

Study of the Antimicrobial Potential of the Arthrospira platensis, Planktothrix agardhii, Leptolyngbya cf. ectocarpi, Roholtiella mixta nov., Tetraselmis viridis, and Nanofrustulum shiloi against Gram-Positive, Gram-Negative Bacteria, and Mycobacteria

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Cyanobacteria and Microalgae’s Biochemical Profiles
2.2. The Ceanobacteria and Microalgae Extract’s Cytotoxic Potential
2.3. Cyanobacteria and Microalgae Extracts Have Antibacterial Properties
2.4. Microalgae’s Antimycobacterial Potential
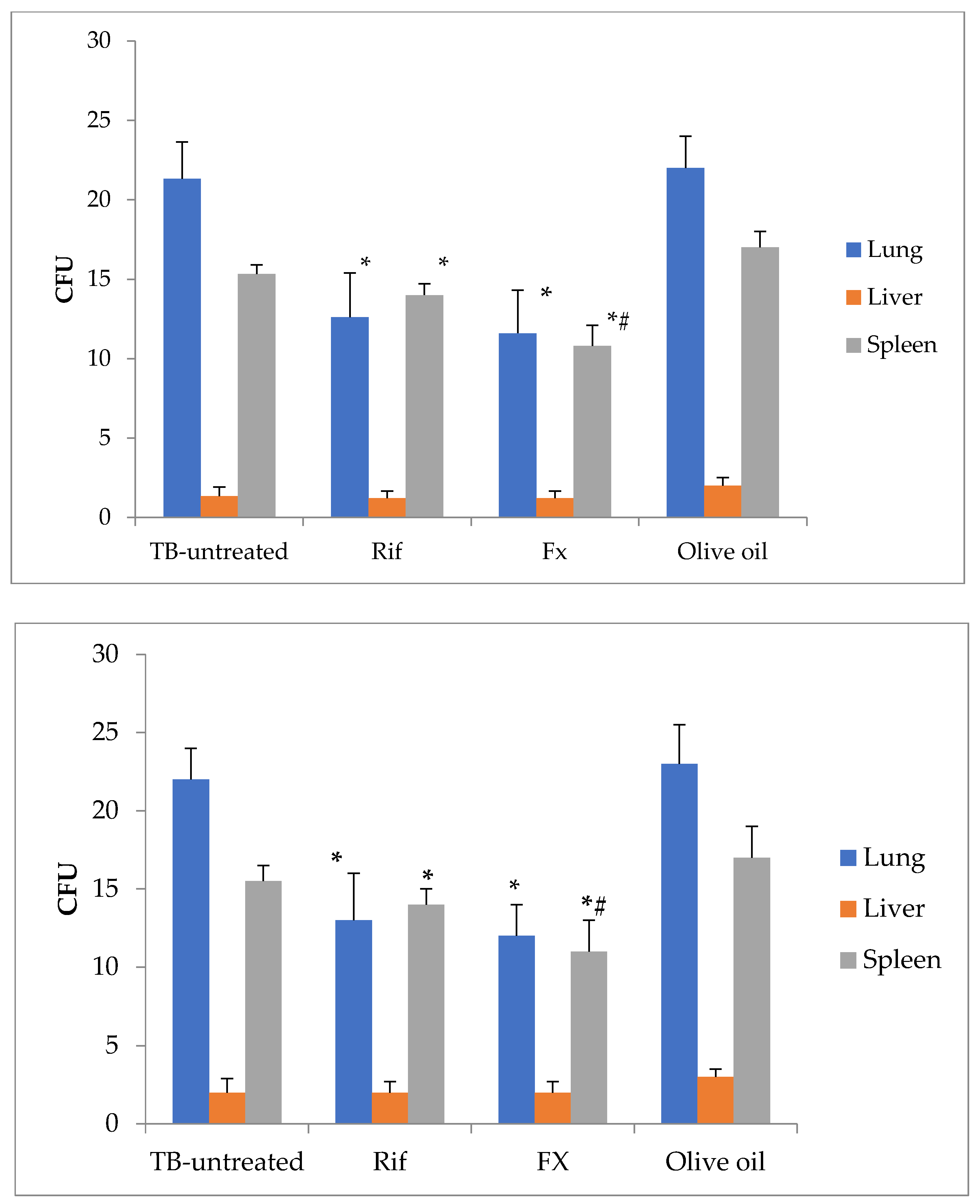
2.5. Effect of Fucoxanthin Administration in Tuberculosis-Infected Mice
3. Discussion
4. Materials and Methods
4.1. Microalgae
4.2. Chemical Analysis
4.3. Fucoxanthin Extraction
4.4. Microalgae Extracts
4.5. Cytotoxicity Assay
4.6. Antimicrobial Assay
4.7. Mice Model of Tuberculosis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mitishev, A.; Kurdyukov, E.; Rodina, O.; Semenova, E.; Moiseeva, I.; Fadeeva, T. Microalgae as a new source of biologically active compounds with antibacterial activity. Probl. Biol. Med. Pharm. Chem. 2021, 24, 24–29. [Google Scholar] [CrossRef]
- Cakmak, Y.S.; Kaya, M.; Asan-Ozusaglam, M. Biochemical composition and bioactivity screening of various extracts from Dunaliella salina, a green microalga. EXCLI J. 2014, 13, 679–690. [Google Scholar]
- de Morais, M.G.; Vaz Bda, S.; de Morais, E.G.; Costa, J.A. Biologically Active Metabolites Synthesized by Microalgae. BioMed Res. Int. 2015, 2015, 835761. [Google Scholar] [CrossRef]
- Nieri, P.; Carpi, S.; Esposito, R.; Costantini, M.; Zupo, V. Bioactive Molecules from Marine Diatoms and Their Value for the Nutraceutical Industry. Nutrients 2023, 15, 464. [Google Scholar] [CrossRef]
- Grubišić, M.; Šantek, B.; Zorić, Z.; Čošić, Z.; Vrana, I.; Gašparović, B.; Čož-Rakovac, R.; Ivančić Šantek, M. Bioprospecting of Microalgae Isolated from the Adriatic Sea: Characterization of Biomass, Pigment, Lipid and Fatty Acid Composition, and Antioxidant and Antimicrobial Activity. Molecules 2022, 27, 1248. [Google Scholar] [CrossRef]
- Santiago-Morales, I.S.; Trujillo-Valle, L.; Márquez-Rochoc, S.; Trujillo-Valle, L.; Márquez-Rocha, F.J.; Hernández, J.F.L. Tocopherols, phycocyanin and superoxide dismutase from microalgae: As potential food antioxidants. Appl. Food Biotechnol. 2018, 5, 19–27. [Google Scholar] [CrossRef]
- Hu, J.; Nagarajan, D.; Zhang, Q.; Chang, J.-S.; Lee, D.-J. Heterotrophic cultivation of microalgae for pigment production: A review. Biotechnol. Adv. 2018, 36, 54–67. [Google Scholar] [CrossRef]
- Mazumdar, N.; Novis, P.M.; Visnovsky, G.; Gostomski, P.A. Nutrient effects on the growth of a novel alpine Haematococcus (Chlorophyceae) strain from New Zealand. Phycol. Res. 2019, 67, 21–27. [Google Scholar] [CrossRef]
