

Jasmonates and Ethylene Shape Floridoside Synthesis during Carposporogenesis in the Red Seaweed Grateloupia imbricata



Abstract
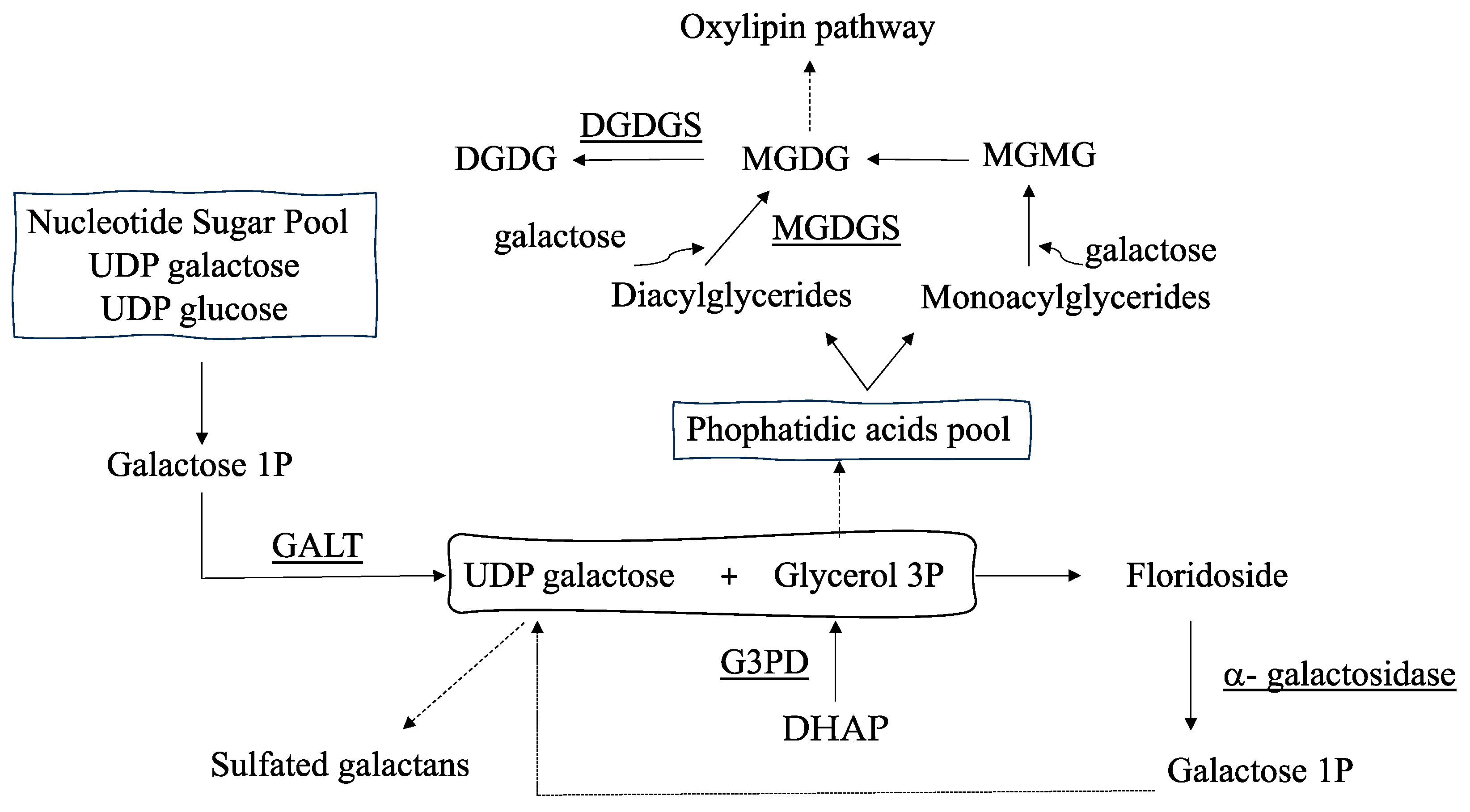
:1. Introduction
2. Results
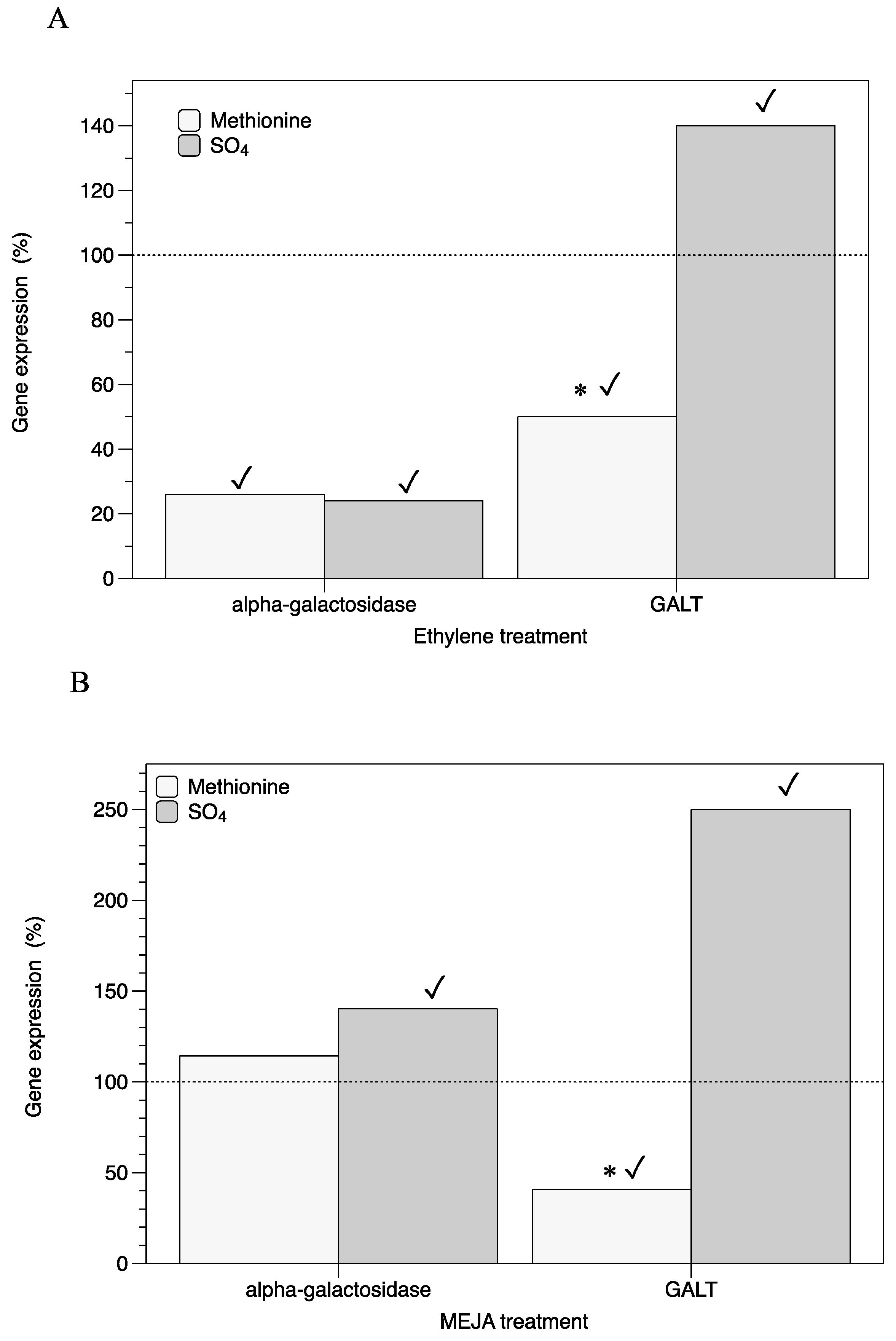
2.1. Influence of Ethylene and Meja on Expression of Genes Involved in Floridoside Synthesis alongside Cystocarp Development
2.2. Influence of Salinity on Gene Expression Involved in Floridoside Synthesis
3. Discussion
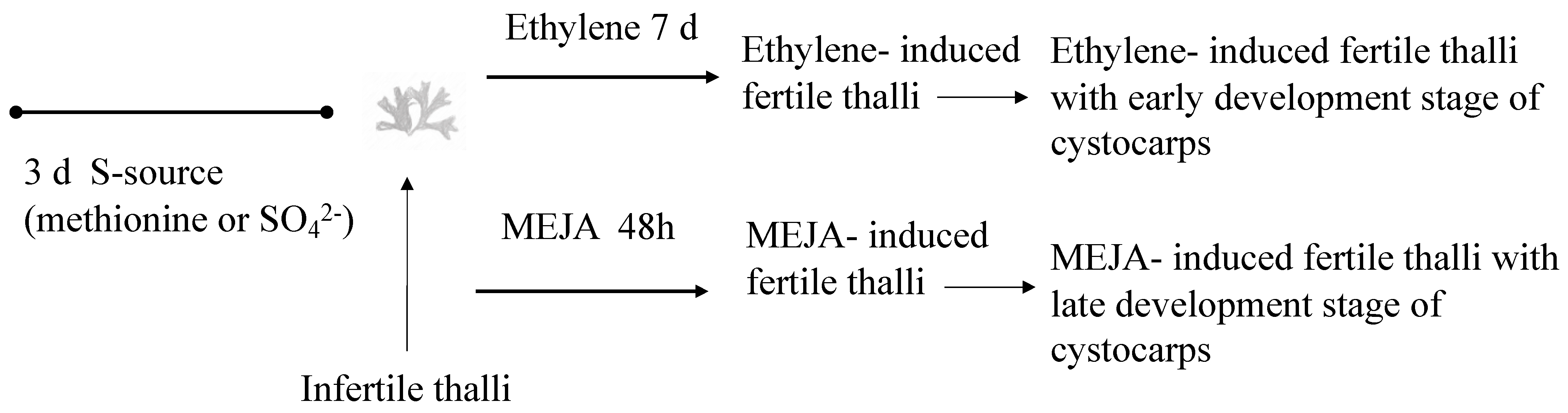
4. Materials and Methods
4.1. Plant Material and Culture Conditions
4.2. Influence of Ethylene and Methyl Jasmonate in Floridoside Synthesis
4.3. Influence of Salinity Changes in Floridoside Synthesis
4.4. RNA Extraction
4.5. Droplet Digital PCR (ddPCR) Primers and Protocol Implementation
4.6. Data Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kremer, B.P.; Kirst, G.O. Biosynthesis of 2-OD-glycerol-α-D-galactopyranoside (Floridoside) in marine Rhodophyceae. Plant Sci. Lett. 1981, 23, 349–357. [Google Scholar] [CrossRef]
- Garcia-Jimenez, P.; Mantesa, S.R.; Robaina, R.R. Expression ofgenes related to carrageenan synthesis during carposporogenesis of the red seaweed Grateloupia imbricata. Mar. Drugs 2020, 18, 432. [Google Scholar] [CrossRef]
- del Rosario-Santana, D.; Robaina, R.R.; Garcia-Jimenez, P. Jasmonates disrupt carrageenan synthesis during carposporogenesis in the red seaweed Grateloupia imbricata. Front. Mar. Sci. 2023, 10, 1188493. [Google Scholar] [CrossRef]
