
Karenia brevis Extract Induces Cellular Entry through Distinct Mechanisms in Phagocytic RAW 264.7 Macrophages versus Non-Phagocytic Vero Cells

 , , ,
, , ,
Abstract
:
{kind=link}
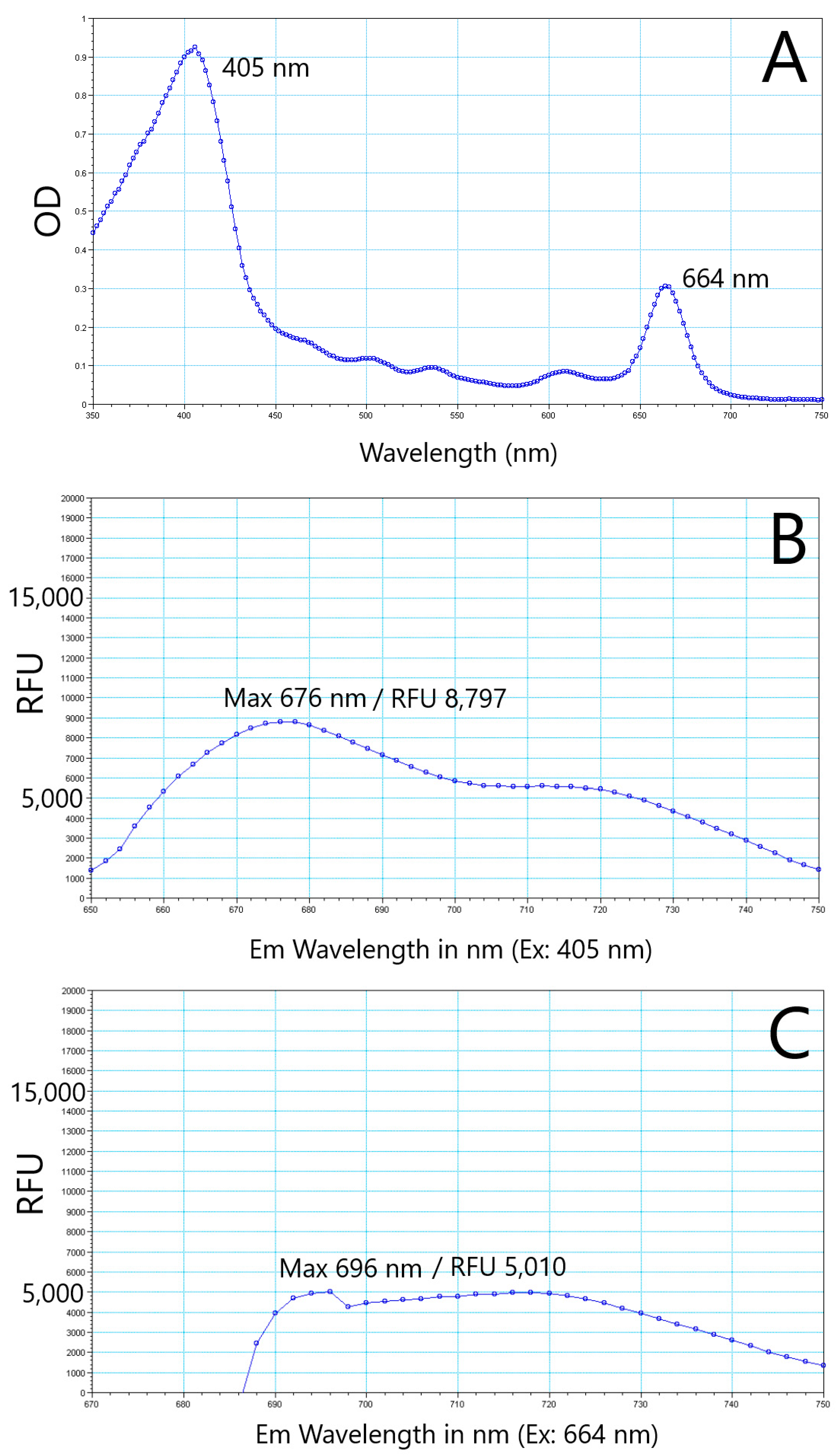{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. K. brevis Algal Extracts Absorb and Emit Fluorescence
2.2. Fluorescent K. brevis Extracts Contain Both Chlorophyll α and Pheophytin α
2.3. Assessment Fluorescence in Phagocytic Macrophages and Non-Phagocytic Vero Cells Using Flow Cytometry
2.4. Fluorescence Accumulates through Distinct Mechanisms in RAW 264.7 and Vero Cells
2.5. K. brevis Extract Downregulated RAW 264.7 Expression of CD206, in the Absence of Changes in Cytokine Secretion
3. Discussion
4. Materials and Methods
4.1. Algal Extracts
4.2. Cell Culture
4.3. Flow Cytometry Analysis
4.4. Spectral Analysis
4.5. Fluorescence Microscopy
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Poli, M.A.; Mende, T.J.; Baden, D.G. Brevetoxins, Unique Activators of Voltage-Sensitive Sodium Channels, Bind to Specific Sites in Rat Brain Synaptosomes. Mol. Pharmacol. 1986, 30, 129–135. [Google Scholar] [PubMed]
- Gannon, D.; Berens McCabe, E.; Camilleri, S.; Gannon, J.; Brueggen, M.; Barleycorn, A.; Palubok, V.; Kirkpatrick, G.; Wells, R. Effects of Karenia brevis Harmful Algal Blooms on Nearshore Fish Communities in Southwest Florida. Mar. Ecol. Prog. Ser. 2012, 378, 171–186. [Google Scholar] [CrossRef]
- Fleming, L.E.; Kirkpatrick, B.; Backer, L.C.; Walsh, C.J.; Nierenberg, K.; Clark, J.; Reich, A.; Hollenbeck, J.; Benson, J.; Cheng, Y.S.; et al. Review of Florida Red Tide and Human Health Effects. Harmful Algae 2011, 10, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Tilney, C.L.; Shankar, S.; Hubbard, K.A.; Corcoran, A.A. Is Karenia brevis Really a Low-Light-Adapted Species? Harmful Algae 2019, 90, 101709. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, W.G.; Kang, Y.; Zika, R.G. Resolving DOM Fluorescence Fractions during a Karenia brevis Bloom Patch on the Southwest Florida Shelf. Cont. Shelf Res. 2012, 32, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.J.; Arrington, D.A.; Porter, D.; Lovko, V. Documenting the Duration and Chlorophyll Pigments of an Allochthonous Karenia brevis Bloom in the Loxahatchee River Estuary (LRE), Florida. Harmful Algae 2020, 97, 101851. [Google Scholar] [CrossRef] [PubMed]
- Colman, J.R.; Ramsdell, J.S. The Type B Brevetoxin (PbTx-3) Adversely Affects Development, Cardiovascular Function, and Survival in Medaka (Oryzias latipes) Embryos. Environ. Health Perspect. 2003, 111, 1920–1925. [Google Scholar] [CrossRef]
