
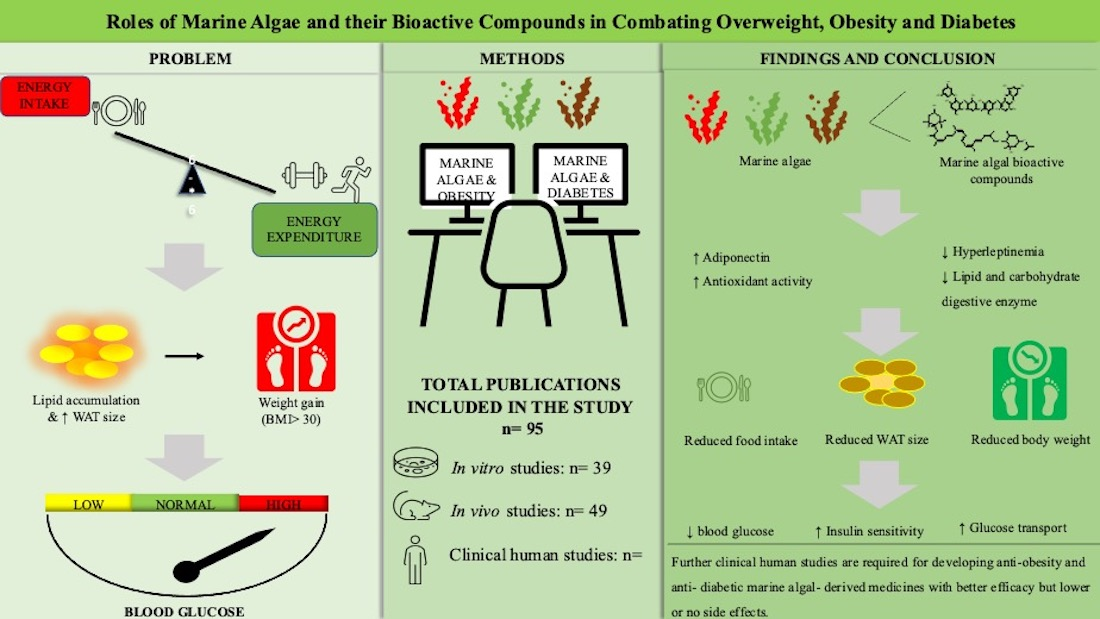
Roles of Marine Macroalgae or Seaweeds and Their Bioactive Compounds in Combating Overweight, Obesity and Diabetes: A Comprehensive Review



Abstract
:
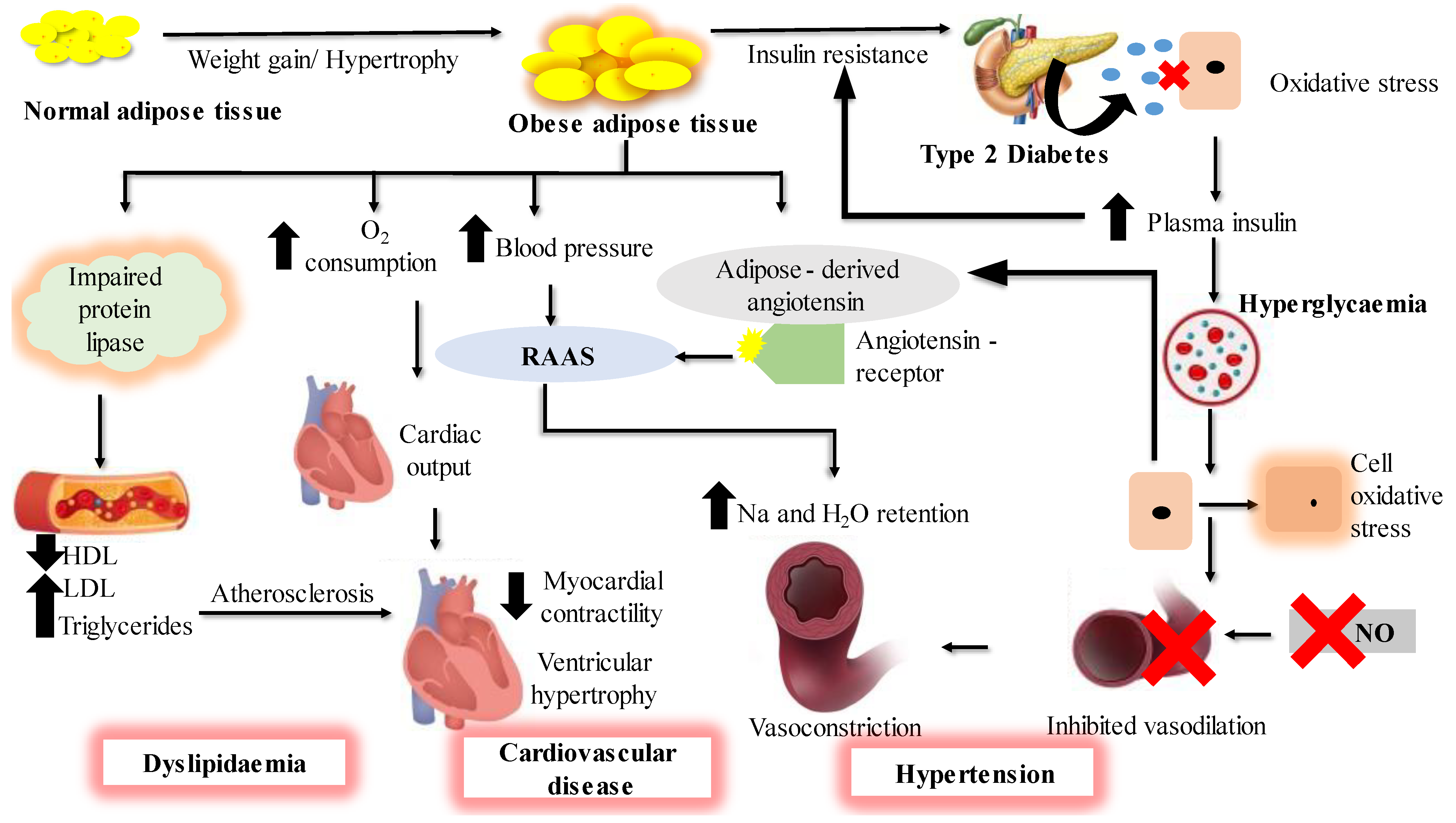
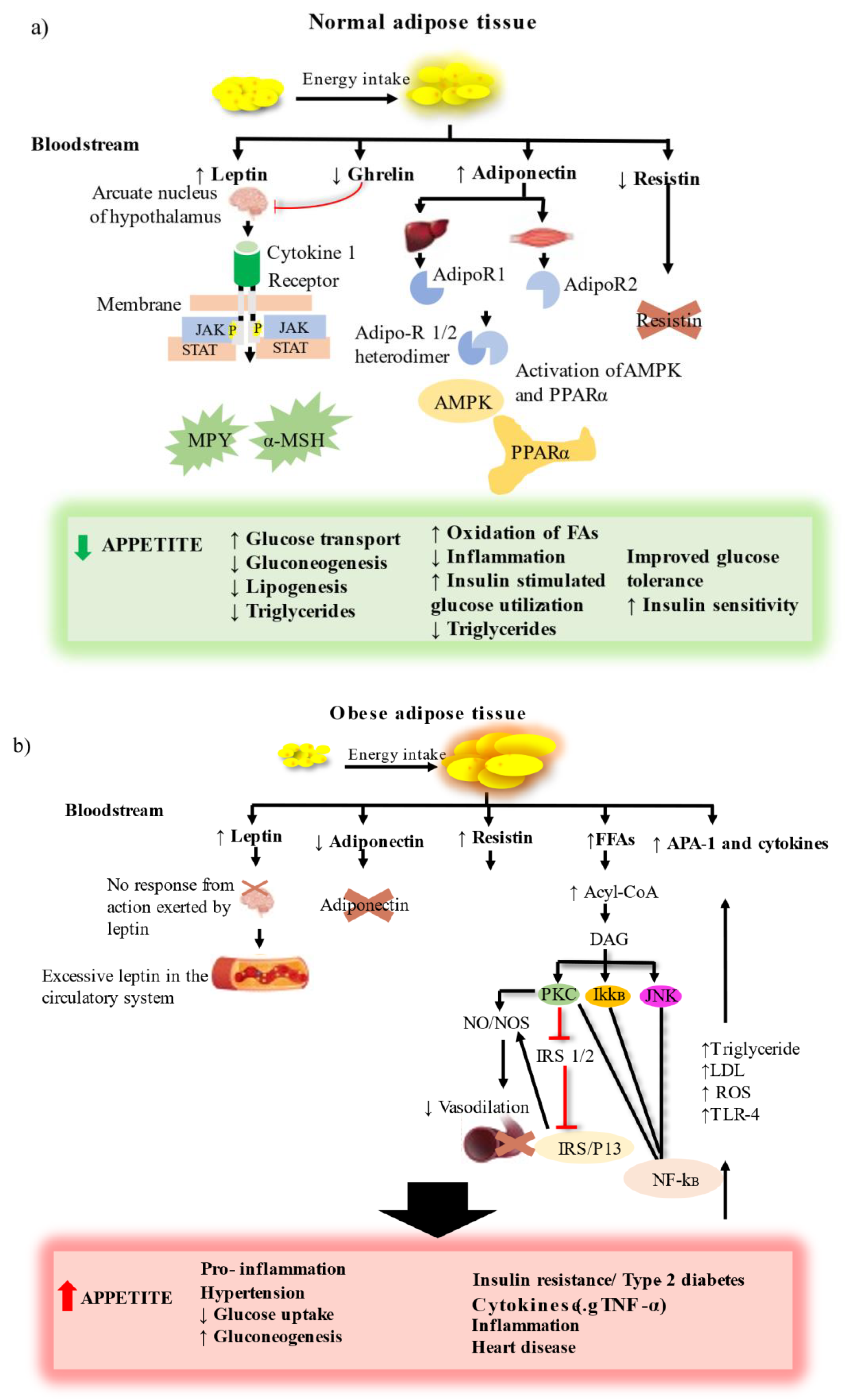
1. Introduction
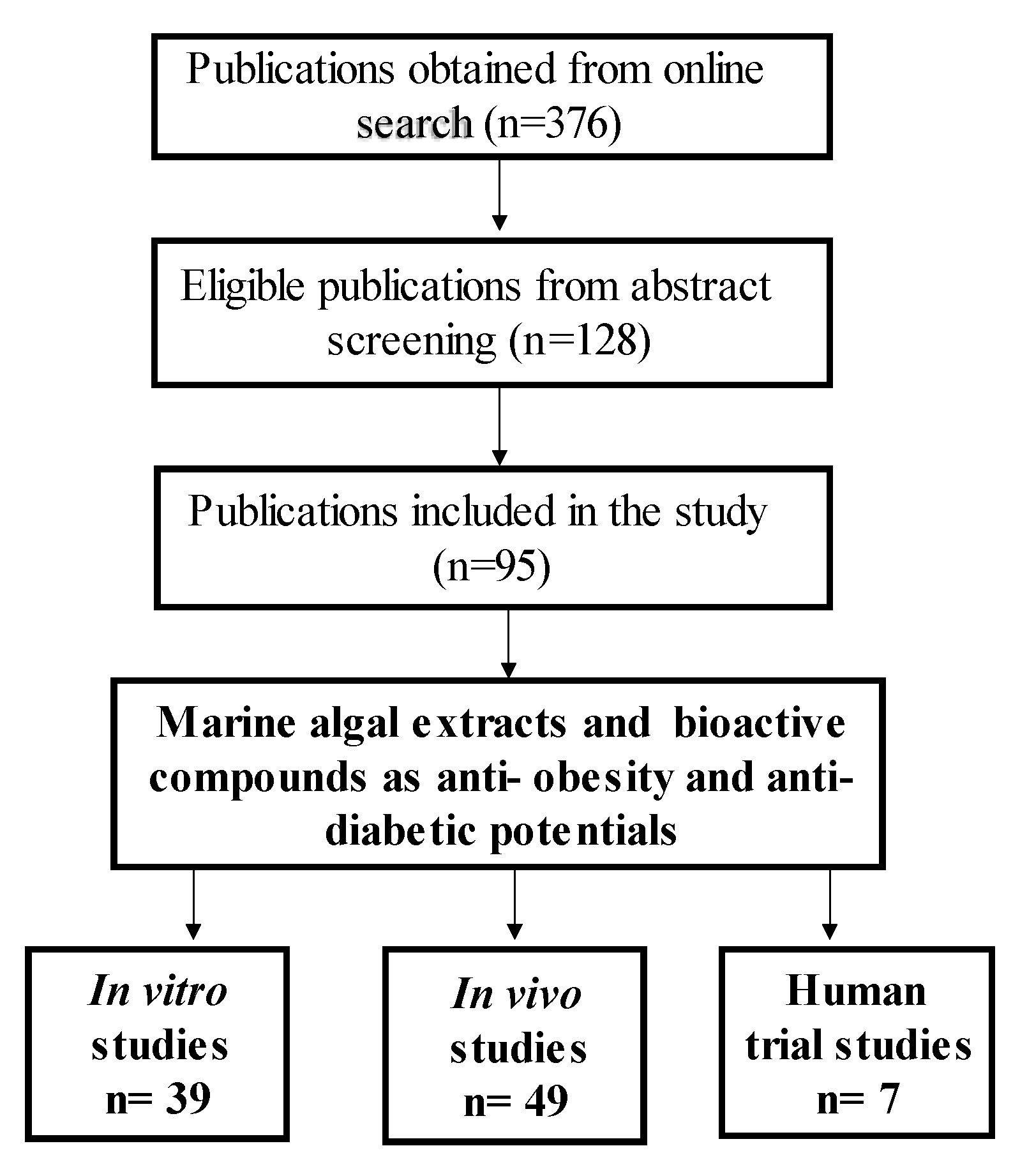
2. Primary Marine Algal Bioactive Compounds to Treat Obesity and T2D
2.1. In Vitro Studies
2.2. In Vivo Studies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Compounds | Models | Biological Effects | References |
|---|---|---|---|
| Fucoxanthin | 3T3-L1 cells | ↓ Intracellular lipid accumulation | [25] |
| Inhibition of α- amylase | |||
| Weak inhibition of α- glucosidase | |||
| ↓ Lipid accumulation | |||
| ↓ Glycerol-3-phosphate dehydrogenase activity | |||
| ↓ PPARγ regulation | |||
| Fucoxanthinol | 3T3-L1 adipocyte cells and a RAW264.7 macrophage | ↓ TNF- α and MCP- 1 mRNA expression | [26] |
| ↓ protein levels and macrophage cells | |||
| Siphonaxanthin | 3T3-L1 preadipocytes | ↓ Adipogenesis | [34] |
| ↓ Adipocyte differentiation | |||
| ↓ Protein kinase b phosphorylation | |||
| ↓ Gene expression of C/EBPα, PPARγ, FABP4 and SCD1 | |||
| 3T3-L1 adipocytes | ↓ Lipid accumulation | [43] | |
| HepG2 cell line | ↓ Triglyceride accumulation | [35] | |
| ↓ Fatty acid translocase and FABP4 expression | |||
| ↓ Hepatic lipogenesis | |||
| Astaxanthin | 3T3-L1 adipocytes | ↓ Lipid accumulation | [39] |
| ↓ Lipogenesis genes | |||
| Kinetics analysis | Inhibition of pancreatic lipase | [40] | |
| Inhibition of α- glucosidase | [41] | ||
| Neoxanthin | 3T3-L1 adipocyte | ↓ Glycerol-3- dehydrogenase | [43] |
| ↓ Expression of C/EBPα and PPARγ mRNA | |||
| ↓ Adipogenesis | |||
| ↓ Lipid accumulation | [44] | ||
| Fucoidan | 3T3-L1 cells | ↓ Expression of AAP2, ACC and PPARγ gene | [46,47] |
| ↓ Reactive oxygen species (ROS) | |||
| ↓ Lipid accumulation | |||
| ↑ Lipolysis | |||
| ↓ Adipogenesis | [30] | ||
| In vitro without cell line | Inhibition of pancreatic lipase | [31,49,50] | |
| Inhibition of α- amylase | |||
| Inhibition of α- glucosidase | |||
| Phlorotannins | 3T3-L1 preadipocytes | ↓ Lipid accumulation | [30,31] |
| ↓ Expression of C/EBPα and PPARγ Mrna | |||
| ↓ Expression of SREBP1 and FABP4 | |||
| MAPK activation | |||
| Carbohydrate digesting enzyme and tyrosine phosphate 1B inhibition | Inhibition of α- glucosidase | [32] | |
| Inhibition of tyrosine phosphate 1B | |||
| Alginates | Lipid digestive enzyme inhibition, antioxidant activity | Inhibition of pancreatic lipase | [55] |
| ↓ Reactive oxygen species (ROS) | [56] |
2.3. Clinical Studies
| Algal Compounds | Dose/Concentration and Route of Administration | Model | Biological Effects | References |
|---|---|---|---|---|
| Fucoxanthin | Standard dose: 1 mg/kg, intragastric administration | HFD-induced obesity | ↓ WAT mass, serum triacylglycerol | [57] |
| ↓ Expression of leptin and IL-6 | ||||
| Upregulation of UCP- 1 expression in BAT, PPARα, PGC1α and PPARγ | ||||
| ↑ β- Oxidation, energy expenditure | ||||
| ↓ Serum level of adiponectin and leptin expression | ||||
| Improved insulin, reduced blood pressure | ||||
| ↓ Adiponectin concentration | ||||
| Low dose 1.06% and high dose 2.22%, dietary supplement | HFD-induced obesity | ↓ Weight gain and WAT size | [58] | |
| ↓ Hyperglycemia | ||||
| ↓ Hyperinsulinemia | ||||
| ↓ Hyperleptinemia | ||||
| ↓ Adipose tissue size and weight gain | ||||
| ↑ UCP-1 expression | ||||
| ↑ GLUT- 4 mRNA expression | ||||
| ↑ Adrβ3 mRNA expression | ||||
| 0.2% fucoxanthin, dietary supplement | Diabetic/obese KK-Ay mice | Improved insulin, blood glucose and leptin | [59] | |
| ↓ MCP- 1 mRNA expression | ||||
| ↑ mRNA expression of Adrβ3 | ||||
| ↓ Inflammatory markers | ||||
| ↓ Lipid accumulation | ||||
| ↓ Decreased glycerol-3-phosphate dehydrogenase activity (lipid differentiation) | ||||
| Upregulation of UCP- 1 expression in BAT ↓ PPARα, PGC1α and PPARγ | ||||
| Doses: 0.2–0.6%, intragastric administration | HFD-induced obesity | ↓ Weight gain | [60] | |
| ↓ Mammary gland inflammation | ||||
| ↓ MDA levels | ||||
| ↓ Myeloperoxidase (MPO) | ||||
| ↓ Production of IL-1β, TNF-α, iNOS and COX-2 | ||||
| 0.1%, dietary supplement | Diabetic model KK-Ay | Improved glucose tolerance | [75] | |
| ↓ Proinflammation | ||||
| Low dose 0.2% and high dose 0.4%, dietary supplement | C57BL/KsJ- db/db mice | Improved insulin resistance | [26,61] | |
| Improved lipid metabolism | ||||
| ↑ Regulation IRS-1/PI3K/AKT and AMPK | ||||
| ↓ Plasma lipid levels | ||||
| Standard dose: 0.2%, dietary supplement | Diabetic/obese KK-Ay mice | ↑HDL and non-HDL cholesterol | [80] | |
| ↑ SREBP1, SREBP2 | ||||
| ↓ Food intake | ||||
| ↓ Epididymal WAT gain | ||||
| Standard dose: 0.2%, dietary supplement | Diabetic/obese KK-Ay mice | ↓ Hyperglycemia | [81] | |
| ↓ Hyperinsulinemia | ||||
| ↑ GLUT- 4 mRNA expression | ||||
| Standard dose: 400 mg/kg, intragastric administration | STZ-induced diabetes mice | ↓ Hyperinsulinemia | [82] | |
| ↓ Plasma triglyceride | ||||
| ↓ LDL cholesterol | ||||
| ↑ Regeneration of pancreatic β cells | ||||
| Astaxanthin | Low dose 1.2 mg/kg, medium dose 6 mg/kg and high dose 30 mg/kg, dietary supplementation | ddY mice | ↓ Body and weight, and WAT size | [65] |
| ↓ Liver and plasma triglyceride and total cholesterol | ||||
| Standard dose: 0.02%, dietary supplementation | ICR mice | ↑ Fat utilization during exercise | [66] | |
| ↑ CPT- 1 activation | ||||
| ↓ Fat accumulation | ||||
| Standard dose: 6 mg/kg bw, oral administration | High fat and high fructose diet (HFFD)-fed model | ↑ Insulin sensitivity | [67] | |
| ↓ Liver damage by ↓ CYP2E1, myeloperoxidase, nitro-oxide stress | ||||
| Improved antioxidant activity | ||||
| ↓ Lipid deposition | ||||
| ↓ TGF-β1 expression | ||||
| Standard dose: 30 mg/kg bw, form of administration not indicated | Pregnant C57BL/KsJ db/+ | ↑ Antioxidant activity | [68] | |
| Improved glucose tolerance | ||||
| Improved reproductive outcomes | ||||
| ↑ Regeneration of pancreatic β cells | ||||
| Low dose10 mg/kg bw, medium dose 25 mg/kg bw, high dose 40 mg/kg bw, oral administration | C57BL/KsJ+/+ (wild-type) and C57 BL/KsJdb/+(db/+) | Improved insulin sensitivity | [83] | |