- Falaise, C.; François, C.; Travers, M.-A.; Morga, B.; Haure, J.; Tremblay, R.; Mouget, J.-L. Antimicrobial Compounds from Eukaryotic Microalgae against Human Pathogens and Diseases in Aquaculture. Mar. Drugs. 2016, 14, 159. [Google Scholar] [CrossRef]
- Tolpeznikaite, E.; Bartkevics, V.; Ruzauskas, M.; Pilkaityte, R.; Viskelis, P.; Urbonaviciene, D.; Zavistanaviciute, P.; Zokaityte, E.; Ruibys, R.; Bartkiene, E. Characterization of Macro- and Microalgae Extracts Bioactive Compounds and Micro- and Macroelements Transition from Algae to Extract. Foods 2021, 10, 2226. [Google Scholar] [CrossRef]
- Cepas, V.; Gutiérrez-Del-Río, I.; López, Y.; Redondo-Blanco, S.; Gabasa, Y.; Iglesias, M.J.; Soengas, R.; Fernández-Lorenzo, A.; López-Ibáñez, S.; Villar, C.J.; et al. Microalgae and Cyanobacteria Strains as Producers of Lipids with Antibacterial and Antibiofilm Activity. Mar. Drugs 2021, 19, 75. [Google Scholar] [CrossRef]
- Zaharieva, M.M.; Zheleva-Dimitrova, D.; Rusinova-Videva, S.; Ilieva, Y.; Brachkova, A.; Balabanova, V.; Gevrenova, R.; Kim, T.C.; Kaleva, M.; Georgieva, A.; et al. Antimicrobial and Antioxidant Potential of Scenedesmus obliquus Microalgae in the Context of Integral Biorefinery Concept. Molecules 2022, 27, 519. [Google Scholar] [CrossRef]
- Kumar, T.S.; Josephine, A.; Sreelatha, T.; Azger Dusthackeer, V.N.; Mahizhaveni, B.; Dharani, G.; Kirubagaran, R.; Raja Kumar, S. Fatty acids-carotenoid complex: An effective anti-TB agent from the chlorella growth factor-extracted spent biomass of Chlorella vulgaris. J. Ethnopharmacol. 2020, 249, 112392. [Google Scholar] [CrossRef]
- Lauritano, C.; Martín, J.; de la Cruz, M.; Reyes, F.; Romano, G.; Ianora, A. First identification of marine diatoms with anti-tuberculosis activity. Sci. Rep. 2018, 8, 2284. [Google Scholar] [CrossRef]
- Ramos, D.F.; Halicki, P.C.B.; Caprara, C.d.S.C.; Borges, P.; M.D’Oca, C.d.R.; Santos, M.d.F.C.; D’Oca, M.G.M.; Roselet, F.; da Silva, P.E.A.; Abreu, P.C. Chemical Profile and Antimicrobial Activity of the Marine Diatom Chaetoceros muelleri. Chem. Biodivers 2022, 19, e202100846. [Google Scholar] [CrossRef]
- Viazau, Y.V.; Manankina, E.E.; Filipchik, E.A.; Goncharik, R.G.; Shalygo, N.V. Effectivness of repeated usage of the modified Zarrouk culture medium for cultivation of Spirulina platensis. Proc. Natl. Acad. Sci. Belarus Biol. Ser. 2018, 63, 426–436. [Google Scholar]
- Favas, R.; Morone, J.; Martins, R.; Vasconcelos, V.; Lopes, G. Cyanobacteria Secondary Metabolites as Biotechnological Ingredients in Natural Anti-Aging Cosmetics: Potential to Overcome Hyperpigmentation, Loss of Skin Density and UV Radiation-Deleterious Effects. Mar. Drugs 2022, 20, 183. [Google Scholar] [CrossRef]
- Vahdati, S.N.; Behboudi, H.; Tavakoli, S.; Aminian, F.; Ranjbar, R. Antimicrobial Potential of the Green Microalgae Isolated from the Persian Gulf. Iran. J. Public Health 2022, 51, 1134–1142. [Google Scholar] [CrossRef]
