
An Eco-Friendly Extraction and Purification Approach for Obtaining Active Ingredients for Cosmetics from Two Marine Brown Seaweeds

 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
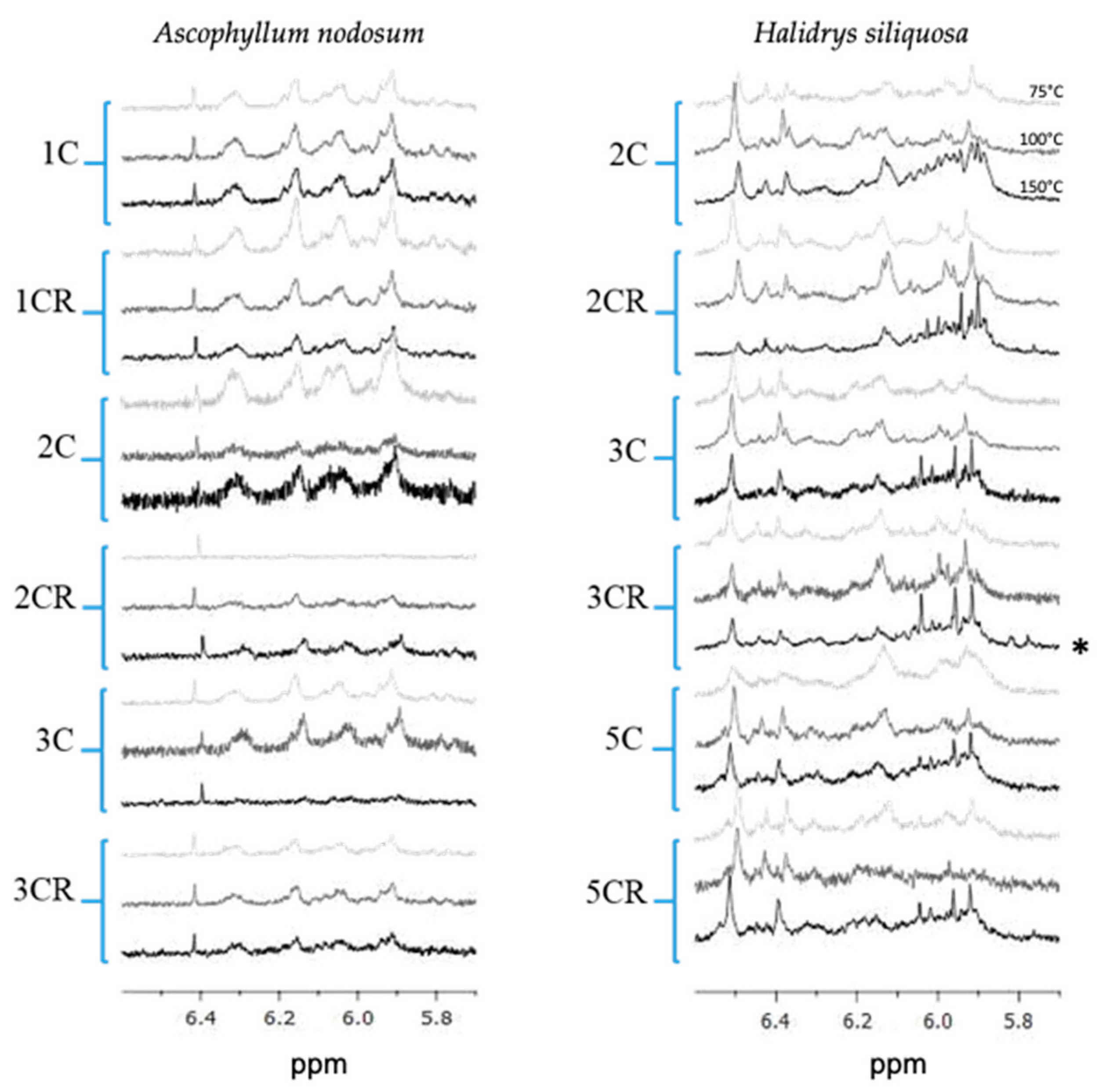
2.1. 1H NMR Spectra of ASE Extracts
2.2. Total Phenolic Content (TPC) and Extraction Yield Using ASE Process
2.3. Antioxidant Activities of ASE Extracts
2.4. Photoprotective Sunscreen Activities of ASE Extracts
2.5. Anti-Aging Activities of ASE Extracts
2.6. Correlation between Phenolic Compounds and Activities for ASE Extracts
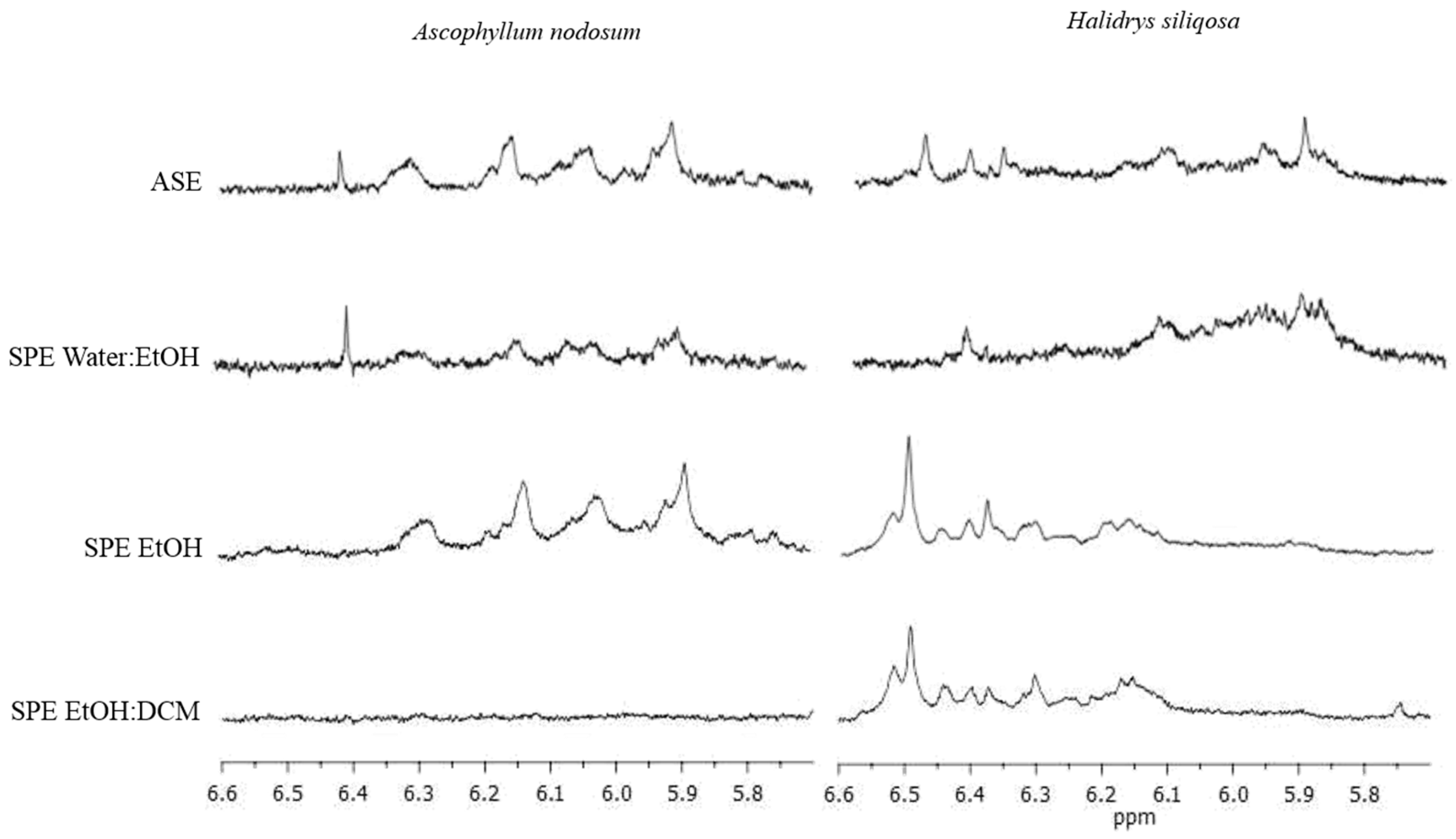
2.7. Semi-Purification of ASE Extracts Using SPE Procedure
2.8. Interest in Using NMR to Check the Presence/Absence of Phlorotannins in Extracts
3. Materials and Methods
3.1. Algal Material
3.2. Extraction and Purification Procedures
3.3. Quantification of Phenolic Compounds
3.4. Activity Tests
3.5. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mesnildrey, L.; Jacob, C.; Frangoudes, K.; Reunavot, M.; Lesueur, M. Seaweed industry in France. In Report Interreg Program NETALGAE; 2012; 42p, Available online: https://institut-agro-rennes-angers.hal.science/hal-00840572 (accessed on 22 February 2024).