| Improved glucose tolerance | ||||
| Improved antioxidant activity | ||||
| Improved reproductive outcomes | ||||
| ↑ GLUT- 4 mRNA expression | ||||
| ↓ Inflammation | ||||
| Fucoidan | 20 mg/kg bw, dietary supplementation | Male ICR mice | ↓ Fasting blood glucose | [64] |
| Restored phosphorylation of Akt | ||||
| ↓ Malondialdehyde (MDA) | ||||
| Activated Nrf2 pathway | ||||
| ↑ GSH/GSSG ratio | ||||
| ↑ Antioxidant enzymes | ||||
| Low dose 1.5 and high dose 2.0%, dietary supplement | C57BL/6 mice | ↓ Triglycerides, total cholesterol and LDL proteins | [84] | |
| ↓ Body weight, liver and epididymal fats | ||||
| ↓ Food efficiency ration | ||||
| Standard dose: 45 mg/kg bw, oral administration | C57BL/KSJ db/db mice | ↓ Hyperglycemia | [71] | |
| ↓ Blood glucose levels | ||||
| Alginates | Standard dose: 50 mg/kg, oral gavage | HFD-induced obese mice | ↓ Fat, cholesterol and triglyceride accumulation | [72] |
| Alteration in colonic genome for immune regulation | ||||
| ↓ Blood glucose levels | ||||
| Standard dose: 1 mL/100 mg bw, oral gavage | HFD-induced obese mice | Alteration in colonic genome for immune regulation | [73] | |
| ↓ Inflammatory bacteria | ||||
| Regulate gut microbiota | ||||
| ↓ Hyperlipidemia | ||||
| ↓ Hyperinsulinemia | ||||
| ↓ Blood glucose levels | ||||
| Standard dose: 50 mg/kg, macroencapsulation of islets | STZ-induced diabetes pigs | ↓ Blood glucose levels | [74] | |
| ↓ Diabetes for 6 months | ||||
| Phlorotannins | 60 mg/kg bw, oral gavage | STZ-induced diabetes rat | ↓ MDA levels | [62] |
| ↓ Blood glucose | ||||
| ↓ α-glucosidase and α-amylase activities | ||||
| ↑ Serum insulin | ||||
| ↑ Antioxidant activity | ||||
| ↑ Hepatic glutathione and AMPK-α | ||||
| Siphonaxanthin | Standard dose: 1.3 mg/kg bw, oral gavage | KK-Ay mice | ↓ Total weight of WAT | [34] |
| ↓ Lipogenesis | ||||
| ↑ Fatty acid oxidation in adipose tissues | ||||
| Standard dose: 0.016%, dietary supplement | C57BL/6JhamSlc-Ob/Ob | ↓ Plasma glucose and alanine transaminase (ALT) | [64] | |
| ↓ Lipid peroxidation | ||||
| ↑ Antioxidant signaling β-Oxidation |
3. Secondary Marine Algal Compounds to Treat Obesity and T2D
3.1. In Vitro Studies
3.2. In Vivo Studies
| Algal Compounds | Dose/Concentration and Route of Administration | Participants | Duration of the Study | Biological Effects | References |
|---|---|---|---|---|---|
| Astaxanthin | 5 mg and 20 mg, oral dose | n = 23 healthy men and women | 3 weeks | ↓ Lipid peroxidation | [75] |
| ↑ Antioxidant defense system | |||||
| Fucoidan | 500 mg, oral dose | n = 25 obese and overweight adults | 3 months | ↓ Diastolic blood pressure | [78] |
| ↓ LDL cholesterol | |||||
| ↑ Insulin secretion and sensitivity | |||||
| Alginates | 1.25% alginate chocolate milk, 2.5% alginate chocolate or 2.5% alginate solution, dietary supplementation | n = 24 healthy men | 1 day experiment | ↑ Weight loss when supplemented in diet | [76] |
| ↑ Satiety | |||||
| ↓ Energy intake | |||||
| Delayed gastric clearance | |||||
| ↑ Viscosity of digesta | |||||
| ↓ Nutrient absorption in small intestines | |||||
| 1.5 g, dietary supplement | n = 96 obese men and women | 12 weeks | ↑ Weight loss when supplemented in diet | [77] | |
| ↓ Body fat | |||||
| No changes in metabolic risk markers | |||||
| 1.5 g, supplemented drink | n = 68 healthy men and women | 4 weeks | ↓ Energy intake | [79] | |
| 1.5 g, supplemented drink | n = 14 healthy men | 1 day experiment | ↓ Cholesterol, triglycerides | [79] | |
| ↓ Blood glucose |
| Compounds | In Vitro Models | Biological Effects | References |
|---|---|---|---|
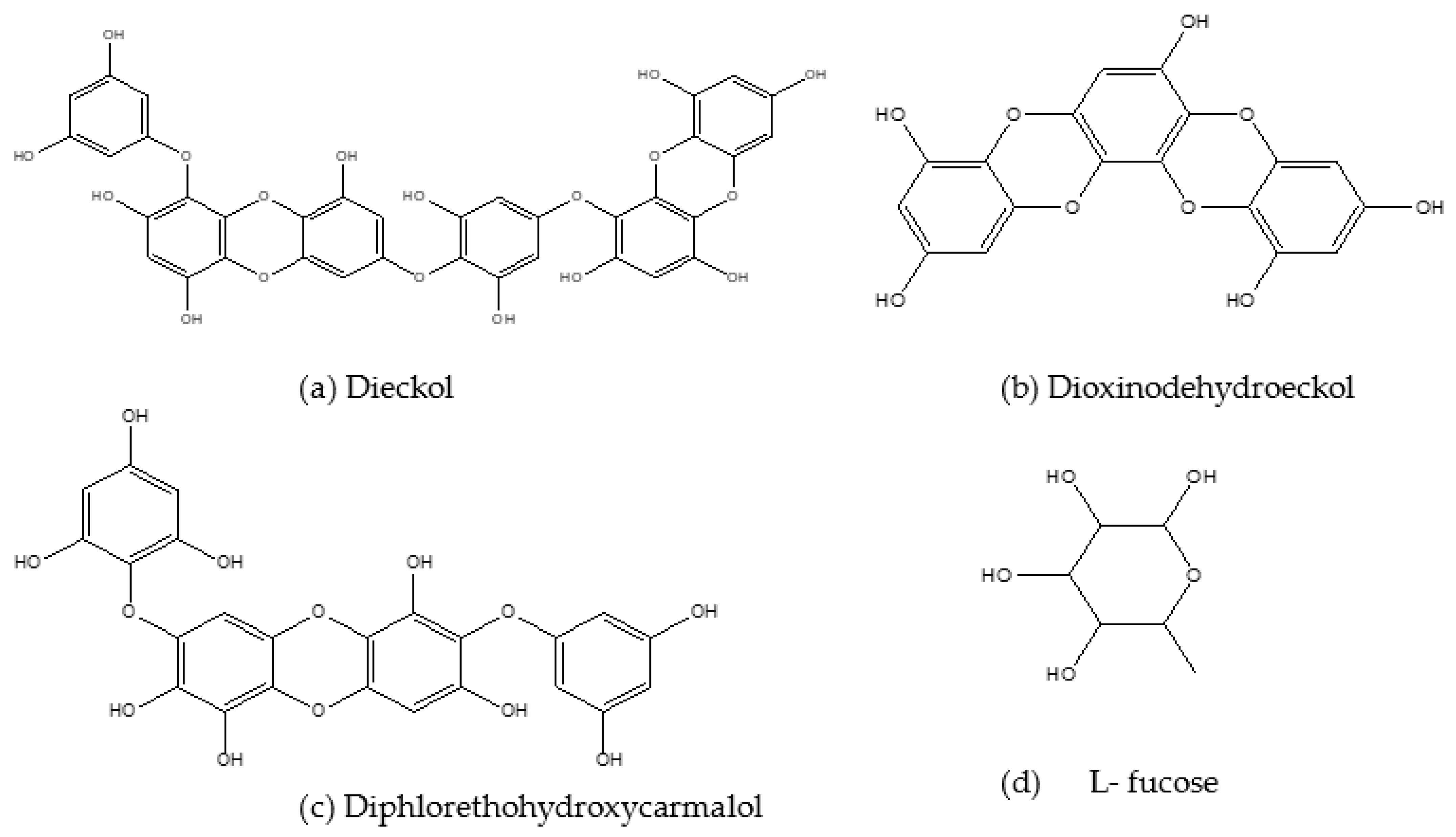
| Dieckol and seapolynol | 3T3 L1 preadipocytes | ↓ Lipid accumulation | [86] |
| ↓ 3- hydroxyl- methyl glutaryl coenzyme reductase | |||
| Dieckol | 3T3 L1 adipocytes | ↓ Adipogenesis | [31,93] |
| ↓ Adipogenesis gene and protein expression | |||
| ↑ AMPK pathway | |||
| ↓ Expression of C/EBPα, FABP4 and PPARγ mRNA | |||
| Lipid digestive enzyme inhibition | ↓ Pancreatic lipase | [87] | |
| Carbohydrate digestive enzyme inhibition | Inhibition of α- amylase | [92] | |
| Inhibition of α- glucosidase | |||
| Dioxinodehydroeckol (DHE) | 3T3 L1 adipocytes | ↓ Adipogenesis gene and protein expression | [69] |
| ↓ Lipid accumulation | |||
| ↑ AMPK pathway | |||
| ↓ Expression of C/EBPα, PPARγ mRNA, SREBP1, FATP1, FAS, LPL, ACS1 and FABP4 | |||
| Carbohydrate digestive enzyme inhibition | ↓ α- amylase activity and α- glucosidase activity | [89] | |
| Antioxidant activity | ↓ Antioxidant activities | [90] | |
| Indole derivatives STC-1 and STC- 5/Sargassum thunbergii | 3T3-L1 adipocytes | ↓ Expression of C/EBPα, SREBP1 and PPARγ mRNA | [91] |
| MAPK activation | |||
| Diphlorethohydroxycarmalol (DPHC) | Carbohydrate digestive enzyme inhibition | Inhibition of α- amylase | [103] |
| Inhibition of α- glucosidase |
3.3. Effective Marine Algae Extracts in the Management of Obesity and T2D
| Algal Compounds | Dose/Concentration and Route of Administration | Models | Biological Effects | References |
|---|---|---|---|---|
| Dieckol | Low dose 15 mg/kg and high dose 60 mg/kg, dietary supplement. | ICR mice | ↓ Lipid accumulation and adipogenesis gene expression | [93] |
| Standard dose: 100 mg/kg bw, oral gavage | ICR mice | ↓ Blood glucose | [92] | |
| Delayed carbohydrate absorption | ||||
| Dieckol and Seapolynol | Low dose 30 mg/kg and high dose 120 mg/kg bw, oral gavage | C57BL/KsJ-db/db mice | ↓ Body weight and water intake | [94] |
| ↓ Fasting blood glucose and serum insulin levels | ||||
| ↓ Cholesterol and triglyceride levels | ||||
| ↑ Serum adiponectin, glucose and lipid metabolism | ||||
| 12.5-5 mg seapolynol and 0.5- 2 mg dieckol, oral gavage | Male ICR mice | ↓ Body weight gain | [86] | |
| ↓ Levels of total cholesterol | ||||
| ↓ Triglycerides | ||||
| ↓ LDL cholesterol levels | ||||
| Pyrogallol-Phloroglucinol-6,6′-Bieckol (PPB)/Ecklonia cava | 2.5 mg/kg of PPB, oral gavage | C57BL/6N mice | ↓ Body weight | [96] |
| ↓ Visceral fat/adipocyte size | ||||
| ↓ Serum triglycerides | ||||
| ↓ Cholesterol | ||||
| ↓ Production of RAGE ligands in adipose tissues | ||||