- Abraham, W.M.; Bourdelais, A.J.; Sabater, J.R.; Ahmed, A.; Lee, T.A.; Serebriakov, I.; Baden, D.G. Airway Responses to Aerosolized Brevetoxins in an Animal Model of Asthma. Am. J. Respir. Crit. Care Med. 2005, 171, 26–34. [Google Scholar] [CrossRef]
- Pesek, J.J.; Matyska, M.T.; Hiltz, T.; McCall, J. Application of a Cholesterol-Based Stationary Phase for the Analysis of Brevetoxins. J. Sep. Sci. 2023, 46, 2200666. [Google Scholar] [CrossRef]
- Finol-Urdaneta, R.K.; Zhorov, B.S.; Baden, D.G.; Adams, D.J. Brevetoxin versus Brevenal Modulation of Human Nav1 Channels. Mar. Drugs 2023, 21, 396. [Google Scholar] [CrossRef]
- Hort, V.; Abadie, E.; Arnich, N.; Dechraoui Bottein, M.-Y.; Amzil, Z. Chemodiversity of Brevetoxins and Other Potentially Toxic Metabolites Produced by Karenia Spp. and Their Metabolic Products in Marine Organisms. Mar. Drugs 2021, 19, 656. [Google Scholar] [CrossRef] [PubMed]
- Keeler, D.M.; Grandal, M.K.; McCall, J.R. Brevenal, a Marine Natural Product, Is Anti-Inflammatory and an Immunomodulator of Macrophage and Lung Epithelial Cells. Mar. Drugs 2019, 17, 184. [Google Scholar] [CrossRef] [PubMed]
- McCall, J.R.; Sausman, K.T.; Keeler, D.M.; Brown, A.P.; Turrise, S.L. Immune Modulating Brevetoxins: Monocyte Cytotoxicity, Apoptosis, and Activation of M1/M2 Response Elements Is Dependent on Reactive Groups. Mar. Drugs 2022, 20, 233. [Google Scholar] [CrossRef] [PubMed]
- Jeglitsch, G.; Rein, K.; Baden, D.G.; Adams, D.J. Brevetoxin-3 (PbTx-3) and Its Derivatives Modulate Single Tetrodotoxin-Sensitive Sodium Channels in Rat Sensory Neurons. J. Pharmacol. Exp. Ther. 1998, 284, 516–525. [Google Scholar] [PubMed]
- Chen, W.; Tuladhar, A.; Rolle, S.; Lai, Y.; del Rey, F.R.; Zavala, C.E.; Liu, Y.; Rein, K.S. Brevetoxin-2, Is a Unique Inhibitor of the C-Terminal Redox Center of Mammalian Thioredoxin Reductase-1. Toxicol. Appl. Pharmacol. 2017, 329, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Tuladhar, A.; Hondal, R.J.; Colon, R.; Hernandez, E.L.; Rein, K.S. Effectors of Thioredoxin Reductase: Brevetoxins and Manumycin-A. Comp. Biochem. Physiol. Toxicol. Pharmacol. CBP 2019, 217, 76–86. [Google Scholar] [CrossRef]
- Kang, H.; Lee, C.H.; Kim, J.R.; Kwon, J.Y.; Seo, S.G.; Han, J.G.; Kim, B.G.; Kim, J.-E.; Lee, K.W. Chlorella vulgaris Attenuates Dermatophagoides Farinae-Induced Atopic Dermatitis-Like Symptoms in NC/Nga Mice. Int. J. Mol. Sci. 2015, 16, 21021–21034. [Google Scholar] [CrossRef]
- Konishi, F.; Tanaka, K.; Himeno, K.; Taniguchi, K.; Nomoto, K. Antitumor Effect Induced by a Hot Water Extract of Chlorella vulgaris (CE): Resistance to Meth-A Tumor Growth Mediated by CE-Induced Polymorphonuclear Leukocytes. Cancer Immunol. Immunother. 1985, 19, 73–78. [Google Scholar] [CrossRef]
