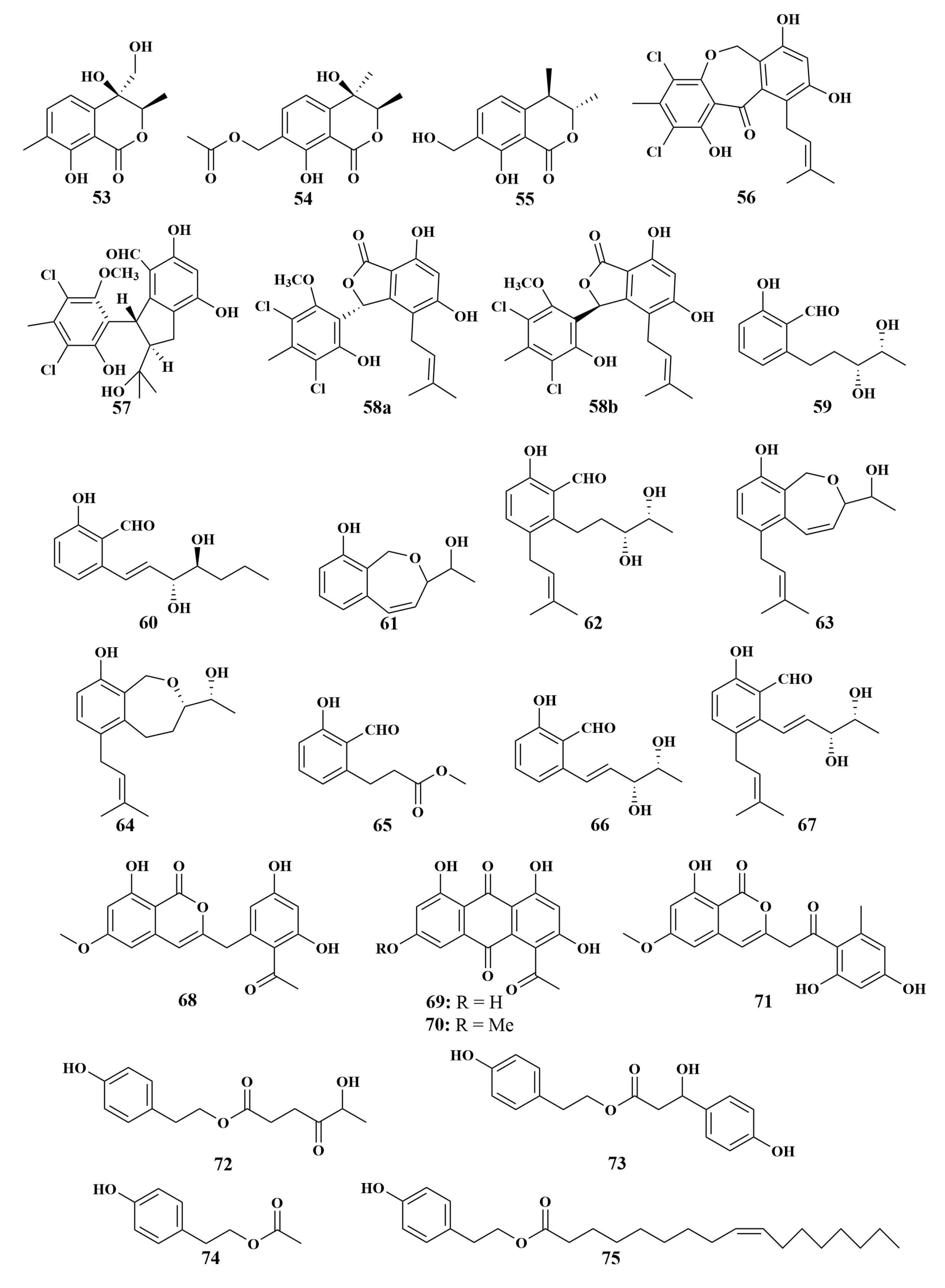
| 53, 54, 55, 56, 57, 58a/b | S. aureus, B. subtilis, C. albicans, C. neoformans, Candida parapsilosis (C. parapsilosis) | |
| 59–67 | S. aureus, B. subtilis, C. neoformans, C. parapsilosis | |
| 72, 73, 74, 75 | S. aureus, MRSA, E. coli | |
| 176, 177, 178, 179 | Aspergillus fumigatus (A. fumigatus), Aspergillus niger (A. niger) | |
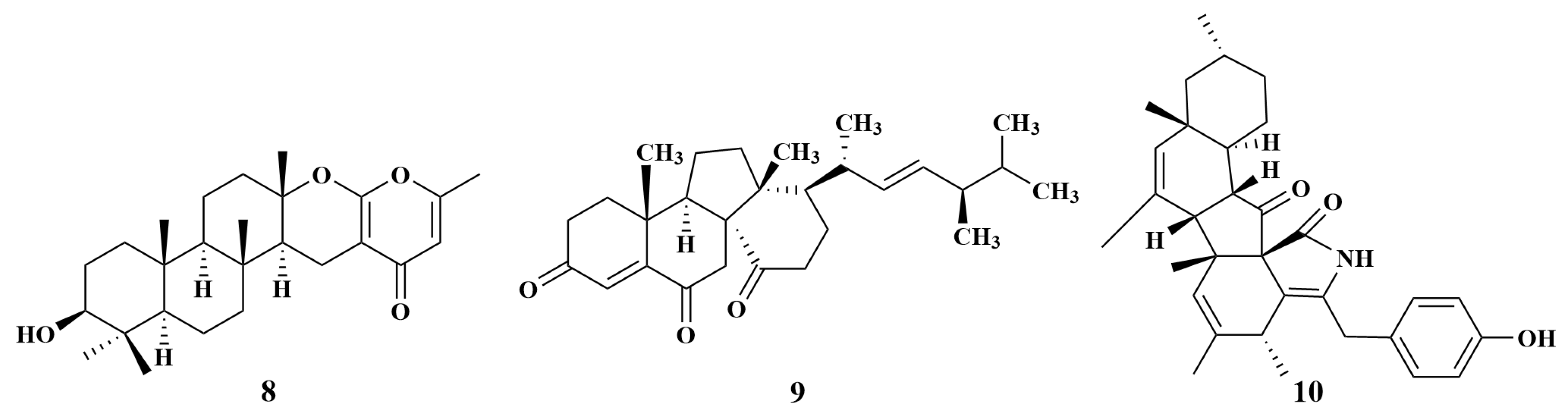
| Phomopsis sp. | 10 | S. aureus, MRSA |
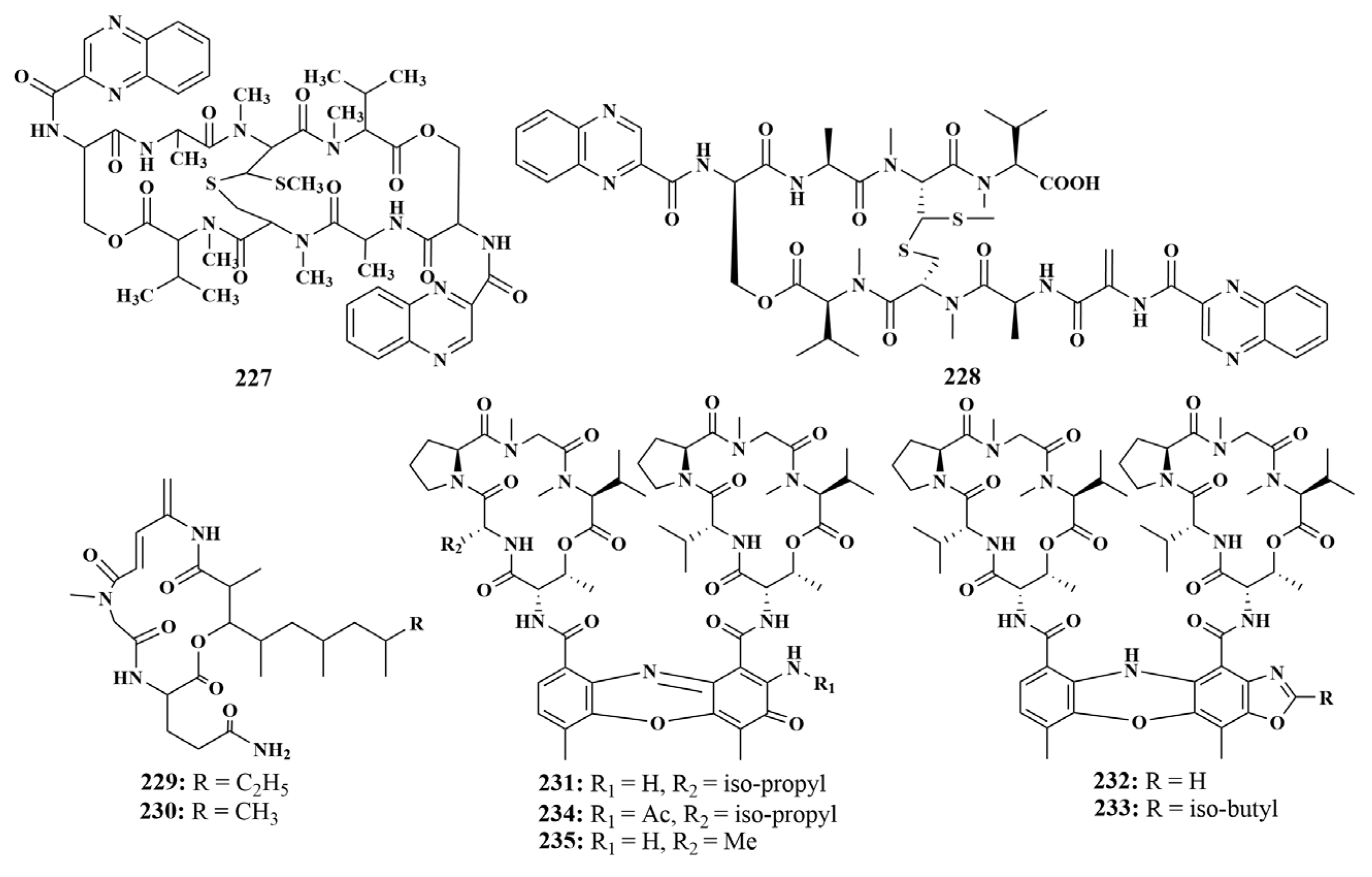
| Phyllospongia foliascens | 231–235 | MRSA |
| Reniera japonica | 116, 117 | Helicobacter pylori (H. pylori) |
| Rhabdermia sp. | 8 | MRSA |
| Scopalina ruetzleri | 189 | B. subtilis, Mycolicibacterium smegmatis (M. smegmatis) |