| ↓ TNF-a mRNA expression | ||||
| Standard dose: 2.5 mg/kg/day, oral gavage | C57BL/6N mice | Improved blood circulation | [95] | |
| Standard dose: 2.5 mg/kg/day, oral gavage | C57BL/6N and leptin-deficient (ob/ob) mice | Restored brain leptin sensitivity | [97] | |
| ↑ Macrophage markers and proinflammatory cytokines | ||||
| ↑ TLR4 and NF-κB expression | ||||
| Indole derivative (indole-3- carbinol) | Standard dose: 5 mg/kg bw, intraperitoneally | C57BL/6J mice | Improved glucose tolerance | [99] |
| ↑ Serum adiponectin | ||||
| ↓ Serum glucose, triacylglycerol, insulin, leptin | ||||
| ↓ Expression of PPARγ and ACC | ||||
| Krill oil and Xanthigen | Standard dose: 25 g, dietary supplement | C57BL/6J mice | ↓ Triacylglycerol accumulation | [100] |
| ↓ Body weight, adipose mass | ||||
| Improved diet-induced hepatic steatosis | ||||
| Xanthigen | Standard dose: 1%, dietary supplement | C57BL/6J mice | ↓ PPARγ | [101] |
| Activation of AMPK- α and β, and ACC | ||||
| ↓ Expression of HMG-CoA reductase | ||||
| Diphlorethohydroxycarmalol (DPHC) | Standard dose: 100 mg/kg bw, oral gavage | Male ICR mice | ↓ Blood glucose | [103] |
| Inhibition of α- amylase | ||||
| Inhibition of α- glucosidase | ||||
| Low dose 25 mg/kg bw and high dose 50 mg/kg bw, oral gavage | C57BL/6J mice | ↓ SREBP-1c, FABP4, PPARγ, C/EBP and FAS | [102] | |
| ↓ Levels of triglycerides and low-density lipoprotein cholesterol | ||||
| ↓ Leptin and aspartate transaminase | ||||
| L-fucose | 0.03 g/kg bw, administered intragastrical | C57BL/6 mice | ↓ Weight gain and lipid accumulation | [105] |
| ↓ Hepatic triglyceride elevation |
| Genus | Species Names | Dosages | Models | Biological Effects | References |
|---|---|---|---|---|---|
| Sargassum | S. miyabei yendo | Standard dose: 20 mg/mL | 3T3-L1 adipocytes | Potent 2,2′-azinobis-3-ehtlbezothiazoline-6-sulfonic acid radical decolorization (ABTS) and 2,2-diphenyl-1-picrylhydrazyl (DPPH) antioxidant activity (IC50: 0.2868 ± 0.011 mg/mL and 0.2941 ± 0.014 mg/mL) | [69] |
| ↓ Expression of C/EBPα, PPARγ mRNA, SREBP1, FATP1, FAS, LPL, ACS1 and FABP4 | |||||
| Gelidium | G. amansii | Low dose 10, medium dose 20 and high dose 40 μg/mL | 3T3 L1 adipocytes | ↓ PPARγ and aP2 (adipocyte protein 2) | [116] |
| ↓ ROS-generator, NOX4 | |||||
| ↑ Adiponectin and GLUT4 | |||||
| Ecklonia | E. stolonifera | Dose range: 12–200 µg/mL | 3T3-L1 preadipocytes | ↓ Lipid accumulation, adipogenesis, adipocyte size | [117,118] |
| ↑ Lipolysis and browning of WAT | |||||
| ↑ MAPK, expression of lipolytic enzymes including ATGL, p-HSL and MGL. | |||||
| ↑ Thermogenic genes, CPT1, PRDM16 and UCP-1 | |||||
| E. cava | Dose range: 12.5–200 µg/mL | 3T3-L1 adipocytes | ↓ Expression of C/EBPα, SREBP1, FAS, LPL and FABP4 | [119] | |
| Gracilaria | G. verrucosa | Dose range: 1–40 mg/mL | 3T3-L1 adipocyte | ↓ Lipid accumulation and ROS production | [120] |
| Improved glucose uptake |
| In Vivo Studies | |||||
|---|---|---|---|---|---|
| Genus | Species | Dose/Concentration and Route of Administration | Models | Biological Effects | References |
| Sargassum sp. | S. siliquosum | Standard dose: 5%, dietary supplement | Male Wistar rats | Decreased body weight, retroperitoneal fat and liver fat | [108] |
| No changes in liver enzyme activities, systolic blood pressure, serum glucose, lipid profile and insulin metabolism | |||||
| S. horneri | Low dose 2% and high dose 6%, dietary supplement | C57BL/6J mice | ↓ Weight gain | [109] | |
| Improved insulin resistance | |||||
| Inhibited pancreatic lipase | |||||
| S. thunbergii | Low dose 100 mg/kg bw, high dose 300 mg/kg bw, dietary | Male C57BL/6 mice | ↓ Body weight and fat accumulation | [110] | |
| ↓ Serum insulin and triglycerides, liver fats and total cholesterol | |||||
| ↓ PPARγ | |||||
| ↑ UCP1 and 3 | |||||
| Gelidium sp. | G. elegans | Standard dose: 0.5%, dietary supplement | C57BL/6J-ob/ob mice | ↓ Body weight and fat accumulation | [112] |
| Low dose 50 mg/kg bw and high dose 200 mg/kg bw, oral gavage | Male ICR mice | Improved insulin resistance | [113] | ||
| ↓ Blood glucose | |||||
| G. amansii | Low dose 0.5 %, medium dose 1.0% and high dose 2.0 %, dietary supplement | C57BL/6J mice | ↓ Weight, adipose tissues and liver fat mass | [115] | |
| ↑ Plasma leptin, adiponectin levels | |||||
| ↓ Expression of C/EBPα, PPARγ mRNA, SREBP1, FATP1, FAS, LPL, ACS1 and FABP4 | |||||
| Standard dose: 3%, dietary supplement | Hamsters | ↓ Expression of C/EBPα, PPARγ mRNA, SREBP1, FATP1, FAS, LPL, ACS1 and FABP4 | [121] | ||
| ↓ Plasma and liver triglycerides and total cholesterol | |||||
| Standard dose: 5%, dietary supplement | Sprague Dawley rats | ↓ Plasma glucose | [122] | ||
| ↓ Plasma and liver triglycerides and total cholesterol | |||||
| ↓ Plasma adipokines | |||||
| Ecklonia sp. | E. stolonifera | Low dose 50 mg/kg bw, high dose 150 mg/kg bw, oral gavage | Male Institute of Cancer Research (ICR) mice/Male C57BL/6 mice | ↓ Serum concentrations of triglycerides, total cholesterol, and LDL | [117,118] |
| ↑ HDL | |||||
| Body mass | |||||
| Improved insulin resistance and regulation of blood glucose | |||||
| ↓ Muscle loss, ↑ expression of MRFs | |||||
| E. cava | 200 mg/kg bw, oral intubation | C57BL/6 mice | ↓ Body weight and hyperglycemia | [123] | |
| ↓ Lipid accumulation, ALT, cholesterol and adiposity | |||||
| ↑ mRNA expression of adipogenesis-related genes in adipose tissue | |||||
| Improved insulin resistance | |||||
| Low dose 5 mg/kg, medium dose 25 mg/kg and high dose 150 mg/kg, dietary supplement | C57BL/6 mouse | ↓ Body weight | [124] | ||
| ↓ Triglycerides, HDL, GOT, GPT | |||||
| ↓ Expression of C/EBPα, SREBP1, FAS, LPL and FABP4 | |||||
| Gracilaria sp. | G. birdiae | Standard dose: 6 mg/kg body weight, oral gavage | male Mus musculus mice | Reduced weight | [125] |
| ↓ CCI4-induced damage | |||||
| Clinical Trials | |||||
| Genus | Species | Dose/Concentration and Route of Administration | Participants | Biological Effects | References |
| Gelidium sp. | Gelidium elegans | 1000 mg/day, orally | n = 109 healthy adult volunteers | Waist circumference and hip circumference decreased | [114] |