- Chovancikova, M.; Simek, V. Effects of High-Fat and Chlorella vulgaris Feeding on Changes in Lipid Metabolism in Mice. Biologia 2001, 56, 661–666. [Google Scholar]
- Cherng, J.-Y.; Shih, M.-F. Potential Hypoglycemic Effects of Chlorella in Streptozotocin-Induced Diabetic Mice. Life Sci. 2005, 77, 980–990. [Google Scholar] [CrossRef]
- Kaur, M.; Bhatia, S.; Gupta, U.; Decker, E.; Tak, Y.; Bali, M.; Gupta, V.K.; Dar, R.A.; Bala, S. Microalgal Bioactive Metabolites as Promising Implements in Nutraceuticals and Pharmaceuticals: Inspiring Therapy for Health Benefits. Phytochem. Rev. 2023, 22, 903–933. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Nishizawa, M.; Shimizu, Y.; Saeki, H. Anti-Inflammatory Effects of Dulse (Palmaria palmata) Resulting from the Simultaneous Water-Extraction of Phycobiliproteins and Chlorophyll a. Food Res. Int. 2017, 100, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Edwards, B.S.; Sklar, L.A. Flow Cytometry: Impact on Early Drug Discovery. SLAS Discov. 2015, 20, 689–707. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, K.M. Flow Cytometry: An Overview. Curr. Protoc. Immunol. 2018, 120, 5.1.1–5.1.11. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Kroeger, J.L.; Markowitz, J. Introduction to Multiparametric Flow Cytometry and Analysis of High-Dimensional Data. In Translational Bioinformatics for Therapeutic Development; Methods in Molecular Biology; Humana: New York, NY, USA, 2021; Volume 2194, pp. 239–253. [Google Scholar] [CrossRef]
- Cossarizza, A.; Chang, H.-D.; Radbruch, A.; Acs, A.; Adam, D.; Adam-Klages, S.; Agace, W.W.; Aghaeepour, N.; Akdis, M.; Allez, M.; et al. Guidelines for the Use of Flow Cytometry and Cell Sorting in Immunological Studies (Second Edition). Eur. J. Immunol. 2019, 49, 1457–1973. [Google Scholar] [CrossRef]
- Baell, J.; Walters, M.A. Chemistry: Chemical Con Artists Foil Drug Discovery. Nature 2014, 513, 481–483. [Google Scholar] [CrossRef]
- Capuzzi, S.J.; Muratov, E.N.; Tropsha, A. Phantom PAINS: Problems with the Utility of Alerts for Pan-Assay INterference CompoundS. J. Chem. Inf. Model. 2017, 57, 417–427. [Google Scholar] [CrossRef]
- van der Gronde, T.; Groot, C.A.U.; Pieters, T. Addressing the Challenge of High-Priced Prescription Drugs in the Era of Precision Medicine: A Systematic Review of Drug Life Cycles, Therapeutic Drug Markets and Regulatory Frameworks. PLoS ONE 2017, 12, e0182613. [Google Scholar] [CrossRef]
- DiMasi, J.A.; Grabowski, H.G.; Hansen, R.W. Innovation in the Pharmaceutical Industry: New Estimates of R&D Costs. J. Health Econ. 2016, 47, 20–33. [Google Scholar] [CrossRef]