| Spheciosponge vagabunda | 242 | E. faecalis, S. aureus |
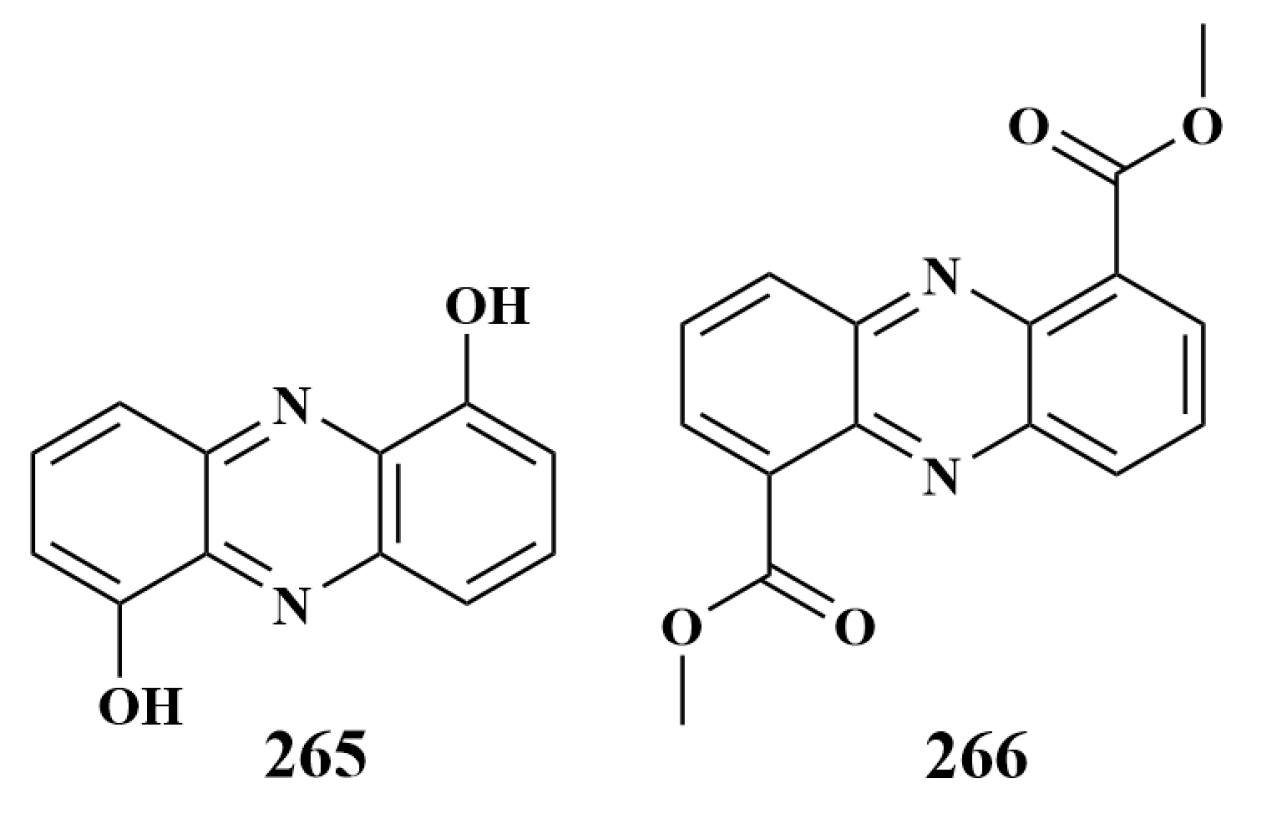
| 265 | B. subtilis | |
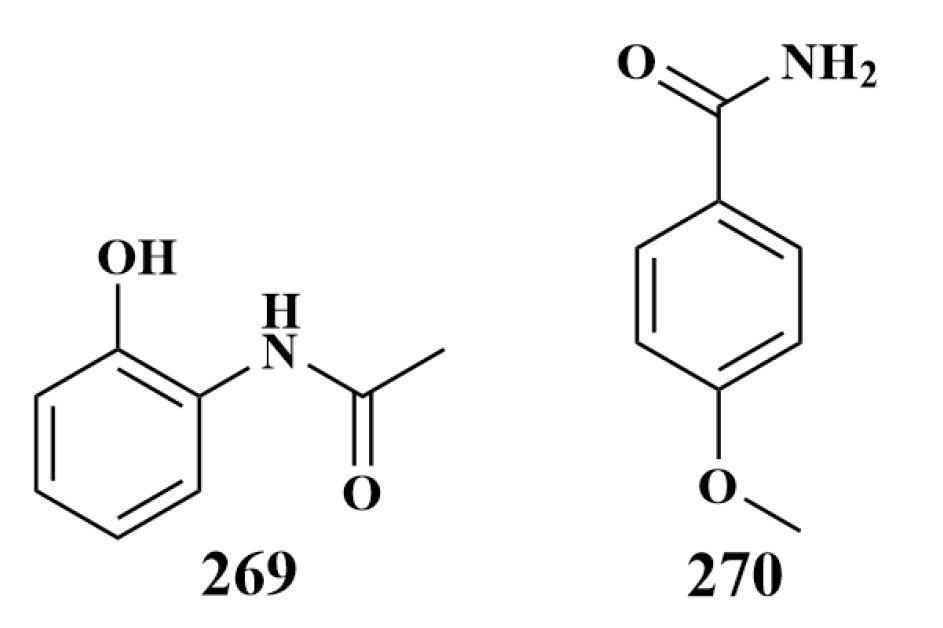
| 266, 269, 270 | S. aureus, B. subtilis, P. aeruginosa, E. coli | |
| Spongia officinalis | 253 | E. coli, S. aureus |
| Stelletta sp. | 139, 140, 141, 142 | S. aureus, Streptococcus iniae (S. iniae), Vibrio ichthyoenteri (V. ichthyoenteri) |
| Suberea sp. | 79, 80 | S. aureus, B. subtilis, Bacillus megaterium (B. megaterium), M. smegmatis |
| 148, 149 | C. albicans, S. aureus, E. coli | |
| Tedania sp. | 255, 256, 257 | M. luteus |