| ↓ Fasting glucose, fasting insulin, triglyceride levels | |||||
4. Materials and Methods
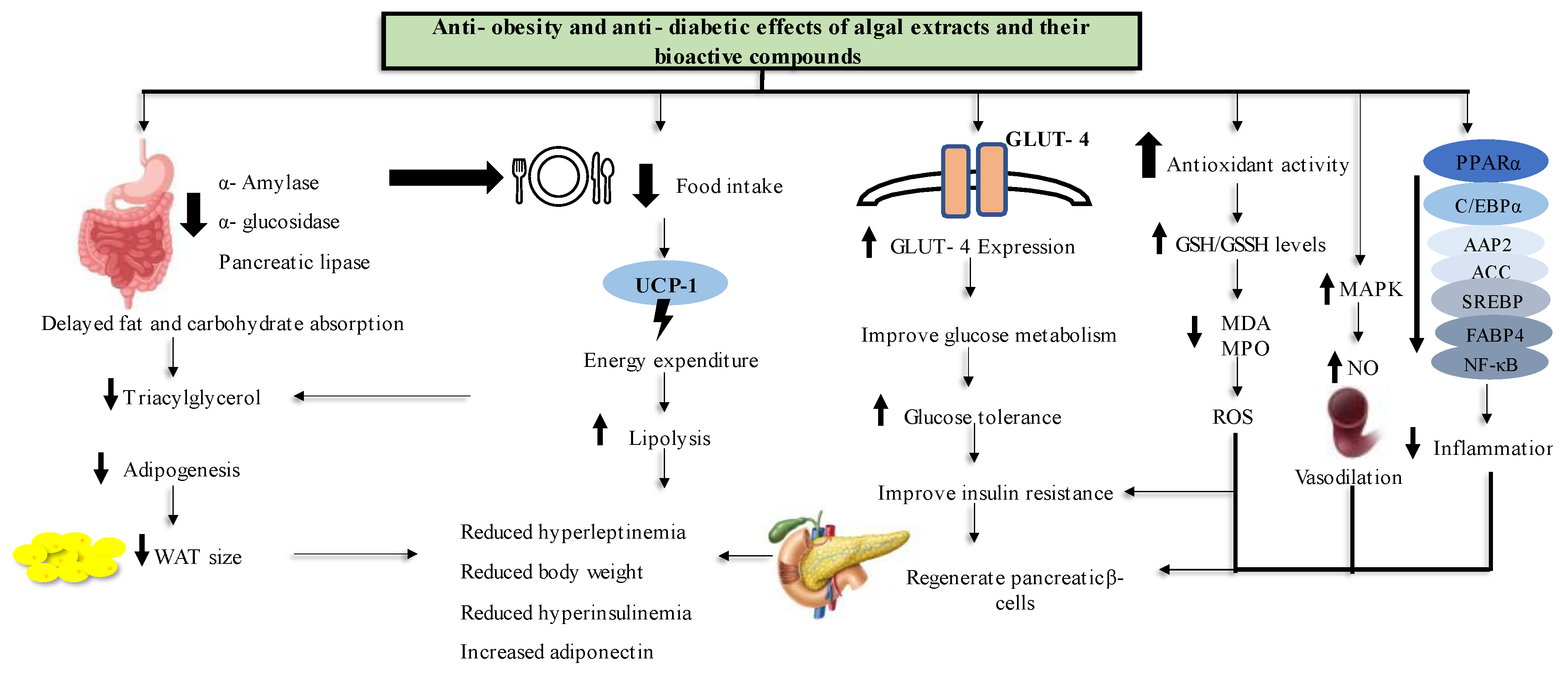
5. Conclusions
6. Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Goossens, G.H. The Metabolic Phenotype in Obesity: Fat Mass, Body Fat Distribution, and Adipose Tissue Function. Obes. Facts. 2017, 10, 207–215. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. World Health Statistics 2018: Monitoring Health for the SDGs, Sustainable Development Goals; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Han, J.C.; Lawlor, D.A.; Kim, S.Y. Childhood obesity. Lancet 2010, 375, 1737–1748. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, H.N.; MacCallum, P.R. The obesity, metabolic syndrome, and type 2 diabetes mellitus pandemic: Part I. Increased cardiovascular disease risk and the importance of atherogenic dyslipidemia in persons with the metabolic syndrome and type 2 diabetes mellitus. J. Cardiometab. Syndr. 2009, 4, 113–119. [Google Scholar] [CrossRef]
- Saltiel, A.R.; Kahn, R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef]
- Hribal, M.L.; Oriente, F.; Accili, D. Mouse models of insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2002, 282, 977–981. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.W.; Lee, M.; Oh, K.J. Adipose Tissue-Derived Signatures for Obesity and Type 2 Diabetes: Adipokines, Batokines and MicroRNAs. J. Clin. Med. 2019, 8, 854. [Google Scholar] [CrossRef] [PubMed]
- Kershaw, E.E.; Flier, J.S. Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef]
- World Health Organization. Diabetes. World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- International Diabetes Federation (IDF). Diabetes Atlas, 10th ed.; International Diabetes Federation: Brussels, Belgium, 2021. [Google Scholar]
- Boachie, M.K.; Thsehla, E.; Immurana, M.; Kohli-Lynch, C.; Hofman, K.J. Estimating the healthcare cost of overweight and obesity in South Africa. Glob. Health Action 2022, 15, 2045092. [Google Scholar] [CrossRef]
- Wadden, T.A.; Webb, V.L.; Moran, H.; Bailer, B.A. Lifestyle modification for obesity: New developments in diet, physical activity, and behavior therapy. Circulation 2012, 125, 1157–1170. [Google Scholar] [CrossRef]
- Khera, R.; Murad, M.H.; Chandar, A.K.; Dulai, P.S.; Wang, Z.; Prokop, L.J.; Loomba, R.; Camilleri, M.; Singh, S. Association of Pharmacological Treatments for Obesity with Weight Loss and Adverse Events: A Systematic Review and Meta-analysis. J. Am. Med. Assoc. 2016, 315, 2424–2434. [Google Scholar] [CrossRef]
- Brashier, D.B.; Sharma, A.K.; Dahiya, N.; Singh, S.K.; Khadka, A. Lorcaserin: A novel antiobesity drug. J. Pharmacol. Pharmacother. 2014, 5, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J. Metformin: Historical overview. Diabetologia 2017, 60, 1566–1576. [Google Scholar] [CrossRef] [PubMed]
- Kasina, S.V.S.K.; Baradhi, K.M. Dipeptidyl Peptidase IV (DPP IV) Inhibitors. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- MacArtain, P.; Gill, I.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional value of edible seaweeds. Nutr. Rev. 2007, 65, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.M.; Allsopp, P.J.; Magee, P.J.; Gill, I.; Nitecki, S.; Strain, R.; McSorley, E.M. Seaweed and human health. Nutr. Rev. 2014, 72, 205–216. [Google Scholar] [CrossRef]
- de Freitas Junior, L.M.; de Almeida, E.B., Jr. Medicinal plants for the treatment of obesity: Ethnopharmacological approach and chemical and biological studies. Am. J. Transl. Res. 2017, 9, 2050–2064. [Google Scholar]
- Kumar, P.; Bhandari, U. Common medicinal plants with antiobesity potential: A special emphasis on fenugreek. Anc. Sci. Life 2015, 35, 58–63. [Google Scholar] [CrossRef]
- Kazemipoor, M.; Jasimah, C.W.; Cordell, G.; Yaze, I. Potential of Traditional Medicinal Plants for Treating Obesity: A Review. ICNFS 2012, 39, 164–169. [Google Scholar]
- D’Orazio, N.; Gemello, E.; Gammone, M.A.; De Girolamo, M.; Ficoneri, C.; Riccioni, G. Fucoxantin: A treasure from the sea. Mar. Drugs 2012, 10, 604–616. [Google Scholar] [CrossRef]
- Kim, S.M.; Jung, Y.J.; Kwon, O.N.; Cha, K.H.; Um, B.H.; Chung, D.; Pan, H. A potential commercial source of fucoxanthin extracted from the microalga Phaeodactylum tricornutum. Appl. Biochem. Biotechnol. 2012, 166, 1843–1855. [Google Scholar] [CrossRef]
- Kim, S.M.; Kang, S.W.; Kwon, O.N.; Chung, D.; Pan, H. Fucoxanthin as a major carotenoid in Isochrysis aff. galbana: Characterization of extraction for commercial application. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 477–483. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Takahashi, N.; Kawada, T.; Miyashita, K. Fucoxanthin and its metabolite, fucoxanthinol, suppress adipocyte differentiation in 3T3-L1 cells. Int. J. Mol. Med. 2006, 18, 147–152. [Google Scholar] [CrossRef]
- Maeda, H.; Kanno, S.; Kodate, M.; Hosokawa, M.; Miyashita, K. Fucoxanthinol, Metabolite of Fucoxanthin, Improves Obesity-Induced Inflammation in Adipocyte Cells. Mar. Drugs 2015, 13, 4799–4813. [Google Scholar] [CrossRef] [PubMed]
- Gammone, M.A.; D’Orazio, N. Anti-obesity activity of the marine carotenoid fucoxanthin. Mar. Drugs 2015, 13, 2196–2214. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.X.; Wijesekara, I.; Li, Y.; Kim, S.K. Phlorotannins as bioactive agents from brown algae. Process Biochem. 2011, 46, 2219–2224. [Google Scholar] [CrossRef]
- Singh, I.; Bharate, S.B. Phloroglucinol compounds of natural origin. Nat. Prod. Rep. 2006, 23, 558–591. [Google Scholar] [CrossRef]
- Jung, H.A.; Jung, H.J.; Jeong, H.Y.; Kwon, H.J.; Ali, M.Y.; Choi, J.S. Phlorotannins isolated from the edible brown alga Ecklonia stolonifera exert anti-adipogenic activity on 3T3-L1 adipocytes by downregulating C/EBPα and PPARγ. Fitoterapia 2014, 92, 260–269. [Google Scholar] [CrossRef]
- Ko, S.C.; Lee, M.; Lee, J.H.; Lee, S.H.; Lim, Y.; Jeon, Y.J. Dieckol, a phlorotannin isolated from a brown seaweed, Ecklonia cava, inhibits adipogenesis through AMP-activated protein kinase (AMPK) activation in 3T3-L1 preadipocytes. Environ. Toxicol. Pharmacol. 2013, 36, 1253–1260. [Google Scholar] [CrossRef]