- Trindade, I.C.; Pound-Lana, G.; Pereira, D.G.S.; de Oliveira, L.A.M.; Andrade, M.S.; Vilela, J.M.C.; Postacchini, B.B.; Mosqueira, V.C.F. Mechanisms of Interaction of Biodegradable Polyester Nanocapsules with Non-Phagocytic Cells. Eur. J. Pharm. Sci. 2018, 124, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Vercammen, M.; Scorza, T.; El Bouhdidi, A.; Van Beeck, K.; Carlier, Y.; Dubremetz, J.F.; Verschueren, H.; El Bouhdidi, A.; Van Beeck, K. Opsonization of Toxoplasma gondii Tachyzoites with Nonspecific Immunoglobulins Promotes Their Phagocytosis by Macrophages and Inhibits Their Proliferation in Nonphagocytic Cells in Tissue Culture. Parasite Immunol. 1999, 21, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 1801. [Google Scholar] [CrossRef] [PubMed]
- Funes, S.C.; Rios, M.; Escobar-Vera, J.; Kalergis, A.M. Implications of Macrophage Polarization in Autoimmunity. Immunology 2018, 154, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J. Macrophage Polarization. Annu. Rev. Physiol. 2017, 79, 541–566. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Martinez, F.O. Alternative Activation of Macrophages: Mechanism and Functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Boutilier, A.J.; Elsawa, S.F. Macrophage Polarization States in the Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 6995. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ma, C.; Gong, L.; Guo, Y.; Fu, K.; Zhang, Y.; Zhou, H.; Li, Y. Macrophage Polarization and Its Role in Liver Disease. Front. Immunol. 2021, 12, 803037. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, S.; Wu, H.; Rong, X.; Guo, J. M2b Macrophage Polarization and Its Roles in Diseases. J. Leukoc. Biol. 2019, 106, 345–358. [Google Scholar] [CrossRef]
- Molecular Devices. FlexStation 3 Multi-Mode Microplate Reader User Guide; Molecular Devices: San Jose, CA, USA, 2018. [Google Scholar]
- Morançais, M.; Mouget, J.-L.; Dumay, J. Chapter 7—Proteins and Pigments. In Microalgae in Health and Disease Prevention; Levine, I.A., Fleurence, J., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 145–175. ISBN 978-0-12-811405-6. [Google Scholar]
- Mackinney, G.; Joslyn, M.A. The Conversion of Chlorophyll to Pheophytin. J. Am. Chem. Soc. 1940, 62, 231–232. [Google Scholar] [CrossRef]
- Petrovic, S.; Zvezdanović, J.; Anđelković, T.; Marković, D. The Identification of Chlorophyll and Its Derivatives in the Pigment Mixtures: HPLC-Chromatography, Visible and Mass Spectroscopy Studies. Savrem. Technol. 2012, 1, 16–24. [Google Scholar]