| Tethya aurantium | 45, 46, 47 | C. albicans, Septoria tritici (S. tritici), Trichophyton rubrum (T. rubrum), B. subtilis, Staphylococcus lentus (S. lentus), MRSA |
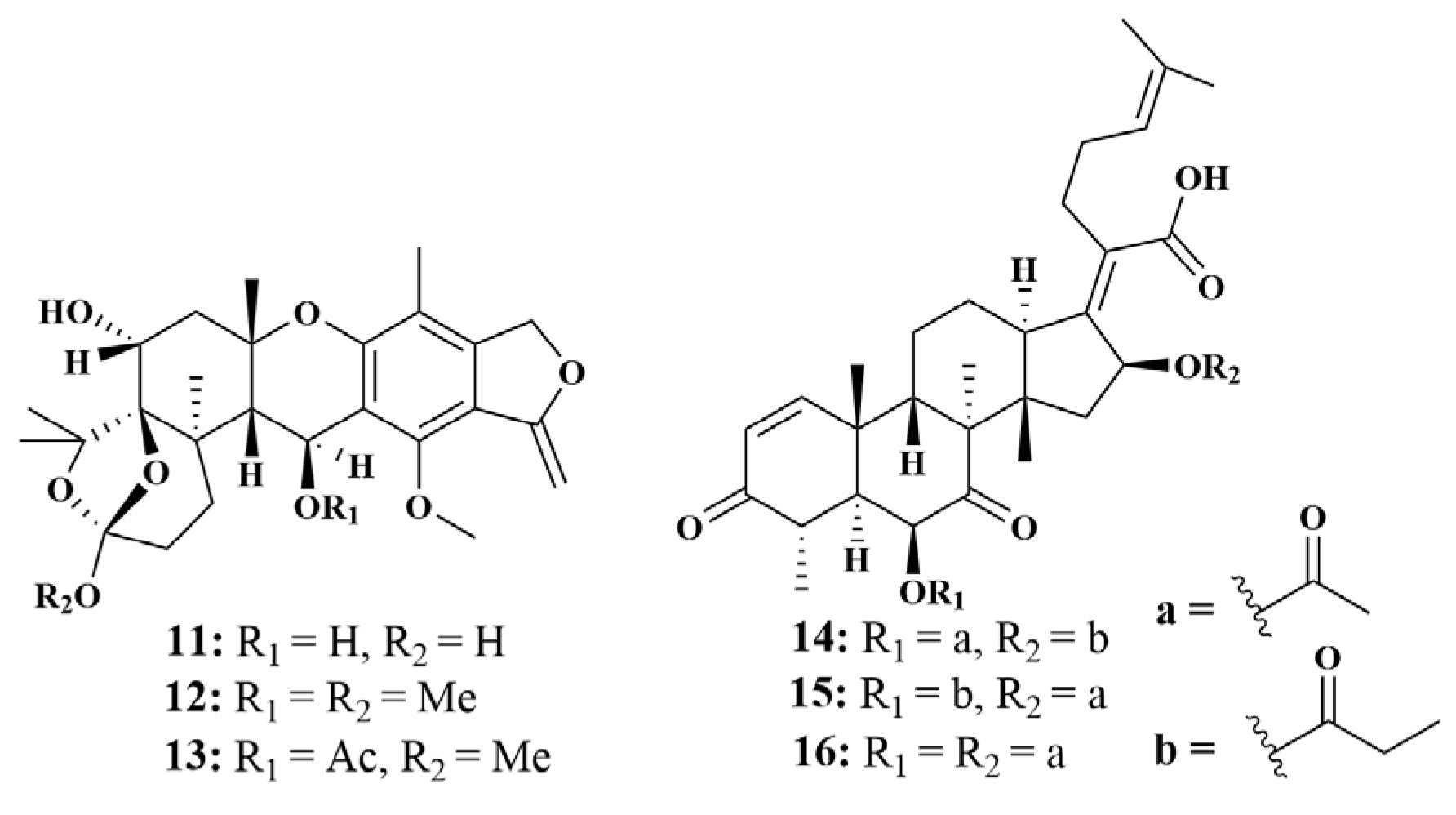
| 11, 12, 13, 92, 159, 160, 161 | S. aureus, Halomonas aquamarina (H. aquamarina), Polaribacter irgensii (P. irgensii), Pseudoalteromonas elyakovii (P. elyakovii), Roseobacter litoralis (R. litoralis), Shewanella putrefaciens (S. putrefaciens), Vibrio natriegens (V. natriegens), V. harvey, Vibrio carchariae (V. carchariae), V. proteolyticus | |
| Theonella sp. | 205 | MRSA, wild-type Staphylococcus aureus (WTSA), VRE E. faecium |
| 206, 207, 208 | MRSA | |