- Del Rosario-Santana, D.; Robaina, R.R.; Garcia-Jimenez, P. S-Assimilation influences in Carrageenan Biosynthesis Genes during Ethylene-Induced Carposporogenesis in Red Seaweed Grateloupia imbricata. Mar. Drugs 2022, 20, 436. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Jimenez, P.; Robaina, R.R. Systems Biology of Marine Ecosystems; Kumar, M., Ralph, P., Eds.; Springer: Gewerbestrasse, Switzerland, 2017; Chapter 5; p. 99. [Google Scholar] [CrossRef]
- Garcia-Jimenez, P.; Robaina, R.R. Effects of ethylene on tetrasporogenesis in Pterocladiella capillacea (Rhodophyta). J. Phycol. 2012, 48, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Jimenez, P.; Brito Romano, O.; Robaina, R.R. Occurrence of jasmonates during cystocarp development in the red alga Grateloupia imbricata. J. Phycol. 2016, 52, 1085–1093. [Google Scholar]
- Yu, Y.; Jia, X.; Wang, W.; Jin, Y.; Liu, W.; Wang, D.; Mao, Y.; Xie, C.; Liu, T. Floridean starch and floridoside metabolic pathways of Neoporphyra haitanensis and their regulatory mechanism under continuous darkness. Mar. Drugs 2021, 19, 664. [Google Scholar] [CrossRef] [PubMed]
- Barbier, G.; Oesterhelt, C.; Larson, M.D.; Halgren, R.G.; Wilkerson, C.; Garavito, R.M.; Benning, C.; Weber, A.P. Comparative genomics of two closely related unicellular thermo-acidophilic red algae, Galdieria sulphuraria and Cyanidioschyzon merolae, reveals the molecular basis of the metabolic flexibility of Galdieria sulphuraria and significant differences in carbohydrate metabolism of both algae. Plant Physiol. 2005, 137, 460–474. [Google Scholar] [PubMed]