- Geiselman, J.A.; McConnell, O.J. Polyphenols in brown algae Fucus vesiculosus and Ascophyllum nodosum: Chemical defenses against the marine herbivorous snail, Littorina littorea. J. Chem. Ecol. 1981, 7, 1115–1133. [Google Scholar] [CrossRef] [PubMed]
- Jennings, J.G.; Steinberg, P.D. Phlorotannins versus other factors affecting epiphyte abundance on the kelp Ecklonia radiata. Oecologia 1997, 109, 461–473. [Google Scholar] [CrossRef]
- Swanson, A.K.; Druehl, L.D. Induction, exudation and the UV protective role of kelp phlorotannins. Aq. Bot. 2002, 73, 241–253. [Google Scholar] [CrossRef]
- Amsler, C.D. Chemical ecology of seaweeds. In Seaweed Biology: Ecological Studies (Analysis and Synthesis); Wiecke, C., Bishof, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 219, pp. 177–188. [Google Scholar]
- Lalegerie, F.; Gager, L.; Stiger-Pouvreau, V.; Connan, S. The stressful life of red and brown seaweeds on the temperate intertidal zone: Effect of abiotic and biotic parameters on the physiology of macroalgae and content variability of particular metabolites. Adv. Bot. Res. 2020, 95, 247–287. [Google Scholar] [CrossRef]
- Stiger-Pouvreau, V.; Guérard, F. Bio-inspired molecules extracted from marine macroalgae: A new generation of active ingredients for cosmetics and human health. In Blue Biotechnology: Production and Use of Marine Molecules; La Barre, S., Bates, S.S., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2018; pp. 709–746. [Google Scholar] [CrossRef]
- Couteau, C.; Coiffard, L. Phycocosmetics and other marine cosmetics, specific cosmetics formulated using marine resources. Mar. Drugs 2020, 18, 322. [Google Scholar] [CrossRef] [PubMed]
- Ragan, M.A.; Glombitza, K.W. Phlorotannins, brown algal polyphenols. Prog. Phycol. Res. 1986, 4, 129–241. [Google Scholar]
- Shalaby, E. Algae as promising organisms for environment and health. Plant Signal. Behav. 2011, 6, 1338–1350. [Google Scholar] [CrossRef]
- Stengel, D.B.; Connan, S.; Popper, Z.A. Algal chemodiversity and bioactivity: Sources of natural variability and implications for commercial application. Biotechnol. Adv. 2011, 29, 483–501. [Google Scholar] [CrossRef]
- Tanniou, A.; Esteban, S.L.; Laurent, V.; Ibáñez, E.; Jose, A.M.; Cérantola, S.; Kervarec, N.; La Barre, S.; Marchal, L.; Stiger-Pouvreau, V. Green improved processes to extract bioactive phenolic compounds from brown macroalgae using Sargassum muticum as model. Talanta 2013, 104, 44–52. [Google Scholar] [CrossRef]
- Stengel, D.B.; Connan, S. Marine algae: A source of biomass for biotechnological applications. In Natural Products From Marine Algae: Methods and Protocols; Stengel, D.B., Connan, S., Eds.; Springer Science+Business Media: New York, NY, USA, 2015; Volume 1308, pp. 103–108. [Google Scholar] [CrossRef]
- Couteau, C.; Coiffard, L. Seaweed application in cosmetics. In Seaweed in Health and Disease Prevention; Elsevier, Inc.: London, UK, 2016; pp. 423–441. [Google Scholar] [CrossRef]
- Le Lann, K.; Surget, G.; Couteau, C.; Coiffard, L.; Cérantola, S.; Gaillard, F.; Larnicol, M.; Zubia, M.; Guérard, F.; Poupart, N.; et al. Sunscreen, antioxidant, and bactericide capacities of phlorotannins from the brown macroalga Halidrys siliquosa. J. Appl. Phycol. 2016, 28, 3547–3559. [Google Scholar] [CrossRef]
- Gager, L.; Lalegerie, F.; Connan, S.; Stiger-Pouvreau, V. Marine Algal Derived Phenolic Compounds and their Biological Activities for Medicinal and Cosmetic Applications. In Recent Advances in Micro and Macroalgal Processing: Food and Health Perspectives; Rajauria, G., Yuan, Y.V., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2021; pp. 278–334. [Google Scholar] [CrossRef]