- Al Naim, H.M.; El Semary, N. Laser Treatment Increases the Antimicrobial Efficacy of Cyanobacterial Extracts against Staphylococcusaureus (SA) and Methicillin-resistantStaphylococcus aureu (MRSA). Int. J. Environ. Res. Public Health 2022, 19, 13305. [Google Scholar] [CrossRef]
- Stěpánková, T.; Ambrožová, L.; Bláha, L.; Giesy, J.P.; Hilscherová, K. In vitro modulation of intracellular receptor signaling and cytotoxicity induced by extracts of cyanobacteria, complex water blooms and their fractions. Aquat. Toxicol. 2011, 105, 497–507. [Google Scholar] [CrossRef]
- Abdullin, S.R.; Nikulin, V.Y.; Nikulin, A.Y.; Manyakhin, A.Y.; Bagmet, V.B.; Suprun, A.R.; Gontcharov, A.A. Roholtiella mixta sp. nov. (Nostocales, Cyanobacteria): Morphology, molecular phylogeny, and carotenoids content. Phycologia 2021, 60, 73–82. [Google Scholar] [CrossRef]
- Erdoğan, A.; Karataş, A.B.; Demir, D.; Demirel, Z.; Aktürk, M.; Çopur, Ö.; Conk-Dalay, M. Manipulation in Culture Conditions of Nanofrustulum shiloi for Enhanced Fucoxanthin Production and Isolation by Preparative Chromatography. Molecules 2023, 28, 1988. [Google Scholar] [CrossRef]
- Guzmán, F.; Wong, G.; Romá, T.; Cárdenas, C.; Alvárez, C.; Schmitt, P.; Albericio, F.; Rojas, V. Identification of Antimicrobial Peptides from the Microalgae Tetraselmis suecica (Kylin) Butcher and Bactericidal Activity Improvement. Mar. Drugs 2019, 17, 453. [Google Scholar] [CrossRef]
- Han, S.; Zhao, J.; Liu, P.; Wang, K.; Qin, S.; Zhao, Z.; Cui, Y. Two Foreign Antimicrobial Peptides Expressed in the Chloroplast of Porphyridium purpureum Possessed Antibacterial Properties. Mar. Drugs 2022, 20, 484. [Google Scholar] [CrossRef]
- Rojas, V.; Rivas, L.; Cárdenas, C.; Guzmán, F. Cyanobacteria and Eukaryotic Microalgae as Emerging Sources of Antibacterial Peptides. Molecules 2020, 25, 5804. [Google Scholar] [CrossRef]
- Pina-Pérez, M.C.; Rivas, A.; Martínez, A.; Rodrigo, D. Antimicrobial potential of macro and microalgae against pathogenic and spoilage microorganisms in food. Food Chem. 2017, 235, 34–44. [Google Scholar] [CrossRef]
- Shaima, A.F.; Mohd Yasin, N.H.; Ibrahim, N.; Takriff, M.S.; Gunasekaran, D.; Ismaeel, M.Y.Y. Unveiling antimicrobial activity of microalgae Chlorella sorokiniana (UKM2), Chlorella sp. (UKM8) and Scenedesmus sp. (UKM9). Saudi J. Biol. Sci. 2022, 29, 1043–1052. [Google Scholar] [CrossRef]
- Dantas, D.M.M.; Oliveira, C.Y.B.; Costa, R.M.P.B.; Carneiro-da-Cunha, M.D.G.; Gálvez, A.O.; Bezerra, R.S. Evaluation of antioxidant and antibacterial capacity of green microalgae Scenedesmus subspicatus. Food Sci. Technol. Int. 2019, 25, 318–326. [Google Scholar] [CrossRef]
- Stirk, W.A.; van Staden, J. Bioprospecting for bioactive compounds in microalgae: Antimicrobial compounds. Biotechnol. Adv. 2022, 59, 107977. [Google Scholar] [CrossRef]