| Xestospongia sp. | 1, 2, 3, 4, 5 | S. aureus, B. subtilis, E. coli, B. cereus, Sarcina lutea (S. lutea), Micrococcus tetragenus (M. tetragenus), Vibrio anguillarum (V. anguillarum), V.parahaemolyticus |
| 6, 7, 68–71 | S. enterica | |
| 186, 187, 188, 240 | M. tuberculosis | |
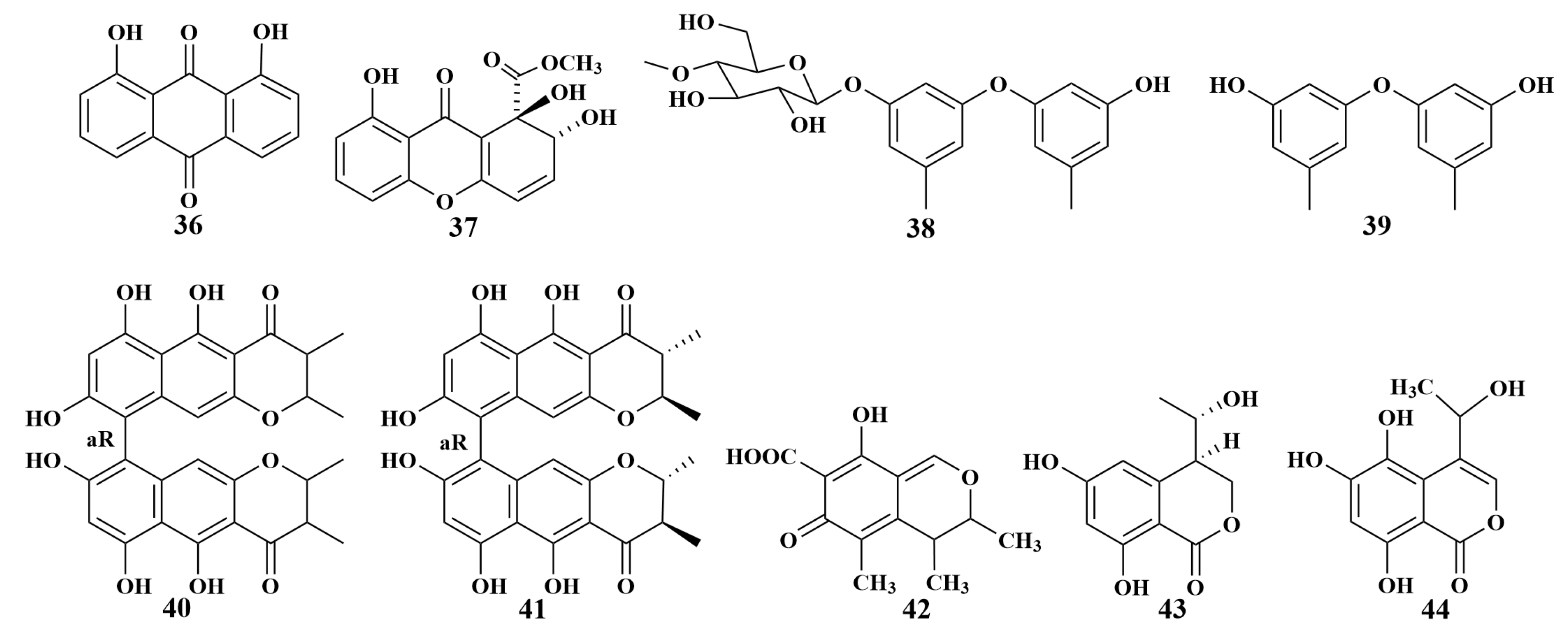
| Unidentified | 36, 37 | C. albicans |
| 38, 39, 40, 41 | Mycobacterium phlei (M. phlei), M. tuberculosis | |
| 43, 44 | B. cereus, V. parahaemolyticus, Streptomyces albus (S. albus) | |
| 144, 145 | C. albicans, C. neoformans, Candida glabrata (C. glabrata) | |
| 150, 151, 152, 153 | V. harvey | |
| 20, 21 | K. pneumoniae, E. faecalis, A. hydrophila | |