- Esquivel-Hernández, D.A.; Ibarra-Garza, I.P.; Rodrìguez-Rodrìguez, J.; Cuéllar-Bermùdez, S.P.; Rostro-Alanis, M.d.J.; Alemán-Nava, G.S.; Garcìa- Pérez, J.S.; Parra-Saldìvar, R. Green extraction technologies for high-value metabolites from algae: A review. Biofuels Bioprod. Biorefin. 2017, 11, 215–231. [Google Scholar] [CrossRef]
- Murray, P.M.; Moane, S.; Collins, C.; Beletskaya, T.; Thomas, O.P.; Duarte, A.W.; Nobre, F.S.; Owoyemi, I.O.; Pagnocca, F.C.; Sette, L.; et al. Sustainable production of biologically active molecules of marine based origin. New Biotechnol. 2013, 30, 839–850. [Google Scholar] [CrossRef]
- Dizerbo, A.; Herpe, E. Liste et Répartition des Algues Marines des Côtes Françaises de la Manche et de L’atlantique, Iles Anglo-Normandes Incluses; Éditions Scientifiques Anaximandre: Landerneau, France, 2007. [Google Scholar]
- Bourgougnon, N.; Stiger-Pouvreau, V. Chemodiversity and bioactivity within red and brown macroalgae along the french coasts, metropole and overseas departements and territories. In Handbook of Marine Macroalgae: Biotechnology and Applied Phycology, 1st ed.; Kim, S.-K., Ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2011; pp. 58–105. [Google Scholar] [CrossRef]
- Burel, T.; Schaal, G.; Grall, J.; Le Duff, M.; Chapalain, G.; Schmitt, B.; Gemin, M.; Boucher, O.; Ar Gall, E. Small-scale effects of hydrodynamics on the structure of intertidal macroalgal communities: A novel approach. Estuar. Coast. Shelf Sci. 2019, 226, 106290. [Google Scholar] [CrossRef]
- Cérantola, S.; Breton, F.; Ar Gall, E.; Deslandes, E. Co-occurrence and antioxidant activities of fucol and fucophlorethol classes of polymeric phenols in Fucus spiralis. Bot. Mar. 2006, 49, 347–351. [Google Scholar] [CrossRef]
- Breton, F.; Cérantola, S.; Ar Gall, E. Distribution and radical scavenging activity of phenols in Ascophyllum nodosum (Phaeophyceae). J. Exp. Mar. Biol. Ecol. 2011, 399, 167–172. [Google Scholar] [CrossRef]
- Stiger-Pouvreau, V.; Jegou, C.; Cerantola, S.; Guérard, F.; Le Lann, K. Phlorotannins in Sargassaceae species from Brittany (France): Interesting molecules for ecophysiological and valorisation purposes. Adv. Bot. Res. 2014, 71, 379–411. [Google Scholar] [CrossRef]
- Catarino, M.D.; Pires, S.M.; Silva, S.; Costa, F.; Braga, S.S.; Pinto, D.C.; Silva, A.M.S.; Cardoso, S.M. Overview of phlorotannins’ constituents in Fucales. Mar. Drugs 2022, 20, 754. [Google Scholar] [CrossRef] [PubMed]
- Connan, S.; Goulard, F.; Stiger, V.; Deslandes, E.; Ar Gall, E. Interspecific and temporal variation in phlorotannin levels in an assemblage of brown algae. Bot. Mar. 2004, 47, 410–416. [Google Scholar] [CrossRef]
- Zubia, M.; Fabre, M.S.; Kerjean, V.; Lann, K.L.; Stiger-Pouvreau, V.; Fauchon, M.; Deslandes, E. Antioxidant and antitumoural activities of some Phaeophyta from Brittany coasts. Food Chem. 2009, 116, 693–701. [Google Scholar] [CrossRef]
- Chemat, F.; Vian, M.A.; Cravotto, G. Green extraction of natural products: Concept and principles. Int. J. Mol. Sci. 2012, 13, 8615–8627. [Google Scholar] [CrossRef] [PubMed]
- Meng, W.; Mu, T.; Sun, H.; Garcia-Vaquero, M. Phlorotannins: A review of extraction methods, structural characteristics, bioactivities, bioavailability, and future trends. Algal Res. 2021, 60, 102484. [Google Scholar] [CrossRef]
- Hardouin, K.; Bedoux, G.; Burlot, A.S.; Donnay-Moreno, C.; Bergé, J.P.; Nyvall-Collèn, P.; Bourgougnon, N. Enzyme-assisted extraction (EAE) for the production of antiviral and antioxidant extracts from the green seaweed Ulva armoricana (Ulvales, Ulvophyceae). Algal Res. 2016, 16, 233–239. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K. Algal extracts: Technology and advances. Eng. Life Sci. 2014, 14, 581–591. [Google Scholar] [CrossRef]
- Poojary, M.M.; Barba, F.J.; Aliakbarian, B.; Donsì, F.; Pataro, G.; Dias, D.A.; Juliano, P. Innovative alternative technologies to extract carotenoids from microalgae and seaweeds. Mar. Drugs 2016, 14, 214. [Google Scholar] [CrossRef]