- French, C.S.; Smith, J.H.C.; Virgin, H.I.; Airth, R.L. Fluorescence-Spectrum Curves of Chlorophylls, Pheophytins, Phycoerythrins, Phycocyanins and Hypericin. Plant Physiol. 1956, 31, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Pathak, C.; Vaidya, F.U.; Waghela, B.N.; Jaiswara, P.K.; Gupta, V.K.; Kumar, A.; Rajendran, B.K.; Ranjan, K. Insights of Endocytosis Signaling in Health and Disease. Int. J. Mol. Sci. 2023, 24, 2971. [Google Scholar] [CrossRef] [PubMed]
- França, A.; Aggarwal, P.; Barsov, E.V.; Kozlov, S.V.; Dobrovolskaia, M.A.; González-Fernández, Á. Macrophage Scavenger Receptor A Mediates the Uptake of Gold Colloids by Macrophages In Vitro. Nanomedicine 2011, 6, 1175–1188. [Google Scholar] [CrossRef]
- Aderem, A.; Underhill, D.M. Mechanisms of Phagocytosis in Macrophages. Annu. Rev. Immunol. 1999, 17, 593–623. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Wei, W.; Yue, Z.; Lv, P.; Wang, L.; Ma, G.; Su, Z. Particle Size Affects the Cellular Response in Macrophages. Eur. J. Pharm. Sci. 2010, 41, 650–657. [Google Scholar] [CrossRef]
- He, C.; Hu, Y.; Yin, L.; Tang, C.; Yin, C. Effects of Particle Size and Surface Charge on Cellular Uptake and Biodistribution of Polymeric Nanoparticles. Biomaterials 2010, 31, 3657–3666. [Google Scholar] [CrossRef]
- Gillespie, E.J.; Ho, C.-L.C.; Balaji, K.; Clemens, D.L.; Deng, G.; Wang, Y.E.; Elsaesser, H.J.; Tamilselvam, B.; Gargi, A.; Dixon, S.D.; et al. Selective Inhibitor of Endosomal Trafficking Pathways Exploited by Multiple Toxins and Viruses. Proc. Natl. Acad. Sci. USA 2013, 110, E4904–E4912. [Google Scholar] [CrossRef]
- Macia, E.; Ehrlich, M.; Massol, R.; Boucrot, E.; Brunner, C.; Kirchhausen, T. Dynasore, a Cell-Permeable Inhibitor of Dynamin. Dev. Cell 2006, 10, 839–850. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.-A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage Plasticity, Polarization, and Function in Health and Disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Liu, L.; Guo, H.; Song, A.; Huang, J.; Zhang, Y.; Jin, S.; Li, S.; Zhang, L.; Yang, C.; Yang, P. Progranulin Inhibits LPS-Induced Macrophage M1 Polarization via NF-κB and MAPK Pathways. BMC Immunol. 2020, 21, 32. [Google Scholar] [CrossRef] [PubMed]
- Aki, T.; Funakoshi, T.; Noritake, K.; Unuma, K.; Uemura, K. Extracellular Glucose Is Crucially Involved in the Fate Decision of LPS-Stimulated RAW264.7 Murine Macrophage Cells. Sci. Rep. 2020, 10, 10581. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.-S.; Xiang, X.-W.; Jin, H.-X.; Guo, X.-Y.; Liu, L.-J.; Huang, Y.-N.; OuYang, X.-K.; Qu, Y.-L. Composition and Anti-Inflammatory Effect of Polysaccharides from Sargassum Horneri in RAW264.7 Macrophages. Int. J. Biol. Macromol. 2016, 88, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Yim, M.-J.; Lee, J.M.; Choi, G.; Lee, D.-S.; Park, W.S.; Jung, W.-K.; Park, S.; Seo, S.-K.; Park, J.; Choi, I.-W.; et al. Anti-Inflammatory Potential of Carpomitra costata Ethanolic Extracts via Inhibition of NF-κB and AP-1 Activation in LPS-Stimulated RAW264.7 Macrophages. Evid.-Based Complement. Altern. Med. 2018, 2018, 6914514. [Google Scholar] [CrossRef] [PubMed]