| 85, 86 | B. subtilis, S. aureus, MRSA, C. albicans, Propionibacterium acnes (P. acnes), S. epidermidis, Xanthomonas campestris (X. campestris), Alternaria alternata (A. alternata) | |
| 169, 170 | Alternaria brassicae (A. brassicae) | |
| 154, 155 | S. aureus, Aeromonas hydrophila (A. hydrophila), V. harvey, Gaeumannomyces gramini (G. gramini), V. parahaemolyticus | |
| 115 | S. aureus, M. tuberculosis | |
| 14, 15, 16 | Streptococcus agalactiae (S. agalactiae) | |
| 166 | V. parahaemolyticus | |
| 87, 88 | P. aeruginosa, M. phlei, meticillin-resistant coagulase negative Staphylococcus (MRCNS), B. subtilis, B. cereus, V. parahemolyticus | |
| 171, 172, 173, 174, 175 | B. cereus, Proteus sp., M. phlei, B. subtilis, MRSA, MRCNS, V. Parahaemolyticus, Edwardsiella tarda (E. tarda) | |
| 162 | S. epidermidis | |
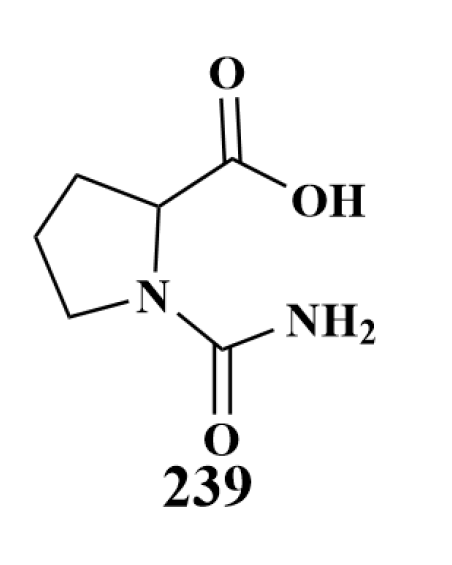
| 238, 239 | P. aeruginosa | |
| 241 | MRSA | |
| 225, 226 | M. luteus, Bacillus mycoides (B. mycoides), S. aureus, E. coli | |
| 237 | M. luteus | |
| 236 | E. coli | |
| 229, 230 | B. subtilis, E. coli | |
| 209 | H. pylori, P. aeroginosa, A. baumanniiin, E. coli, K. pneumonia, S. aureus, C. albicans, E. faecium | |
| 210 | H. pylori, K. pneumonia, S. aureus, E. faecium | |
| 211, 212 | S. agalactiae, B. subtilis, E. coli, S. aureus | |
| 192, 193 | MRSA, E. coli | |
| 213 | K. pneumonia, S. aureus, A. baumannii | |
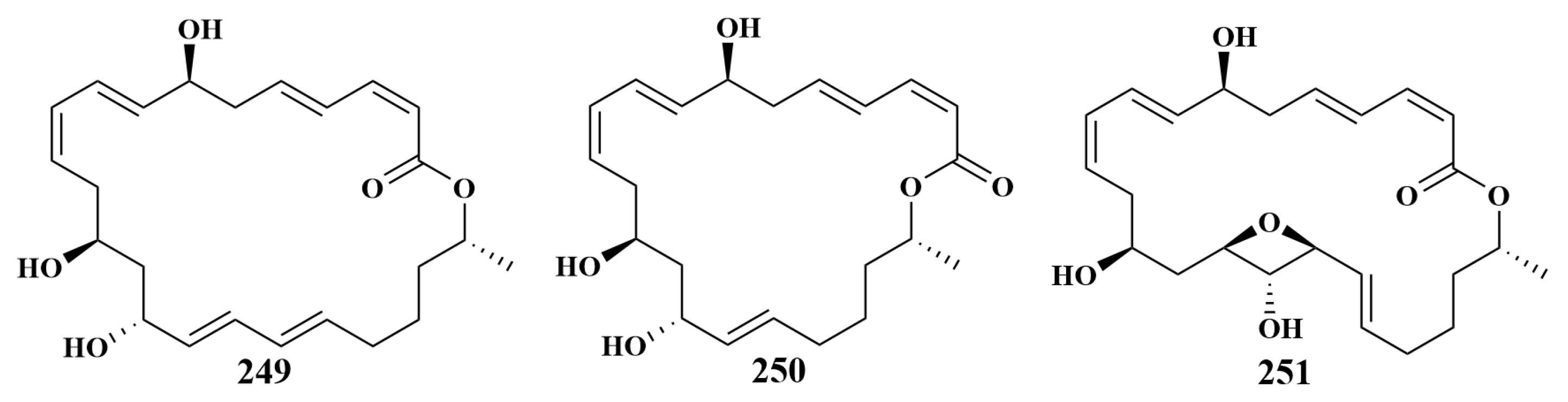