- del Pilar Sánchez-Camargo, A.; Montero, L.; Stiger-Pouvreau, V.; Tanniou, A.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Considerations on the use of enzyme-assisted extraction in combination with pressurized liquids to recover bioactive compounds from algae. Food Chem. 2016, 192, 67–74. [Google Scholar] [CrossRef]
- Montero, L.; Sánchez-Camargo, A.P.; García-Cañas, V.; Tanniou, A.; Stiger-Pouvreau, V.; Russo, M.; Rastrelli, L.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Anti-proliferative activity and chemical characterization by comprehensive two- dimensional liquid chromatography coupled to mass spectrometry of phlorotannins from the brown macroalga Sargassum muticum collected on North-Atlantic coasts. J. Chromatogr. A 2016, 1428, 115–125. [Google Scholar] [CrossRef]
- Turner, C.; Waldebäck, M. Principles of pressurized fluid extraction and environmental, food and agricultural applications. In Separation, Extraction and Concentration in the Food, Beverage and Nutraceutical Industries. Food Science; Rizvi, S.H., Ed.; Technology and Nutrition; WoodHead Publishing: Philadelphia, PA, USA, 2010; pp. 39–70. [Google Scholar] [CrossRef]
- Richter, B.E.; Jones, B.A.; Ezzell, J.L.; Porter, N.L.; Avdalovic, N.; Pohl, C. Accelerated Solvent Extraction: A technique for sample preparation. Anal. Chem. 1996, 68, 1033–1039. [Google Scholar] [CrossRef]
- Sumampouw, G.A.; Jacobsen, C.; Getachew, A.T. Optimization of phenolic antioxidants extraction from Fucus vesiculosus by pressurized liquid extraction. J. Appl. Phycol. 2021, 33, 1195–1207. [Google Scholar] [CrossRef]
- O’Sullivan, A.M.; OíCallaghan, Y.C.; OíGrady, M.N.; Hayes, M.; Kerry, J.P.; OíBrien, N.M. The effect of solvents on the antioxidant activity in Caco-2 cells of Irish brown seaweed extracts prepared using accelerated solvent extraction (ASE®). J. Funct. Foods 2013, 5, 940–948. [Google Scholar] [CrossRef]
- Tierney, M.S.; Smyth, T.J.; Hayes, M.; Soler-Vila, A.; Croft, A.K.; Brunton, N. Influence of pressurised liquid extraction and solid-liquid extraction methods on the phenolic content and antioxidant activities of Irish macroalgae. Int. J. Food Sci. Technol. 2013, 48, 860–869. [Google Scholar] [CrossRef]
- Santana, C.M.; Ferrera, Z.S.; Padrón, M.E.T.; Rodrìguez, J.J.S. Methodologies for the extraction of phenolic compounds from environmental samples: New approaches. Molecules 2009, 14, 298–320. [Google Scholar] [CrossRef] [PubMed]
- Tanniou, A.; Vandanjon, L.; Incera, M.; Serrano Leon, E.; Husa, V.; Le Grand, J.; Nicolas, J.-L.; Poupart, N.; Kervarec, N.; Engelen, A.; et al. Assessment of the spatial variability of phenolic contents and associated bioactivities in the invasive alga Sargassum muticum sampled along its European range from Norway to Portugal. J. Appl. Phycol. 2014, 26, 1215–1230. [Google Scholar] [CrossRef]
- Jégou, C.; Kervarec, N.; Cérantola, S.; Bihannic, I.; Stiger-Pouvreau, V. NMR use to quantify phlorotannins: The case of Cystoseira tamariscifolia, a phloroglucinol-producing brown macroalga in Brittany (France). Talanta 2015, 135, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Le Lann, K.; Kervarec, N.; Payri, C.E.; Deslandes, E.; Stiger-Pouvreau, V. Discrimination of allied species within the genus Turbinaria (Fucales, Phaeophyceae) using HRMAS NMR spectroscopy. Talanta 2008, 74, 1079–1083. [Google Scholar] [CrossRef]
- Le Lann, K.; Kraffe, E.; Kervarec, N.; Cérantola, S.; Payri, C.E.; Stiger-Pouvreau, V. Isolation of turbinaric acid as a chemomarker of Turbinaria conoides (J. Agardh) Kützing from South Pacific Islands. J. Phycol. 2014, 50, 1048–1057. [Google Scholar] [CrossRef]