- Moon, H.E.; Islam, N.; Ahn, B.R.; Chowdhury, S.S.; Sohn, H.S.; Jung, H.A.; Choi, J.S. Protein tyrosine phosphatase 1B and α-glucosidase inhibitory Phlorotannins from edible brown algae, Ecklonia stolonifera and Eisenia bicyclis. Biosci. Biotechnol. Biochem. 2011, 75, 1472–1480. [Google Scholar] [CrossRef]
- Takaichi, S. Carotenoids in Algae: Distributions, Biosyntheses and Functions. Mar. Drugs 2011, 9, 1101–1118. [Google Scholar] [CrossRef]
- Li, Z.S.; Noda, K.; Fujita, E.; Manabe, Y.; Hirata, T.; Sugawara, T. The green algal carotenoid siphonaxanthin inhibits adipogenesis in 3T3-L1 preadipocytes and the accumulation of lipids in white adipose tissue of KK-Ay mice. J. Nutr. 2015, 145, 490–498. [Google Scholar] [CrossRef]
- Zheng, J.; Li, Z.; Manabe, Y.; Kim, M.; Goto, T.; Kawada, T.; Sugawara, T. Siphonaxanthin, a Carotenoid From Green Algae, Inhibits Lipogenesis in Hepatocytes via the Suppression of Liver X Receptor α Activity. Lipids 2018, 53, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Diraison, F.; Dusserre, E.; Vidal, H.; Sothier, M.; Beylot, M. Increased hepatic lipogenesis but decreased expression of lipogenic gene in adipose tissue in human obesity. Am. J. Physiol. Endocrinol. Metab. 2002, 282, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Tao, N.; Wang, X.; Xiao, J.W. Marine-derived bioactive compounds with anti-obesity effect: A review. J. Funct. Foods 2016, 21, 372–387. [Google Scholar] [CrossRef]
- Huang, J.S.; Barlow, S.E.; Quiros-Tejeira, R.E.; Scheimann, A.; Skelton, J.; Suskind, D.; Tsai, P.; Uko, V.; Warolin, J.P.; Xanthakos, S.A.; et al. Childhood obesity for pediatric gastroenterologists. J. Pediatr. Gastroenterol. Nutr. 2013, 56, 99–109. [Google Scholar] [CrossRef]
- Tsai, M.C.; Huang, S.C.; Chang, W.T.; Chen, S.C.; Hsu, L. Effect of Astaxanthin on the Inhibition of Lipid Accumulation in 3T3-L1 Adipocytes via Modulation of Lipogenesis and Fatty Acid Transport Pathways. Molecules 2020, 25, 3598. [Google Scholar] [CrossRef]
- Du, X.; Bai, M.; Huang, Y.; Jiang, Z.; Chen, F.; Ni, H.; Li, Q. Inhibitory effect of astaxanthin on pancreatic lipase with inhibition kinetics integrating molecular docking simulation. J. Funct. Foods. 2018, 48, 551–557. [Google Scholar] [CrossRef]
- Du, X.; Wang, X.; Yan, Y.; Yang, Z.; Jiang, H.; Ni, H. Hypoglycaemic effect of all-trans astaxanthin through inhibiting α-glucosidase. J. Funct. Foods 2020, 74, 104168. [Google Scholar] [CrossRef]
- Ambati, R.R.; Phang, S.M.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications—A review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef]
- Okada, T.; Nakai, M.; Maeda, H.; Hosokawa, M.; Sashima, T.; Miyashita, K. Suppressive effect of neoxanthin on the differentiation of 3T3-L1 adipose cells. J. Oleo. Sci. 2008, 57, 345–351. [Google Scholar] [CrossRef]
- Sekiya, M.; Suzuki, S.; Ushida, Y.; Suganuma, H. Neoxanthin in young vegetable leaves prevents fat accumulation in differentiated adipocytes. Biosci. Biotechnol. Biochem. 2021, 85, 2145–2152. [Google Scholar] [CrossRef]
- Wang, H.; Zhi-Ming, F.; Chun-Chao, H. The Potential Applications of Marine Bioactives Against Diabetes and Obesity. Am. J. Mar. Sci. 2014, 2, 1–8. [Google Scholar]
- Kim, M.J.; Chang, U.J.; Lee, J.S. Inhibitory effects of Fucoidan in 3T3-L1 adipocyte differentiation. Mar. Biotechnol. 2009, 11, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.J.; Lee, B.Y. Fucoidan from the sporophyll of Undaria pinnatifida suppresses adipocyte differentiation by inhibition of inflammation-related cytokines in 3T3-L1 cells. Nutr. Res. 2012, 32, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Koh, H.S.A.; Lu, J.; Zhou, W. Structural Dependence of Sulfated Polysaccharide for Diabetes Management: Fucoidan From Undaria pinnatifida Inhibiting α-Glucosidase More Strongly Than α-Amylase and Amyloglucosidase. Front. Pharmacol. 2020, 11, 831. [Google Scholar] [CrossRef] [PubMed]
- Vinoth Kumar, T.; Lakshmanasenthil, S.; Geetharamani, D.; Marudhupandi, T.; Suja, G.; Suganya, P. Fucoidan—A α-D-glucosidase inhibitor from Sargassum wightii with relevance to type 2 diabetes mellitus therapy. Int. J. Biol. Macromol. 2015, 72, 1044–1047. [Google Scholar] [CrossRef]
- Kim, K.T.; Rioux, L.E.; Turgeon, S.L. Molecular weight and sulfate content modulate the inhibition of α-amylase by fucoidan relevant for type 2 diabetes management. Pharma Nutr. 2015, 3, 108–114. [Google Scholar] [CrossRef]
- Park, M.K.; Jung, U.; Roh, C. Fucoidan from Marine Brown Algae Inhibits Lipid Accumulation. Mar. Drugs 2011, 9, 1359–1367. [Google Scholar] [CrossRef]
- Draget, K.I.; Taylor, C. Chemical, physical and biological properties of alginates and their biomedical implications. Food Hydrocoll. 2011, 25, 251–256. [Google Scholar] [CrossRef]
- Wan-Loy, C.; Siew-Moi, P. Marine Algae as a Potential Source for Anti-Obesity Agents. Mar. Drugs 2016, 14, 222. [Google Scholar] [CrossRef]
- Lange, K.W.; Hauser, J.; Nakamura, Y.; Kanaya, S. Dietary seaweeds and obesity. Food Sci. Hum. Wellness. 2015, 4, 87–96. [Google Scholar] [CrossRef]
- Houghton, D.; Wilcox, M.D.; Chater, P.I.; Brownlee, I.A.; Seal, J.; Pearson, J.P. Biological activity of alginate and its effect on pancreatic lipase inhibition as a potential treatment for obesity. Food Hydrocoll. 2015, 49, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, B.; Xue, C.; Sun, L. Effect of molecular weight on the antioxidant property of low molecular weight alginate from Laminaria japonica. J. Appl. Phycol. 2012, 24, 295–300. [Google Scholar] [CrossRef]
- Grasa-López, A.; Miliar-García, Á.; Quevedo-Corona, L.; Paniagua-Castro, N.; Escalona-Cardoso, G.; Reyes-Maldonado, E.; Jaramillo-Flores, M.E. Undaria pinnatifida and Fucoxanthin Ameliorate Lipogenesis and Markers of Both Inflammation and Cardiovascular Dysfunction in an Animal Model of Diet-Induced Obesity. Mar. Drugs 2016, 14, 148. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Murakami-Funayama, K.; Miyashita, K. Anti-obesity and anti-diabetic effects of fucoxanthin on diet-induced obesity conditions in a murine model. Mol. Med. Rep. 2009, 2, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Miyashita, K. Dietary combination of fucoxanthin and fish oil attenuates the weight gain of white adipose tissue and decreases blood glucose in obese/diabetic KK-Ay mice. J. Agric. Food Chem. 2007, 55, 7701–7706. [Google Scholar] [CrossRef]