- Alsenani, F.; Tupally, K.R.; Chua, E.T.; Eltanahy, E.; Alsufyani, H.; Parekh, H.S.; Schenk, P.M. Evaluation of microalgae and cyanobacteria as potential sources of antimicrobial compounds. Saudi Pharm. J. 2020, 28, 1834–1841. [Google Scholar] [CrossRef]
- Vornoli, A.; Grande, T.; Lubrano, V.; Vizzarri, F.; Gorelli, C.; Raffaelli, A.; Della Croce, C.M.; Baca, S.Z.; Sandoval, C.; Longo, V.; et al. In Vitro Characterization of Antioxidant, Antibacterial and Antimutagenic Activities of the Green Microalga Ettlia pseudoalveolaris. Antioxidants 2023, 12, 1308. [Google Scholar] [CrossRef]
- Martínez, K.A.; Lauritano, C.; Druka, D.; Romano, G.; Grohmann, T.; Jaspars, M.; Martín, J.; Díaz, C.; Cautain, B.; de la Cruz, M.; et al. Amphidinol 22, a New Cytotoxic and Antifungal Amphidinol from the Dinoflagellate Amphidinium carterae. Mar. Drugs 2019, 17, 385. [Google Scholar] [CrossRef]
- Barone, M.E.; Murphy, E.; Parkes, R.; Fleming, G.T.A.; Campanile, F.; Thomas, O.P.; Touzet, N. Antibacterial Activity and Amphidinol Profiling of the Marine Dinoflagellate Amphidinium carterae (Subclade III). Int. J. Mol. Sci. 2021, 22, 12196. [Google Scholar] [CrossRef]
- Sukhikh, S.; Prosekov, A.; Ivanova, S.; Maslennikov, P.; Andreeva, A.; Budenkova, E.; Kashirskikh, E.; Tcibulnikova, A.; Zemliakova, E.; Samusev, I.; et al. Identification of Metabolites with Antibacterial Activities by Analyzing the FTIR Spectra of Microalgae. Life 2022, 12, 1395. [Google Scholar] [CrossRef]
- Dolganyuk, V.; Andreeva, A.; Sukhikh, S.; Kashirskikh, E.; Prosekov, A.; Ivanova, S.; Michaud, P.; Babich, O. Study of the Physicochemical and Biological Properties of the Lipid Complex of Marine Microalgae Isolated from the Coastal Areas of the Eastern Water Area of the Baltic Sea. Molecules 2022, 27, 5871. [Google Scholar] [CrossRef]
- Šudomová, M.; Shariati, M.A.; Echeverría, J.; Berindan-Neagoe, I.; Nabavi, S.M.; Hassan, S.T.S. A Microbiological, Toxicological, and Biochemical Study of the Effects of Fucoxanthin, a Marine Carotenoid, on Mycobacterium tuberculosis and the Enzymes Implicated in Its Cell Wall: A Link between Mycobacterial Infection and Autoimmune Diseases. Mar. Drugs 2019, 17, 641. [Google Scholar] [CrossRef]
- Liu, B.; Eltanahy, E.E.; Liu, H.; Chua, E.T.; Thomas-Hall, S.R.; Wass, T.J.; Pan, K.; Schenk, P.M. Growth-promoting bacteria double eicosapentaenoic acid yield in microalgae. Bioresour Technol. 2020, 316, 123916. [Google Scholar] [CrossRef]
- Dao, G.; Wang, S.; Wang, X.; Chen, Z.; Wu, Y.; Wu, G.; Lu, Y.; Liu, S.; Hu, H. Enhanced Scenedesmus sp. growth in response to gibberellin secretion by symbiotic bacteria. Sci. Total Environ. 2020, 740, 140099. [Google Scholar] [CrossRef]
- Iida, H.; Aburai, N.; Fujii, K. Microalga-bacteria Community with High Level Carbon Dioxide Acclimation and Nitrogen-fixing Ability. Protist 2023, 174, 125957. [Google Scholar] [CrossRef]