- Ar Gall, E.; Lelchat, F.; Hupel, M.; Jégou, C.; Stiger-Pouvreau, V. Extraction and purification of phlorotannins from brown algae. In Natural Products from Marine Algae; Stengel, D.B., Connan, S., Eds.; Humana Press: New York, NY, USA, 2015; pp. 131–143. [Google Scholar] [CrossRef]
- Gager, L.; Connan, S.; Molla, M.; Couteau, C.; Arbona, J.-F.; Coiffard, L.; Cérantola, S.; Stiger-Pouvreau, V. Temporal variation of active phlorotannins determined by 1H NMR and in vitro assays. J. Appl. Phycol. 2020, 32, 2375–2386. [Google Scholar] [CrossRef]
- Plaza, M.; Santoyo, S.; Jaime, L.; García-Blairsy Reina, G.; Herrero, M.; Señorans, F.J.; Ibáñez, E. Screening for bioactive compounds from algae. Adv. Biofuels Bioprod. 2010, 9781461433, 833–872. [Google Scholar] [CrossRef]
- Herrero, M.; Plaza, M.; Cifuentes, A.; Ibáñez, E. Green processes for extraction of bioactives from Rosemary: Chemical and functional characterization via UPLC-MS/MS and In-Vitro assays. J. Chromatogr. A. 2010, 1217, 2512–2520. [Google Scholar] [CrossRef] [PubMed]
- Onofrejová, L.; Vašíčková, J.; Klejdus, B.; Stratil, P.; Mišurcová, L.; Kráčmar, S.; Kopecký, J.; Vacek, J. Bioactive phenols in algae: The application of pressurized-liquid and solid-phase extraction techniques. J. Pharm. Biomed. Anal. 2010, 51, 464–470. [Google Scholar] [CrossRef]
- Kronholm, J.; Hartonen, K.; Riekkola, M.L. Analytical extractions with water at elevated temperatures and pressures. TrAC 2007, 26, 396–412. [Google Scholar] [CrossRef]
- Santoyo, S.; Rodríguez-Meizoso, I.; Cifuentes, A.; Jaime, L.; García-Blairsy Reina, G.; Señorans, F.J.; Ibáñez, E. Green processes based on the extraction with pressurized fluids to obtain potent antimicrobials from Haematococcus pluvialis microalgae. LWT Food Sci. Technol. 2009, 42, 1213–1218. [Google Scholar] [CrossRef]
- Rodríguez-Meizoso, I.; Jaime, L.; Santoyo, S.; Señorans, F.J.; Cifuentes, A.; Ibáñez, E. Subcritical water extraction and characterization of bioactive compounds from Haematococcus pluvialis microalga. J. Pharm. Biomed. Anal. 2010, 51, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Getachew, A.T.; Jacobsen, C.; Holdt, S.L. Emerging technologies for the extraction of marine phenolics: Opportunities and challenges. Mar. Drugs 2020, 18, 389. [Google Scholar] [CrossRef] [PubMed]
- Le Lann, K.; Connan, S.; Stiger-Pouvreau, V. Phenology, TPC and size-fractioning phenolics variability in temperate Sargassaceae (Phaeophyceae, Fucales) from Western Brittany: Native versus introduced species. Mar. Env. Res. 2012, 80, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Couteau, C.; Pommier, M.; Paparis, E.; Coiffard, L.J.M. Study of the efficacy of 18 sun filters authorized in European Union tested in vitro. Pharmazie 2007, 62, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Puspita, M.; Deniel, M.; Widowati, I.; Radjasa, O.; Douzenel, P.; Bedoux, G.; Bourgougnon, N. Antioxidant and antibacterial activity of solid-liquid and enzyme-assisted extraction of phenolic compound from three species of tropical Sargassum. IOP Conf. Ser. Earth Environ. Sci. 2017, 55, 533–539. [Google Scholar] [CrossRef]
- Balboa, E.M.; Conde, E.; Moure, A.; Falqué, E.; Domínguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef] [PubMed]
- Audibert, L.; Fauchon, M.; Blanc, N.; Hauchard, D.; Ar Gall, E. Phenolic compounds in the brown seaweed Ascophyllum nodosum: Distribution and radical-scavenging activities. Phytochem. Anal. 2010, 21, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Kuda, T.; Ikemori, T. Minerals, polysaccharides and antioxidant properties of aqueous solutions obtained from macroalgal beach-casts in the Noto Peninsula, Ishikawa, Japan. Food Chem. 2009, 112, 575–581. [Google Scholar] [CrossRef]