| 249, 250, 251 | P. aeruginosa, S. aureus, Rhodococcus baikonurensis (R. baikonurensis), E. coli | |
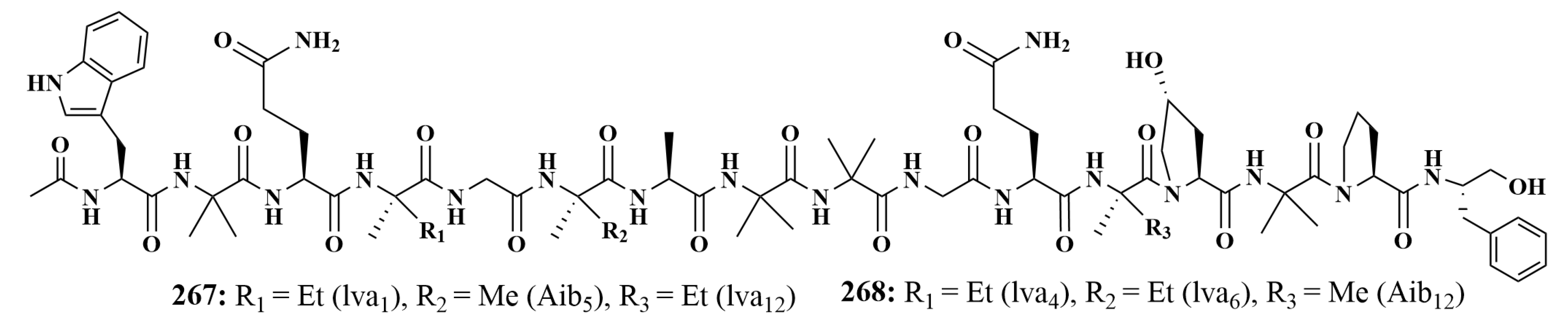
| 267, 268, 263, 264 | B. subtilis, S. aureus, MRSA, C. albicans |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, J.; She, J.; Fu, J.; Wang, J.; Ye, Y.; Yang, B.; Liu, Y.; Zhou, X.; Tao, H. Advances in Natural Products from the Marine-Sponge-Associated Microorganisms with Antimicrobial Activity in the Last Decade. Mar. Drugs 2023, 21, 236. https://doi.org/10.3390/md21040236
Liang J, She J, Fu J, Wang J, Ye Y, Yang B, Liu Y, Zhou X, Tao H. Advances in Natural Products from the Marine-Sponge-Associated Microorganisms with Antimicrobial Activity in the Last Decade. Marine Drugs. 2023; 21(4):236. https://doi.org/10.3390/md21040236
Chicago/Turabian StyleLiang, Jiaqi, Jianglian She, Jun Fu, Jiamin Wang, Yuxiu Ye, Bin Yang, Yonghong Liu, Xuefeng Zhou, and Huaming Tao. 2023. "Advances in Natural Products from the Marine-Sponge-Associated Microorganisms with Antimicrobial Activity in the Last Decade" Marine Drugs 21, no. 4: 236. https://doi.org/10.3390/md21040236
APA StyleLiang, J., She, J., Fu, J., Wang, J., Ye, Y., Yang, B., Liu, Y., Zhou, X., & Tao, H. (2023). Advances in Natural Products from the Marine-Sponge-Associated Microorganisms with Antimicrobial Activity in the Last Decade. Marine Drugs, 21(4), 236. https://doi.org/10.3390/md21040236