- Astafyeva, Y.; Gurschke, M.; Streit, W.R.; Krohn, I. Interplay between the microalgae Micrasterias radians and its symbiont Dyadobacter sp. HH091. Front. Microbiol. 2022, 13, 1006609. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar]
- Lowry, O.H.; Rosenbrough, N.J.; Faar, A.L.; Randall, R.J. Protein measurment with folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related sunstances. Anakytic Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Ryabusshko, L.I.; Zheleznova, S.N.; Nekhoroshev, M.V. Effect of nitrogen on fucoxanthin accumulation in the diatom Cylindrotheca closterium (Ehrenb.) Reimann et Lewin. Int. J. Algae 2017, 19, 79–84. [Google Scholar] [CrossRef]
- Hashimoto, T.; Ozaki, Y.; Taminato, M.; Dass, S.K.; Mizuno, M.; Yoshimura, K.; Maoka, T.; Kanazawa, K. The distribution and accumulation of fucoxanthin and its metabolites. Br. J. Nutr. 2009, 102, 242–248. [Google Scholar] [CrossRef]
- Bennett, A.; Bogorad, L. Complementary chromatic adaptation in a filamentous blue-green alga. J. Cell Biol. 1973, 58, 419–435. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content, mg/g of Dry Biomass | L. ectocarpi | N. shiloi | R. mixta sp. nov. | P. agardhii | T. viridis |
|---|---|---|---|---|---|
| Proteins | 560 ± 5 | 450 ± 5 | 680 ± 10 | 600 ± 10 | 350 ± 10 |
| Lipids | 250 ± 5 | 280 ± 5 | 50 ± 5 | 100 ± 5 | 195 ± 6 |
| Carbohydrates | 50 ± 3 | 70 ± 3 | 110 ± 5 | 180 ± 5 | 340 ± 5 |
| FX | ND | 15 ± 0.5 | ND | ND | ND |
| C-phycoerythrin | 78.8 ± 0.8 | ND | 35 ± 0.8 | 20 ± 1.2 | ND |
| C-phycocyanin | 22.0 ± 0.8 | ND | 140 ± 1 | 88.8 ± 1 | ND |
| PUFA, include: | 50 ± 1.2 | 67 ± 1.25 | 10 ±1.1 | 11 ± 1.2 | 70 ± 1.25 |
| Eicosapentaenoic acid (C20:5 ω3) | ND | 47.3 ± 0.61 | ND | ND | 4.8 ± 0.7 |
| Arachidonic acid (C20:4ω6) | ND | 8.5 ± 0.31 | ND | ND | 0.6 ± 0.6 |
| γ-Linolenic acid (C18:3 n 6) | 18 ± 0.04 | 0.22 ± 0.01 | 2.75 ± 0.02 | 1.5 ± 0.01 | 10.7 ± 0.05 |
| Linoleic acid (C18:2 n 6) | 25 ± 0.05 | 1.455 ± 0.04 | 3.16 ± 0.02 | 2.1 ± 0.01 | 2.5 ± 0.02 |
| Oleic acid (C18:1 (n 9) | 0.1 ± 0.02 | 1.46 ± 0.01 | ND | 2 ± 0.01 | 0,1 ± 0.02 |
| α-Linolenic acid (C18:3 n 3) | 10.0 ± 0.05 | ND | ND | ND | 6.5 ± 0.05 |
| Parameters | K. pneumoniae Clinical Isolate (s) | K. pneumoniae Clinical Isolate (r) | MSSA | MRSA | S. aureus Clinical Isolate (r) | A. baumannii Clinical Isolate (r) | E. faecalis Clinical Isolate (r) | P. aeruginosa Clinical Isolate (s) | P. aeruginosa Clinical Isolate (r) | S. pyogenes Clinical Isolate (s) |
|---|---|---|---|---|---|---|---|---|---|---|
| On Day 14 | ||||||||||
| Control | 1.18 ± 0.08 | 1.75 ± 0.2 | 1.1 ± 0.09 | 1.47 ± 0.08 | 1.46 ± 0.16 | 0.98 ± 0.17 | 1.08 ± 0.09 | 1.94 ± 0.33 | 1.37 ± 0.33 | 1.88 ± 0.32 |
| Cef | 0.82 ± 0.08 * | 1.24 ± 0.04 * | 1.03 ± 0.14 | 1.32 ± 0.1 * | 1.49 ± 0.13 | 0.7 ± 0.1 * | 2.1 ± 0.32 | 1.29 ± 0.03 * | 1.3 ± 0.03 | 1.53 ± 0.27 |