- Rajauria, G.; Jaiswal, A.K.; Abu-Ghannam, N.; Gupta, S. Effect of hydrothermal processing on colour, antioxidant and free radical scavenging capacities of edible Irish brown seaweeds. Int. J. Food Sci. Technol. 2010, 45, 2485–2493. [Google Scholar] [CrossRef]
- Yangthong, M.; Hutadilok-Towatana, N.; Phromkunthong, W. Antioxidant activities of four edible seaweeds from the Southern coast of Thailand. Plant Foods Hum. Nutr. 2009, 64, 218–223. [Google Scholar] [CrossRef]
- Mhadhebi, L.; Laroche-Clary, A.; Robert, J.; Bouraoui, A. Antioxidant, anti- inflammatory, and antiproliferative activities of organic fractions from the Mediterranean brown seaweed Cystoseira sedoides. Can. J. Physiol. Pharmacol. 2011, 89, 911–921. [Google Scholar] [CrossRef]
- Wang, T.; Jonsdottir, R.; Ólafsdóttir, G. Total phenolic compounds, radical scavenging and metal chelation of extracts from Icelandic seaweeds. Food Chem. 2009, 116, 240–248. [Google Scholar] [CrossRef]
- Wang, T.; Jónsdóttir, R.; Liu, H.; Gu, L.; Kristinsson, H.G.; Raghavan, S.; Ólafsdóttir, G. Antioxidant capacities of phlorotannins extracted from the brown algae Fucus vesiculosus. J. Agri. Food Chem. 2012, 60, 5874–5883. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Rajauria, G. Optimization and validation of reverse phase HPLC method for qualitative and quantitative assessment of polyphenols in seaweed. J. Pharmaceut. Biomed. Anal. 2018, 148, 230–237. [Google Scholar] [CrossRef]
- Allwood, J.W.; Evans, H.; Austin, C.; McDougall, G.J. Extraction, enrichment, and LC-MSn-based characterization of phlorotannins and related phenolics from the brown seaweed, Ascophyllum nodosum. Mar. Drugs 2020, 18, 448. [Google Scholar] [CrossRef]
- Wekre, M.E.; Hellesen Brunvoll, S.; Jordheim, M. Advancing quantification methods for polyphenols in brown seaweeds—Applying a selective qNMR method compared with the TPC assay. Phytochem. Anal. 2022, 33, 1099–1110. [Google Scholar] [CrossRef]
- Linares, I.B.; Arráez-Román, D.; Herrero, M.; Ibáñez, E.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Comparison of different extraction procedures for the comprehensive characterization of bioactive phenolic compounds in Rosmarinus officinalis by reversed-phase high-performance liquid chromatography with diode array detection coupled to electrospray time-of-flight mass spectrometry. J. Chromat. A 2011, 1218, 7682–7690. [Google Scholar] [CrossRef]
- Ford, L.; Theodoridou, K.; Sheldrake, G.N.; Walsh, P.J. A critical review of analytical methods used for the chemical characterisation and quantification of phlorotannin compounds in brown seaweeds. Phytochem. Anal. 2019, 30, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Sallenave, J.M.; Xing, Z.; Simpson, A.J.; Graham, F.L.; Gauldie, J. Adenovirus-mediated expression of an elastase-specific inhibitor (elafin): A comparison of different promoters. Gene Ther. 1998, 5, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.W.C.; Lim, Y.Y.; Wong, L.F.; Lianto, F.S.; Wong, S.K.; Lim, K.K.; Joe, C.E.; Lim, T.Y. Antioxidant and tyrosinase inhibition properties of leaves and rhizomes of ginger species. Food Chem. 2008, 109, 477–483. [Google Scholar] [CrossRef]
- Lim, T.Y.; Lim, Y.Y.; Yule, C.M. Evaluation of antioxidant, antibacterial and anti-tyrosinase activities of four Macaranga species. Food Chem. 2009, 114, 594–599. [Google Scholar] [CrossRef]
- Couteau, C.; Philippe, A.; Vibet, M.A.; Paparis, E.; Coiffard, L. Study of the influence of substrate and spectrophotometer characteristics on the In Vitro measurement of sunscreens efficiency. Eur. J. Pharmaceut. Sci. 2018, 12, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Domínguez, M.C.; Mendiola, J.A.; Sánchez-Martínez, J.D.; Bueno, M.; Cerezal-Mezquita, P.; Ibáñez, E. Evaluation of the antioxidant and neuroprotective activity of the seaweed Durvillaea antarctica (cochayuyo) extracts using pressurized liquids. J. Appl. Phycol. 2023, 35, 835–847. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Cycle | Temperature (°C) | Yield (%) |