- Tan, P.; Hou, Y.H. First evidence for the anti-inflammatory activity of fucoxanthin in high-fat-diet-induced obesity in mice and the antioxidant functions in PC12 cells. Inflammation 2014, 37, 443–450. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, W.; Huang, X.; Zhao, Y.; Ren, Q.; Hong, Z.; Huang, M.; Xing, X. Fucoxanthin ameliorates hyperglycemia, hyperlipidemia and insulin resistance in diabetic mice partially through IRS-1/PI3K/Akt and AMPK pathway. J. Funct. Foods. 2018, 48, 515–524. [Google Scholar] [CrossRef]
- Gheda, S.; Naby, M.A.; Mohamed, T.; Pereira, L.A. Antidiabetic and antioxidant activity of phlorotannins extracted from the brown seaweed Cystoseira compressa in streptozotocin-induced diabetic rats. Envrion. Sci. Poll. Res. Int. 2021, 28, 22886–22901. [Google Scholar] [CrossRef]
- Erpel, F.; Mateos, R.; Pérez-Jiménez, J.; Pérez-Correa, J.R. Phlorotannins: From isolation and structural characterization, to the evaluation of their antidiabetic and anticancer potential. Food Res. Int. 2020, 137, 109589. [Google Scholar] [CrossRef]
- Zheng, J.; Manabe, Y.; Sugawara, T. Siphonaxanthin, a carotenoid from green algae Codium cylindricum, protects Ob/Ob mice fed on a high-fat diet against lipotoxicity by ameliorating somatic stresses and restoring anti-oxidative capacity. Nutr. Res. 2020, 77, 29–42. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Koyama, T.; Takahashi, J.; Yazawa, K. Effects of astaxanthin in obese mice fed a high-fat diet. Biosci. Biotechnol. Biochem. 2007, 71, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Aoi, W.; Naito, Y.; Takanami, Y.; Ishii, T.; Kawai, Y.; Akagiri, S.; Kato, Y.; Osawa, T.; Yoshikawa, T. Astaxanthin improves muscle lipid metabolism in exercise via inhibitory effect of oxidative CPT I modification. Biochem. Biophys. Res. Commun. 2008, 366, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Bhuvaneswari, S.; Arunkumar, E.; Viswanathan, P.; Anuradha, V. Astaxanthin restricts weight gain, promotes insulin sensitivity and curtails fatty liver disease in mice fed an obesity-promoting diet. Process Biochem. 2010, 45, 1406–1414. [Google Scholar] [CrossRef]
- Chen, Y.; Tang, J.; Zhang, Y.; Du, J.; Wang, Y.; Yu, H.; He, Y. Astaxanthin alleviates gestational diabetes mellitus in mice through suppression of oxidative stress. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 2517–2527. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Lee, S.G.; Kim, M.; Hahn, D.; Jung, S.K.; Cho, T.O.; Nam, J.O. Sargassum miyabei Yendo Brown Algae Exert Anti-Oxidative and Anti-AdipogenicEffects on 3T3-L1 Adipocytes by Downregulating PPARγ. Medicina 2020, 56, 634. [Google Scholar] [CrossRef]
- Zhang, Y.; Zuo, J.; Yan, L.; Cheng, Y.; Li, Q.; Wu, S.; Chen, L.; Thring, R.W.; Yang, Y.; Gao, Y.; et al. Sargassum fusiforme Fucoidan Alleviates High-Fat Diet-Induced Obesity and Insulin Resistance Associated with the Improvement of Hepatic Oxidative Stress and Gut Microbiota Profile. J. Agric. Food Chem. 2020, 68, 10626–10638. [Google Scholar] [CrossRef]
- Kim, K.J.; Yoon, K.Y.; Lee, B.Y. Fucoidan regulate blood glucose homeostasis in C57BL/KSJ m+/+db and C57BL/KSJ db/db mice. Fitoterapia 2012, 83, 1105–1109. [Google Scholar] [CrossRef]
- Wang, X.; Liu, F.; Gao, Y.; Xue, H.; Li, R.W.; Tang, Q.J. Transcriptome analysis revealed anti-obesity effects of the Sodium Alginate in high-fat diet-induced obese mice. Int. J. Biol. Marcomol. 2018, 115, 861–870. [Google Scholar] [CrossRef]
- Li, S.; Wang, L.; Liu, B.; He, N. Unsaturated alginate oligosaccharides attenuated obesity-related metabolic abnormalities by modulating gut microbiota in high-fat-diet mice. Food Funct. 2020, 11, 4773–4784. [Google Scholar] [CrossRef]
- Dufrane, D.; Goebbels, R.M.; Gianello, P. Alginate macroencapsulation of pig islets allows correction of streptozotocin-induced diabetes in primates up to 6 months without immunosuppression. Transplantation 2010, 90, 1054–1062. [Google Scholar] [CrossRef]
- Choi, H.D.; Kim, J.H.; Chang, M.J.; Kyu-Youn, Y.; Shin, W.G. Effects of astaxanthin on oxidative stress in overweight and obese adults. PTR 2011, 25, 1813–1818. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, D.; Goff, H.D.; Berengut, S.; Kubant, R.; Anderson, G.H. Effect of sodium alginate addition to chocolate milk on glycemia, insulin, appetite and food intake in healthy adult men. Eur. J. Clin. Nutr. 2014, 68, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Georg Jensen, M.; Pedersen, C.; Kristensen, M.; Frost, G.; Astrup, A. Review: Efficacy of alginate supplementation in relation to appetite regulation and metabolic risk factors: Evidence from animal and human studies. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2014, 14, 129–144. [Google Scholar] [CrossRef]
- Hernández-Corona, D.M.; Martínez-Abundis, E.; González-Ortiz, M. Effect of fucoidan administration on insulin secretion and insulin resistance in overweight or obese adults. J. Med. Food. 2014, 17, 830–832. [Google Scholar] [CrossRef] [PubMed]
- Paxman, J.R.; Richardson, J.C.; Dettmar, P.W.; Corfe, B.M. Daily ingestion of alginate reduces energy intake in free-living subjects. Appetite 2008, 51, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Beppu, F.; Hosokawa, M.; Niwano, Y.; Miyashita, K. Effects of dietary fucoxanthin on cholesterol metabolism in diabetic/obese KK-A(y) mice. Lipids Health Dis. 2012, 11, 112. [Google Scholar] [CrossRef]
- Nishikawa, S.; Hosokawa, M.; Miyashita, K. Fucoxanthin promotes translocation and induction of glucose transporter 4 in skeletal muscles of diabetic/obese KK-A(y) mice. Phytomedicine 2012, 19, 389–394. [Google Scholar] [CrossRef]
- Oliyaei, N.; Moosavi-Nasab, M.; Tamaddon, A.M.; Tanideh, N. Antidiabetic effect of fucoxanthin extracted from Sargassum angustifolium on streptozotocin-nicotinamide-induced type 2 diabetic mice. Food Sci. Nutr. 2021, 9, 3521–3529. [Google Scholar] [CrossRef]
- Feng, W.; Wang, Y.; Guo, N.; Huang, P.; Mi, Y. Effects of Astaxanthin on Inflammation and Insulin Resistance in a Mouse Model of Gestational Diabetes Mellitus. Dose-Response 2020, 18, 1559325820926765. [Google Scholar] [CrossRef]