- Porta, C.; Riboldi, E.; Ippolito, A.; Sica, A. Molecular and Epigenetic Basis of Macrophage Polarized Activation. Semin. Immunol. 2015, 27, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Liang, H.; Zen, K. Molecular Mechanisms That Influence the Macrophage M1–M2 Polarization Balance. Front. Immunol. 2014, 5, 614. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the Full Spectrum of Macrophage Activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Evers, S.; Roeder, D.; Parlow, A.F.; Risteli, J.; Risteli, L.; Lee, Y.C.; Feizi, T.; Langen, H.; Nussenzweig, M.C. Mannose Receptor-Mediated Regulation of Serum Glycoprotein Homeostasis. Science 2002, 295, 1898–1901. [Google Scholar] [CrossRef]
- Gazi, U.; Martinez-Pomares, L. Influence of the Mannose Receptor in Host Immune Responses. Immunobiology 2009, 214, 554–561. [Google Scholar] [CrossRef]
- Liu, H.-F.; Zhang, H.-J.; Hu, Q.-X.; Liu, X.-Y.; Wang, Z.-Q.; Fan, J.-Y.; Zhan, M.; Chen, F.-L. Altered Polarization, Morphology, and Impaired Innate Immunity Germane to Resident Peritoneal Macrophages in Mice with Long-Term Type 2 Diabetes. J. Biomed. Biotechnol. 2012, 2012, 867023. [Google Scholar] [CrossRef]
- Sica, A.; Mantovani, A. Macrophage Plasticity and Polarization: In Vivo Veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Daniel, B.; Nagy, G.; Czimmerer, Z.; Horvath, A.; Hammers, D.W.; Cuaranta-Monroy, I.; Poliska, S.; Tzerpos, P.; Kolostyak, Z.; Hays, T.T.; et al. The Nuclear Receptor PPARγ Controls Progressive Macrophage Polarization as a Ligand-Insensitive Epigenomic Ratchet of Transcriptional Memory. Immunity 2018, 49, 615–626. [Google Scholar] [CrossRef] [PubMed]
- BD Biosciences. BD FACSCelestaTM Flow Cytometer User’s Guide; BD Biosciences: San Jose, CA, USA, 2020. [Google Scholar]
- Ottoni, M.H.F.; dos Santos, M.G.; de Almeida, V.G.; de Costa, L.A.; Meireles, A.B.; de Avelar-Freitas, B.A.; dos Santos, J.A.T.; de Fátima Pereira, W.; Brito-Melo, G.E.A. Background Autofluorescence Induced by Plant Extracts in Human Lymphocytes: A Flow Cytometric Analysis of a Critical Bias. J. Immunol. Methods 2019, 468, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Romero-Ramírez, L.; Mey, J. Retinoic Acid Increases Phagocytosis of Myelin by Macrophages. J. Cell. Physiol. 2021, 236, 3929–3945. [Google Scholar] [CrossRef] [PubMed]