| A. platensis | 0.6 ± 0.17 * | 0.8 ± 0.05 *# | 0.83 ± 0.01 *# | 0.96 ± 0.11 *# | 0.96 ± 0.04 *# | 0.68 ± 0.11 * | 1.15 ± 0.13 # | 0.49 ± 0.15 *# | 0.67 ± 0.03 *# | 1.3 ± 0.07 * |
| FX | 0.83 ± 0.1 * | 0.66 ± 0.05 *# | 0.72 ± 0.08 *# | 0.46 ± 0.07 *# | 1.16 ± 0.05 *# | 0.68 ± 0.14 | 2 ± 0.26 | 0.5 ± 0.06 *# | 0.66 ± 0.07 *# | 2.07 ± 0.29 # |
| On Day 21 | ||||||||||
| Control | 2.52 ± 0.9 | 3.66 ± 0.39 | 4 ± 1 | 4 ± 0.9 | 4 ± 1 | 3.09 ± 1 | 4 ± 1 | 4 ± 0.8 | 4 ± 1 | 4 ± 0.9 |
| Cef | 2.15 ± 0.9 | 3.53 ± 0.54 | 2.07 ± 0.45 * | 3.28 ± 0.96 | 1.23 ± 1.03 * | 4 ± 0.9 | 4 ± 0.8 | 3.75 ± 0.51 | 3.23 ± 0.97 | 2.39 ± 1.67 |
| A. platensis | 0.54 ± 0.24 *# | 2.33 ± 0.1 *# | 1.98 ± 0.43 * | 2.07 ± 0.51 * | 2 ± 0.65 * | 0.85 ± 0.29 *# | 2.01 ± 0.65 *# | 1.99 ± 0.69 *# | 1.89 ± 0.79 * | 2 ± 0.65 * |
| FX | 1.11 ± 0.03 *# | 2.75 ± 0.2 *# | 2.26 ± 0.62 * | 2.16 ± 0.92 | 2.68 ± 0.34 # | 1.06 ± 0.01 *# | 2.6 ± 0.34 *# | 2.85 ± 0.2 *# | 2.89 ± 0.1 * | 2.2 ± 0.5 * |
| Parameters | N. shiloi | T. viridis | Pl. agardhii | L. ectocarpi | R. mixta sp. nov. | FX |
|---|---|---|---|---|---|---|
| K. pneumonia clinical isolate (s) [Control = 1.38 ± 0.01, Ceftazidime = 0.92 ± 0.01] | ||||||
| 1% v/v | 1.79 ± 0.22 | 1.7 ± 0.04 | 2.18 ± 0.01 | 2.06 ± 0.04 | 2.05 ± 0.04 | 1.4 ± 0.01 |
| 0.5% v/v | 2.11 ± 0.08 | 2.01 ± 0.02 | 2.33 ± 0.02 | 1.91 ± 0.07 | 1,74 ± 0.05 | 1.35 ± 0.01 |
| 0.25% v/v | 1.72 ± 0.01 | 1.75 ± 0.01 | 2.11 ± 0.08 | 1.8 ± 0.07 | 1.29 ± 0.01 * | 1.18 ± 0.01 * |
| K. pneumonia clinical isolate (r) [Control = 1.83 ± 0.02, Ceftazidime = 1.72 ± 0.01] | ||||||
| 1% v/v | 1.3 ± 0.01 *# | 1.76 ± 0.01 * | 1.7 ± 0.01 | 1.46 ± 0.01 | 1.36 ± 0.02 *# | 1.18 ± 0.01 *# |
| 0.5% v/v | 1.19 ± 0.01 *# | 1.68 ± 0. 01 * | 1.68 ± 0.01 * | 1.24 ± 0.02 *# | 1.23 ± 0.01 *# | 0.83 ± 0.01 *# |
| 0.25% v/v | 1.22 ± 0.01 *# | 1.47 ± 0.01 *# | 1.1 ± 0.01 *# | 1.04 ± 0.03 *# | 0.97 ± 0.01 *# | 1.19 ± 0.01 *# |
| MSSA [Control = 1.92 ± 0.04, Ceftazidime = 0.43 ± 0.01] | ||||||
| 1% v/v | 1.45 ± 0.01 * | 1.19 ± 0.01 * | 2 ± 0.01 | 1.53 ± 0.01 * | 1.47 ± 0.01 * | 1.61 ± 0.01 * |
| 0.5% v/v | 1.08 ± 0.01 * | 1.27 ± 0.01 * | 1.47 ± 0.01 * | 1.69 ± 0.0 * | 1.49 ± 0.01 * | 1.53 ± 0.01 * |
| 0.25% v/v | 1.34 ± 0.01 * | 1.43 ± 0.01 * | 1.51 ± 0.01 * | 1.38 ± 0.01 * | 2.01 ± 0.01 | 1.19 ± 0.01 * |
| MRSA [Control = 2.11 ± 0.07, Ceftazidime = 2.34 ± 0.01] | ||||||
| 1% v/v | 1.52 ± 0.03 *# | 1.41 ± 0.01 *# | 2.01 ± 0.02 | 1.92 ± 0.1 *# | 2.16 ± 0.03 | 1.35 ± 0.01 *# |
| 0.5% v/v | 1.58 ± 0.01 *# | 1.08 ± 0.01 *# | 1.92 ± 0.01 *# | 1.8 ± 0.01 *# | 1.83 ± 0.01 *# | 1.25 ± 0.01 *# |
| 0.25% v/v | 1.59 ± 0.03 *# | 1.46 ± 0.01 *# | 1.31 ± 0.01 *# | 1.58 ± 0.01 *# | 1.98 ± 0.01 *# | 0.55 ± 0.01 *# |
| A. baumannii clinical isolate (r) [Control = 2.09 ± 0.03, Ceftazidime = 0.92 ± 0.01] | ||||||
| 1% v/v | 2.92 ± 0.01 | 2.39 ± 0.02 | 3.62 ± 0.08 | 2.28 ± 0.01 | 2.23 ± 0.01 | 1.43 ± 0.02 * |