- Kim, M.J.; Jeon, J.; Lee, J.S. Fucoidan prevents high-fat diet-induced obesity in animals by suppression of fat accumulation. Phytother. Res. 2014, 28, 137–143. [Google Scholar] [CrossRef]
- Kim, M.M.; Ta, Q.V.; Mendis, E.; Rajapakse, N.; Jung, W.K.; Byun, H.G.; Jeon, Y.J.; Kim, S.K. Phlorotannins in Ecklonia cava extract inhibit matrix metalloproteinase activity. Life Sci. 2006, 79, 1436–1443. [Google Scholar] [CrossRef] [PubMed]
- Yeo, A.R.; Lee, J.; Tae, I.H.; Park, S.R.; Cho, Y.H.; Lee, B.H.; Shin, H.C.; Kim, S.H.; Yoo, Y.C. Anti-hyperlipidemic Effect of Polyphenol Extract (Seapolynol(™)) and Dieckol Isolated from Ecklonia cava in in vivo and in vitro Models. Prev. Nutr. Food Sci. 2012, 17, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.A. Lipase Inhibitory Mode of Dieckol Isolated from Eisenia bicyclis Ethanol Extract. Korean J. Microbiol. Biotechnol. 2013, 41, 112–118. [Google Scholar] [CrossRef]
- Kim, S.K.; Kong, S. Anti-adipogenic effect of dioxinodehydroeckol via AMPK activation in 3T3-L1 adipocytes. Chemico-Biological Interact. 2010, 186, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.H.; Lee, S.H.; Yoon, N.Y.; Jung, W.K.; Jeon, Y.J.; Kim, S.K.; Lee, M.S.; Kim, Y.M. α-Glucosidase- and α-amylase-inhibitory activities of phlorotannins from Eisenia bicyclis. J. Sci. Food Agric. 2012, 92, 2084–2090. [Google Scholar] [CrossRef]
- Kim, A.R.; Shin, T.S.; Lee, M.S.; Park, J.Y.; Park, K.E.; Yoon, N.Y.; Kim, J.S.; Choi, J.S.; Jang, B.C.; Byun, D.S.; et al. Isolation and identification of phlorotannins from Ecklonia stolonifera with antioxidant and anti-inflammatory properties. J. Agric. Food Chem. 2009, 57, 3483–3489. [Google Scholar] [CrossRef]
- Kang, M.C.; Ding, Y.; Kim, E.A.; Choi, Y.K.; de Araujo, T.; Heo, S.J.; Lee, S.H. Indole Derivatives Isolated from Brown Alga Sargassum thunbergii Inhibit Adipogenesis through AMPK Activation in 3T3-L1 Preadipocytes. Mar. Drugs 2017, 15, 119. [Google Scholar] [CrossRef]
- Lee, S.H.; Park, M.H.; Heo, S.J.; Kang, S.M.; Ko, S.C.; Han, J.S.; Jeon, Y.J. Dieckol isolated from Ecklonia cava inhibits alpha-glucosidase and alpha-amylase in vitro and alleviates postprandial hyperglycemia in streptozotocin-induced diabetic mice. Food Chem. Toxicol. 2010, 48, 2633–2637. [Google Scholar] [CrossRef]
- Choi, H.S.; Jeon, H.J.; Lee, O.H.; Lee, B.Y. Dieckol, a major phlorotannin in Ecklonia cava, suppresses lipid accumulation in the adipocytes of high-fat diet-fed zebrafish and mice: Inhibition of early adipogenesis via cell-cycle arrest and AMPKα activation. Mol. Nutr. Food Res. 2015, 59, 1458–1471. [Google Scholar] [CrossRef]
- Jeon, H.; Yoon, K.; Koh, E.; Choi, J.; Kim, K.; Choi, H.; Lee, B. Seapolynol and Dieckol Improve Insulin Sensitivity through the Regulation of the PI3K Pathway in C57BL/KsJ-db/db Mice. J. Food Nutr. Res. 2015, 3, 648–652. [Google Scholar] [CrossRef]
- Son, M.; Oh, S.; Lee, H.S.; Ryu, B.; Jiang, Y.; Jang, J.T.; Jeon, Y.J.; Byun, K. Pyrogallol-Phloroglucinol-6,6’-Bieckol from Ecklonia cava Improved Blood Circulation in Diet-Induced Obese and Diet-Induced Hypertension Mouse Models. Mar. Drugs 2019, 17, 272. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Oh, S.; Son, M.; Byun, K. Pyrogallol-Phloroglucinol-6,6-Bieckol Alleviates Obesity and Systemic Inflammation in a Mouse Model by Reducing Expression of RAGE and RAGE Ligands. Mar. Drugs 2019, 17, 612. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Oh, S.; Choi, J.; Jang, J.T.; Choi, H.; Park, K.Y.; Son, K.H.; Byun, K. Attenuation of Inflammation and Leptin Resistance by Pyrogallol-Phloroglucinol-6,6-Bieckol on in the Brain of Obese Animal Models. Nutrients 2019, 11, 2773. [Google Scholar] [CrossRef] [PubMed]
- Baldrick, F.R.; McFadden, K.; Ibars, M.; Sung, C.; Moffatt, T.; Megarry, K.; Thomas, K.; Mitchell, P.; Wallace, J.M.W.; Pourshahidi, L.K.; et al. Impact of a (poly)phenol-rich extract from the brown algae Ascophyllum nodosum on DNA damage and antioxidant activity in an overweight or obese population: A randomized controlled trial. Am. J. Clin. Nutr. 2018, 108, 688–700. [Google Scholar] [CrossRef]
- Chang, H.P.; Wang, M.L.; Chan, M.H.; Chiu, Y.S.; Chen, Y.H. Antiobesity activities of indole-3-carbinol in high-fat-diet-induced obese mice. Nutrition 2011, 27, 463–470. [Google Scholar] [CrossRef]
- Lee, M.; Lai, C.; Cheng, A.; Hou, J.; Badmaev, V.; Ho, C.; Pan, M. Krill oil and xanthigen separately inhibit high fat diet induced obesity and hepatic triacylglycerol accumulation in mice. J. Funct. Foods 2014, 19, 913–921. [Google Scholar] [CrossRef]
- Choi, K.M.; Jeon, Y.S.; Kim, W. Xanthigen attenuates high-fat diet-induced obesity through down-regulation of PPARγ and activation of the AMPK pathway. Food Sci. Biotechnol. 2014, 23, 931–935. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, L.; Im, S.; Hwang, O.; Kim, H.S.; Kang, M.C.; Lee, S.H. Anti-Obesity Effect of Diphlorethohydroxycarmalol Isolated from Brown Alga Ishige okamurae in High-Fat Diet-Induced Obese Mice. Mar. Drugs 2019, 17, 637. [Google Scholar] [CrossRef]
- Heo, S.J.; Hwang, J.Y.; Choi, J.I.; Han, J.S.; Kim, H.J.; Jeon, Y.J. Diphlorethohydroxycarmalol isolated from Ishige okamurae, a brown algae, a potent alpha-glucosidase and alpha-amylase inhibitor, alleviates postprandial hyperglycemia in diabetic mice. Eur. J. Pharmacol. 2009, 615, 252–256. [Google Scholar] [CrossRef]
- Yuan, X.; Nakao, T.; Satone, H.; Ohara, K.; Kominami, Y.; Ito, M.; Aizawa, T.; Ueno, T.; Ushio, H. The Effects of Brown Algae-Derived Monosaccharide L-Fucose on Lipid Metabolism in C57BL/6J Obese Mice. Nutrients 2020, 12, 3798. [Google Scholar] [CrossRef]
- Wu, G.; Niu, M.; Tang, W.; Hu, J.; Wei, G.; He, Z.; Chen, Y.; Jiang, Y.; Chen, P. L-Fucose ameliorates high-fat diet-induced obesity and hepatic steatosis in mice. J. Trnals. Med. 2018, 16, 344. [Google Scholar] [CrossRef] [PubMed]
- Yip, Z.T.; Quek, R.Z.B.; Huang, D. Historical biogeography of the widespread macroalga Sargassum (Fucales, Phaeophyceae). J. Phycol. 2020, 56, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Yende, S.R.; Harle, U.N.; Chaugule, B.B. Therapeutic potential and health benefits of Sargassum species. Pharmacogn. Rev. 2014, 8, 1–7. [Google Scholar] [CrossRef]