- Martinet, W.; Schrijvers, D.M.; Timmermans, J.-P.; Herman, A.G.; De Meyer, G.R.Y. Phagocytosis of Bacteria Is Enhanced in Macrophages Undergoing Nutrient Deprivation. FEBS J. 2009, 276, 2227–2240. [Google Scholar] [CrossRef]
- Clemente, L.P.; Rabenau, M.; Tang, S.; Stanka, J.; Cors, E.; Stroh, J.; Culmsee, C.; von Karstedt, S. Dynasore Blocks Ferroptosis through Combined Modulation of Iron Uptake and Inhibition of Mitochondrial Respiration. Cells 2020, 9, 2259. [Google Scholar] [CrossRef]
- Preta, G.; Cronin, J.G.; Sheldon, I.M. Dynasore—Not Just a Dynamin Inhibitor. Cell Commun. Signal. 2015, 13, 24. [Google Scholar] [CrossRef]
- Paul, D.; Achouri, S.; Yoon, Y.-Z.; Herre, J.; Bryant, C.E.; Cicuta, P. Phagocytosis Dynamics Depends on Target Shape. Biophys. J. 2013, 105, 1143–1150. [Google Scholar] [CrossRef]
- Greaves, D.R.; Gordon, S. The Macrophage Scavenger Receptor at 30 Years of Age: Current Knowledge and Future Challenges. J. Lipid Res. 2009, 50, S282–S286. [Google Scholar] [CrossRef]
- McNeil, P.L.; Tanasugarn, L.; Meigs, J.B.; Taylor, D.L. Acidification of Phagosomes Is Initiated before Lysosomal Enzyme Activity Is Detected. J. Cell Biol. 1983, 97, 692–702. [Google Scholar] [CrossRef]
- McCall, J.R.; Sausman, K.T. Systematic Approach in Macrophage Polarization Experiments: Maintaining Integrity and Reproducibility Using Flow Cytometry and Sample Preparation. J. Immunol. Methods 2021, 492, 112969. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minns, L.A.; Sausman, K.T.; Brown, A.P.; York, R.A.; McCall, J.R. Karenia brevis Extract Induces Cellular Entry through Distinct Mechanisms in Phagocytic RAW 264.7 Macrophages versus Non-Phagocytic Vero Cells. Mar. Drugs 2024, 22, 4. https://doi.org/10.3390/md22010004
Minns LA, Sausman KT, Brown AP, York RA, McCall JR. Karenia brevis Extract Induces Cellular Entry through Distinct Mechanisms in Phagocytic RAW 264.7 Macrophages versus Non-Phagocytic Vero Cells. Marine Drugs. 2024; 22(1):4. https://doi.org/10.3390/md22010004
Chicago/Turabian StyleMinns, Laurie A., Kathryn T. Sausman, Ariel P. Brown, Robert A. York, and Jennifer R. McCall. 2024. "Karenia brevis Extract Induces Cellular Entry through Distinct Mechanisms in Phagocytic RAW 264.7 Macrophages versus Non-Phagocytic Vero Cells" Marine Drugs 22, no. 1: 4. https://doi.org/10.3390/md22010004
APA StyleMinns, L. A., Sausman, K. T., Brown, A. P., York, R. A., & McCall, J. R. (2024). Karenia brevis Extract Induces Cellular Entry through Distinct Mechanisms in Phagocytic RAW 264.7 Macrophages versus Non-Phagocytic Vero Cells. Marine Drugs, 22(1), 4. https://doi.org/10.3390/md22010004