| 0.5% v/v | 2.75 ± 0.01 | 2.52 ± 0.12 | 2.68 ± 0.02 | 2.33 ± 0.04 | 2.02 ± 0.18 | 1.18 ± 0.24 * |
| 0.25% v/v | 2.15 ± 0.01 | 2.53 ± 0.06 | 2.64 ± 0.05 | 2.18 ± 0.01 | 2.03 ± 0.01 | 0.9 ± 0.1 * |
| E. faecalis clinical isolate (r) [Control = 2.61 ± 0.03, Ceftazidime = 0.92 ± 0.03] | ||||||
| 1% v/v | 1.67 ± 0.01 * | 1.54 ± 0.01 * | 2.11 ± 0.01 * | 2.16 ± 0.02 * | 2.46 ± 0.02 * | 1.08 ± 0.01 * |
| 0.5% v/v | 1.64 ± 0.01 * | 1.82 ± 0.01 * | 2 ± 0.01 * | 2.18 ± 0.01 * | 2.4 ± 0.01 * | 1.01 ± 0.01 * |
| 0.25% v/v | 1.57 ± 0.01 * | 1.56 ± 0. 01 * | 1.38 ± 0.01 * | 1.69 ± 0.03 * | 1.61 ± 0.01 * | 0.59 ± 0.01 *# |
| P. aeruginosa clinical isolate (s) [Control = 1.77 ± 0.01, Ceftazidime = 1.4 ± 0.05] | ||||||
| 1% v/v | 2.8 ± 0.04 | 1.65 ± 0.01 * | 2.83 ± 0.02 | 1.84 ± 0.01 | 1.55 ± 0.01 * | 0.78 ± 0,01 *# |
| 0.5% v/v | 1.72 ± 0.01 | 1.45 ± 0.01 * | 1.81 ± 0.01 | 1.68 ± 0.01 | 1.55 ± 0.01 * | 0.55 ± 0.01 *# |
| 0.25% v/v | 1.5 ± 0.01 | 1.41 ± 0.01 * | 1.47 ± 0.01 * | 1.36 ± 0.01 * | 1.59 ± 0.02 * | 0.44 ± 0.01 # |
| P. aeruginosa clinical isolate (r) [Control = 1.73 ± 0.01, Ceftazidime = 1.25 ± 0.01] | ||||||
| 1% v/v | 2.33 ± 0.02 | 1.35 ± 0.01 * | 2.28 ± 0.02 | 1.46 ± 0.01 * | 0.92 ± 0.01 *# | 0.88 ± 0.01 *# |
| 0.5% v/v | 1.9 ± 0.01 | 2.13 ± 0.01 | 1,71 ± 0.01 | 1.36 ± 0.03 * | 0.75 ± 0.01 *# | 0.7 ± 0.01 *# |
| 0.25% v/v | 1.64 ± 0.01 * | 1.45 ± 0.02 * | 1.63 ± 0.01 | 1.19 ± 0.03 * | 0.67 ± 0.01 *# | 0.43 ± 0.01 *# |
| S. aureus clinical isolate (r) [Control = 0.98 ± 0.01, Ceftazidime = 0.99 ± 0.01] | ||||||
| 1% v/v | 0.66 ± 0.01 * | 0.84 ± 0.01 * | 1.05 ± 0.01 | 0.91 ± 0.01 * | 0.99 ± 0.03 | 0.47 ± 0.04 *# |
| 0.5% v/v | 0.53 ± 0.06 * | 0.79 ± 0.03 * | 0.95 ± 0.08 | 0.28 ± 0.02 *# | 0.68 ± 0.02 *# | 0.54 ± 0.04 *# |
| 0.25% v/v | 1.55 ± 0.07 | 0.36 ± 0.05 * | 1.14 ± 0.06 | 0.99 ± 0.09 | 0.78 ± 0.08 *# | 0.5 ± 0.02 *# |
| Control | Ceftazidime | Chlorophyllipt | N. shiloi | T. viridis | P. agardhii | L. ectocarpi | R.mixta sp. nov. | FX |
|---|---|---|---|---|---|---|---|---|
| K. pneumoniae clinical isolate (r) | ||||||||
| 2.27 ± 0.05 | 0.78 ± 0.01 | 0.98 ± 0.01 * | 1.85 ± 0.01 * | 1.53 ± 0.01 * | 1.78 ± 0.01 * | 1.93 ± 0.04 * | 1.7 ± 0.01 * | 0.77 ± 0.01 *& |
| MRSA | ||||||||
| 3.02 ± 0.03 | 2.44 ± 0.01 | 0.72 ± 0.0 *# | 0.9 ± 0.01 *# | 1.2 ± 0.01 *# | 1.93 ± 0.02 *# | 0.93 ± 0.01 *# | 0.9 ± 0.01 *# | 0.6 ± 0.01 *#& |
| A. bauman ii clinical isolate (r) | ||||||||
| 3.49 ± 0.03 | 0.92 ± 0.01 | 1.04 ± 0.01 * | 1.6 ± 0.02 * | 1.88 ± 0.04 * | 1.86 ± 0.0 * | 2.27 ± 0.02 * | 1.53 ± 0.01 * | 0.8 ± 0.01 *# |
| E. faecalis clinical isolate (r) | ||||||||
| 2.29 ± 0.02 | 0.92 ± 0.01 | 0.57 ± 0.01 *# | 1.41 ± 0.01 * | 1.78 ± 0.01 * | 1.82 ± 0.26 * | 2.06 ± 0.01 * | 1.7 ± 0.0 * | 2.03 ± 0.02 * |
| P. aeruginosa clinical isolate (r) | ||||||||