- du Preez, R.; Magnusson, M.; Majzoub, M.E.; Thomas, T.; Praeger, C.; Glasson, R.K.; Panchal, S.K.; Brown, L. Brown Seaweed Sargassum siliquosum as an Intervention for Diet-Induced Obesity in Male Wistar Rats. Nutrients 2021, 13, 1754. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Hirazawa, C.; Ohya, T.; Yoshikawa, R.; Mizutani, T.; Ma, N.; Moriyama, M.; Ito, T.; Matsuzaki, C. The Edible Brown Seaweed Sargassum horneri (Turner) Agardh Ameliorates High-Fat Diet-Induced Obesity, Diabetes, and Hepatic Steatosis in Mice. Nutrients 2021, 13, 551. [Google Scholar] [CrossRef]
- Kang, M.C.; Lee, H.G.; Kim, H.S.; Song, K.M.; Chun, Y.G.; Lee, M.H.; Kim, B.K.; Jeon, Y.J. Anti-Obesity Effects of Sargassum thunbergii via Downregulation of Adipogenesis Gene and Upregulation of Thermogenic Genes in High-Fat Diet-Induced Obese Mice. Nutrients 2020, 12, 3325. [Google Scholar] [CrossRef]
- Qin, Y. Bioactive Seaweeds for Food Applications; Academic Press: London, UK, 2018; pp. 3–24. [Google Scholar]
- Park, M.H.; Kang, J.H.; Kim, H.J.; Han, J.S. Gelidium amansii ethanol extract suppresses fat accumulation by down-regulating adipogenic transcription factors in ob/ob mice model. Food Sci. Biotechnol. 2017, 26, 207–212. [Google Scholar] [CrossRef]
- Choi, J.; Kim, K.J.; Koh, E.J.; Lee, B.Y. Gelidium elegans Regulates the AMPK-PRDM16-UCP-1 Pathway and Has a Synergistic Effect with Orlistat on Obesity-Associated Features in Mice Fed a High-Fat Diet. Nutrients 2017, 9, 342. [Google Scholar] [CrossRef]
- Kim, O.; Kim, Y.N.; Lee, D.C. Effects of Gelidium elegans on Weight and Fat Mass Reduction and Obesity Biomarkers in Overweight or Obese Adults: A Randomized Double-Blinded Study. Nutrients 2019, 11, 1513. [Google Scholar] [CrossRef]
- Kang, J.H.; Lee, H.A.; Kim, H.J.; Han, J.S. Gelidium amansii extract ameliorates obesity by down-regulating adipogenic transcription factors in diet-induced obese mice. Nutr. Res. Pract. 2017, 11, 17–24. [Google Scholar] [CrossRef]
- Seo, M.J.; Lee, O.H.; Choi, H.S.; Lee, B.Y. Extract from Edible Red Seaweed (Gelidium amansii) Inhibits Lipid Accumulation and ROS Production during Differentiation in 3T3-L1 Cells. Prev. Nutr. Food Sci. 2020, 17, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Lee, K.; Chei, S.; Oh, H.J.; Lee, K.P.; Lee, B.Y. Ecklonia stolonifera Extract Suppresses Lipid Accumulation by Promoting Lipolysis and Adipose Browning in High-Fat Diet-Induced Obese Male Mice. Cells 2020, 9, 871. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Oh, H.J.; Lee, K.; Han, X.; Lee, O.; Lee, B. Effects of Ecklonia stolonifera extract on the obesity and skeletal muscle regeneration in high-fat diet-fed mice. J. Funct. Foods. 2021, 82, 104511. [Google Scholar] [CrossRef]
- Kim, I.H.; Nam, T.J. Enzyme-treated Ecklonia cava extract inhibits adipogenesis through the downregulation of C/EBPα in 3T3-L1 adipocytes. Int. J. Mol. Med. 2017, 39, 636–644. [Google Scholar] [CrossRef]
- Woo, M.S.; Choi, H.S.; Lee, O.H.; Lee, B.Y. The edible red alga, Gracilaria verrucosa, inhibits lipid accumulation and ROS production, but improves glucose uptake in 3T3-L1 cells. Phytother. Res. 2013, 27, 1102–1105. [Google Scholar] [CrossRef]
- Yang, T.H.; Chiu, Y.; Lu, T.J.; Liu, S.H.; Chiang, M.T. The Anti-Obesity Effect of Polysaccharide-Rich Red Algae (Gelidium amansii) Hot-Water Extracts in High-Fat Diet-Induced Obese Hamsters. Mar. Drugs 2019, 17, 532. [Google Scholar] [CrossRef]
- Yang, T.H.; Yao, H.T.; Chiang, M.T. Red algae (Gelidium amansii) reduces adiposity via activation of lipolysis in rats with diabetes induced by streptozotocin-nicotinamide. J. Food Drug Analysis. 2015, 23, 758–765. [Google Scholar] [CrossRef]
- Park, E.Y.; Kim, E.H.; Kim, M.H.; Seo, Y.W.; Lee, J.I.; Jun, H.S. Polyphenol-Rich Fraction of Brown Alga Ecklonia cava Collected from Gijang, Korea, Reduces Obesity and Glucose Levels in High-Fat Diet-Induced Obese Mice. Evid. Based Complement Altern. Med. 2012, 2012, 418912. [Google Scholar] [CrossRef]
- Kim, I.H.; Choi, J.W.; Lee, M.K.; Kwon, J.; Nam, T.J. Anti-obesity effects of pectinase and cellulase enzyme-treated Ecklonia cava extract in high-fat diet-fed C57BL/6N mice. Int. J. Mol. Med. 2018, 41, 924–934. [Google Scholar]
- Barros-Gomes, J.A.C.; Nascimento, D.L.A.; Silveira, A.C.R.; Silva, R.K.; Gomes, D.L.; Melo, K.R.T.; Almeida-Lima, J.; Camara, R.B.G.; Silva, N.B.; Rocha, H.A.O. In Vivo Evaluation of the Antioxidant Activity and Protective Action of the Seaweed Gracilaria birdiae. Oxidat. Med. Cell. Longev. 2018, 2018, 9354296. [Google Scholar] [CrossRef]
- Kang, C.; Jin, Y.B.; Lee, H.; Cha, M.; Sohn, E.T.; Moon, J.; Park, C.; Chun, S.; Jung, E.S.; Hong, J.S.; et al. Brown alga Ecklonia cava attenuates type 1 diabetes by activating AMPK and Akt signaling pathways. Food Chem. Toxicol. 2021, 48, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Gunathilaka, T.L.; Samarakoon, K.W.; Ranasinghe, P.; Peiris, L.D.C. In-Vitro Antioxidant, Hypoglycemic Activity, and Identification of Bioactive Compounds in Phenol-Rich Extract from the Marine Red Algae Gracilaria edulis (Gmelin) Silva. Molecules 2019, 24, 3708. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magwaza, S.N.; Islam, M.S. Roles of Marine Macroalgae or Seaweeds and Their Bioactive Compounds in Combating Overweight, Obesity and Diabetes: A Comprehensive Review. Mar. Drugs 2023, 21, 258. https://doi.org/10.3390/md21040258
Magwaza SN, Islam MS. Roles of Marine Macroalgae or Seaweeds and Their Bioactive Compounds in Combating Overweight, Obesity and Diabetes: A Comprehensive Review. Marine Drugs. 2023; 21(4):258. https://doi.org/10.3390/md21040258
Chicago/Turabian StyleMagwaza, S’thandiwe Nozibusiso, and Md. Shahidul Islam. 2023. "Roles of Marine Macroalgae or Seaweeds and Their Bioactive Compounds in Combating Overweight, Obesity and Diabetes: A Comprehensive Review" Marine Drugs 21, no. 4: 258. https://doi.org/10.3390/md21040258
APA StyleMagwaza, S. N., & Islam, M. S. (2023). Roles of Marine Macroalgae or Seaweeds and Their Bioactive Compounds in Combating Overweight, Obesity and Diabetes: A Comprehensive Review. Marine Drugs, 21(4), 258. https://doi.org/10.3390/md21040258