- Yu, S. Enzymes of floridean starch and floridoside degradation in red algae: Purification, characterization and physiological studies. Acta Univ. Ups. 1992, 372, 1–47. [Google Scholar]
- Yu, S.; Pedersén, M. The α-galactosidase of Gracilaria tenuistipitata and G. sordida (Gracilariales, Rhodophyta). Phycologia 1990, 29, 454–460. [Google Scholar] [CrossRef]
- Lee, W.K.; Lim, Y.Y.; Leow, A.T.C.; Namasivayam, P.; Abdullah, J.O.; Ho, C.L. Biosynthesis of agar in red seaweeds: A review. Carbohydr. Polym. 2017, 164, 23–30. [Google Scholar] [CrossRef]
- Raymond, J.A.; Morgan-Kiss, R.; Stahl-Rommel, S. Glycerol is an osmoprotectant in two antarctic Chlamydomonas species from an ice-covered saline lake and is synthesized by an unusual bidomain enzyme. Front. Plant Sci. 2020, 11, 1259. [Google Scholar] [CrossRef]
- Robaina, R.R.; Garcia-Jimenez, P.; Garcia-Reina, G.; Luque, A. Morphogenetic effect of glycerol on tissue cultures of the red seaweed Grateloupia doryphora. J. Appl. Phycol. 1990, 2, 137–143. [Google Scholar] [CrossRef]
- Lin, H.; Fang, L.; Low, C.S.; Chow, Y.; Lee, Y.K. Occurrence of glycerol uptake in Dunaliella tertiolecta under hyperosmotic stress. FEBS J. 2013, 280, 1064–1072. [Google Scholar] [CrossRef]
- Yuzawa, Y.; Nishihara, H.; Haraguchi, T.; Masuda, S.; Shimojima, M.; Shimoyama, A.; Yuasa, H.; Okada, N.; Ohta, H. Phylogeny of galactolipid synthase homologs together with their enzymatic analyses revealed a possible origin and divergence time for photosynthetic membrane biogenesis. DNA Res. 2012, 19, 91–102. [Google Scholar] [CrossRef]