|---|---|---|---|
| Ascophyllum nodosum | 1C | 75 | 10.9 ± 0.2 c |
| 100 | 13.2 ± 0.4 b | ||
| 150 | 16.5 ± 1.3 a | ||
| 1CR | 75 | 13.8 ± 0.4 a | |
| 100 | 13.1 ± 4.1 a | ||
| 150 | 22.5 ± 1.1 a | ||
| 2C | 75 | 8.4 ± 3.9 b | |
| 100 | 13.0 ± 1.3 b | ||
| 150 | 19.2 ± 0.7 a | ||
| 2CR | 75 | 20.9 ± 0.6 b | |
| 100 | 22.2 ± 1.1 b | ||
| 150 | 26.4 ± 0.3 a | ||
| 3C | 75 | 15.8 ± 0.7 c | |
| 100 | 19.0 ± 1.1 b | ||
| 150 | 24.7 ± 0.3 a | ||
| 3CR | 75 | 22.0 ± 1.2 b | |
| 100 | 20.6 ± 0.7 b | ||
| 150 | 27.6 ± 0.6 a | ||
| Halidrys siliquosa | 2C | 75 | 28.4 ± 4.1 a |
| 100 | 35.6 ± 0.9 a | ||
| 150 | 34.0 ± 8.9 a | ||
| 2CR | 75 | 38.3 ± 0.6 b | |
| 100 | 40.5 ± 0.4 b | ||
| 150 | 49.5 ± 2.5 a | ||
| 3C | 75 | 33.2 ± 0.8 b | |
| 100 | 37.1 ± 1.0 ab | ||
| 150 | 38.9 ± 3.2 a | ||
| 3CR | 75 | 36.5 ± 3.4 b | |
| 100 | 40.3 ± 0.8 ab | ||
| 150 | 53.9 ± 9.6 a | ||
| 5C | 75 | 35.2 ± 1.7 b | |
| 100 | 32.6 ± 0.6 b | ||
| 150 | 48.0 ± 0.4 a | ||
| 5CR | 75 | 38.7 | |
| 100 | 42.5 | ||
| 150 | 30.9 |
| Cycle | Temperature (°C) | SPF | PF-UVA |
|---|---|---|---|
| 2C | 75 | 1.32 ± 0.04 b | 1.12 ± 0.04 b |
| 100 | 1.29 ± 0.05 b | 1.17 ± 0.03 b | |
| 150 | 2.33 ± 0.46 a | 1.85 ± 0.20 a | |
| 2CR | 75 | 1.30 ± 0.06 a | 1.19 ± 0.04 a |
| 100 | 1.22 ± 0.02 a | 1.12 ± 0.01 a | |
| 150 | 1.57 ± 0.40 a | 1.27 ± 0.21 a | |
| 3C | 75 | 1.29 ± 0.04 b | 1.18 ± 0.03 b |
| 100 | 1.41 ± 0.06 b | 1.24 ± 0.04 b | |
| 150 | 1.93 ± 0.49 a | 1.57 ± 0.40 a | |
| 3CR | 75 | 1.24 ± 0.03 b | 1.13 ± 0.02 b |
| 100 | 1.31 ± 0.03 b | 1.17 ± 0.02 b | |
| 150 | 2.17 ± 0.15 a | 1.65 ± 0.13 a | |
| 5C | 75 | 1.26 ± 0.03 b | 1.14 ± 0.02 b |
| 100 | 1.36 ± 0.07 b | 1.22 ± 0.04 b | |
| 150 | 2.23 ± 0.28 a | 1.67 ± 0.21 a | |
| 5CR | 75 | 1.53 ± 0.07 b | 1.31 ± 0.05 b |
| 100 | 1.21 ± 0.05 c | 1.10 ± 0.04 c | |
| 150 | 2.30 ± 0.12 a | 1.72 ± 0.07 a |
| Anti-Aging Activities | Extract or Control | Concentrations (mg.mL−1) | % of Inhibition | |
|---|---|---|---|---|
| Elastase inhibition | EGCG | 1 | 19.44 | |
| An | 1C75 | 1 | 57.89 | |
| 1C100 | 1 | 80.46 | ||
| 1C150 | 1 | 47.39 | ||
| Hs | 2C75 | 1 | 20.60 | |
| 2C100 | 1 | 14.54 | ||
| 2C150 | 1 | 29.95 | ||
| Tyrosinase inhibition | Kojic acid | 0.1 | 93.46 | |
| An | 1C75 | 0.1 | 33.33 | |
| 1C100 | 0.1 | 12.43 | ||
| 1C150 | 0.1 | 2.48 | ||
| Hs | 2C75 | 0.1 | 15.94 | |
| 2C100 | 0.1 | 0 | ||
| 2C150 | 0.1 | 42.51 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gager, L.; Connan, S.; Cérantola, S.; Petek, S.; Couteau, C.; Coiffard, L.; Stiger-Pouvreau, V. An Eco-Friendly Extraction and Purification Approach for Obtaining Active Ingredients for Cosmetics from Two Marine Brown Seaweeds. Mar. Drugs 2024, 22, 112. https://doi.org/10.3390/md22030112
Gager L, Connan S, Cérantola S, Petek S, Couteau C, Coiffard L, Stiger-Pouvreau V. An Eco-Friendly Extraction and Purification Approach for Obtaining Active Ingredients for Cosmetics from Two Marine Brown Seaweeds. Marine Drugs. 2024; 22(3):112. https://doi.org/10.3390/md22030112
Chicago/Turabian StyleGager, Leslie, Solène Connan, Stéphane Cérantola, Sylvain Petek, Céline Couteau, Laurence Coiffard, and Valérie Stiger-Pouvreau. 2024. "An Eco-Friendly Extraction and Purification Approach for Obtaining Active Ingredients for Cosmetics from Two Marine Brown Seaweeds" Marine Drugs 22, no. 3: 112. https://doi.org/10.3390/md22030112
APA StyleGager, L., Connan, S., Cérantola, S., Petek, S., Couteau, C., Coiffard, L., & Stiger-Pouvreau, V. (2024). An Eco-Friendly Extraction and Purification Approach for Obtaining Active Ingredients for Cosmetics from Two Marine Brown Seaweeds. Marine Drugs, 22(3), 112. https://doi.org/10.3390/md22030112