| 1.08 ± 0.01 | 0.93 ± 0.01 | 0.67 ± 0.01 *# | 0.49 ± 0.01 *#& | 1.23 ± 0.02 | 2.18 ± 0.03 | 2.54 ± 0.03 | 0.74 ± 0.001 *# | 0.66 ± 0.01 *# |
| Parameters | N. shiloi | T. viridis | P. agardhii | L. ectocarpi | R. mixta sp. nov. | FX |
|---|---|---|---|---|---|---|
| Mycobacterium tuberculosis strain H37Rv [Control = 1.38 ± 0.01, Rifampicin 100 µg/mL = 0.96 ± 0.04] | ||||||
| 1% v/v | 1.74 ± 0.01 | 1.72 ± 0.01 | 1.33 ± 0.01 * | 1.09 ± 0.01 * | 0.62 ± 0.01 *# | 1.18 ± 0.01 * |
| 0.5% v/v | 0.97 ± 0.01 * | 1.41 ± 0.01 | 1.06 ± 0.01 * | 0.89 ± 0.01 *# | 0.79 ± 0.01 *# | 0.73 ± 0.01 *# |
| 0.25% v/v | 0.67 ± 0.01 *# | 1.18 ± 0.01 | 0.77 ± 0.01 *# | 0.57 ± 0.01 *# | 1.47 ± 0.01 | 2.06 ± 0.01 |
| Mycobacterium smegmatis [Control = 1.82 ± 0.02, Rifampicin = 1.6 ± 0.02] | ||||||
| 1% v/v | 1.91 ± 0.01 | 1.08 ± 0.01 *# | 2.76 ± 0.01 | 1.12 ± 0.01 *# | 0.99 ± 0.01 *# | 1.43 ± 0.01 *# |
| 0.5% v/v | 0.96 ± 0.01 *# | 2.01 ± 0.01 | 0.93 ± 0.01 *# | 1.01 ± 0.01 *# | 0.76 ± 0.01 *# | 1.13 ± 0.01 *# |
| 0.25% v/v | 1.46 ± 0.01 *# | 1.94 ± 0.01 | 0.84 ± 0.01 *# | 0.54 ± 0.01 *# | 1.57 ± 0.01 *# | 0.99 ± 0.01 *# |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lykov, A.; Salmin, A.; Gevorgiz, R.; Zheleznova, S.; Rachkovskaya, L.; Surovtseva, M.; Poveshchenko, O. Study of the Antimicrobial Potential of the Arthrospira platensis, Planktothrix agardhii, Leptolyngbya cf. ectocarpi, Roholtiella mixta nov., Tetraselmis viridis, and Nanofrustulum shiloi against Gram-Positive, Gram-Negative Bacteria, and Mycobacteria. Mar. Drugs 2023, 21, 492. https://doi.org/10.3390/md21090492
Lykov A, Salmin A, Gevorgiz R, Zheleznova S, Rachkovskaya L, Surovtseva M, Poveshchenko O. Study of the Antimicrobial Potential of the Arthrospira platensis, Planktothrix agardhii, Leptolyngbya cf. ectocarpi, Roholtiella mixta nov., Tetraselmis viridis, and Nanofrustulum shiloi against Gram-Positive, Gram-Negative Bacteria, and Mycobacteria. Marine Drugs. 2023; 21(9):492. https://doi.org/10.3390/md21090492
Chicago/Turabian StyleLykov, Alexander, Alexei Salmin, Ruslan Gevorgiz, Svetlana Zheleznova, Lyubov Rachkovskaya, Maria Surovtseva, and Olga Poveshchenko. 2023. "Study of the Antimicrobial Potential of the Arthrospira platensis, Planktothrix agardhii, Leptolyngbya cf. ectocarpi, Roholtiella mixta nov., Tetraselmis viridis, and Nanofrustulum shiloi against Gram-Positive, Gram-Negative Bacteria, and Mycobacteria" Marine Drugs 21, no. 9: 492. https://doi.org/10.3390/md21090492
APA StyleLykov, A., Salmin, A., Gevorgiz, R., Zheleznova, S., Rachkovskaya, L., Surovtseva, M., & Poveshchenko, O. (2023). Study of the Antimicrobial Potential of the Arthrospira platensis, Planktothrix agardhii, Leptolyngbya cf. ectocarpi, Roholtiella mixta nov., Tetraselmis viridis, and Nanofrustulum shiloi against Gram-Positive, Gram-Negative Bacteria, and Mycobacteria. Marine Drugs, 21(9), 492. https://doi.org/10.3390/md21090492