- Yu, C.W.; Lin, Y.T.; Li, H.M. Increased ratio of galactolipid MGDG: DGDG induces jasmonic acid overproduction and changes chloroplast shape. New Phytol. 2020, 228, 1327–1335. [Google Scholar] [CrossRef]
- Yu, L.; Zhou, C.; Fan, J.; Shanklin, J.; Xu, C. Mechanisms and functions of membrane lipid remodeling in plants. Plant J. 2021, 107, 37–53. [Google Scholar] [CrossRef]
- Goulard, F.; Diouris, M.; Quere, G.; Deslandes, E.; Flocapos, J.Y. Salinity effects on NDP-sugars, floridoside, starch, and carrageenan yield, and UDP-glucose-pyrophosphorylase and-epimerase activities of cultivated Solieria chordalis. J. Plant Physiol. 2001, 158, 1387–1394. [Google Scholar] [CrossRef]
- Siow, R.-S.; Teo, S.-S.; Ho, W.-Y.; Shukor, M.Y.A.; Phang, S.-M.; Ho, C.-L. Molecular cloning and biochemical characterization of galactose-1-phosphate uridylyltransferase from Gracilaria changii (Rhodophyta). J. Phycol. 2021, 48, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Jimenez, P.; Robaina, R.R.; Montero-Fernández, M. Molecular mechanisms underlying Grateloupia imbricata (Rhodophyta) carposporogenesis induced by methyl jasmonate. J. Phycol. 2017, 53, 1340–1344. [Google Scholar] [CrossRef] [PubMed]
- McQueney, M.S.; Anderson, K.S.; Markham, G.D. Energetics of S-adenosylmethionine synthetase catalysis. Biochemistry 2000, 39, 4443–4454. [Google Scholar] [CrossRef] [PubMed]
- Norbeck, J.; Påhlman, A.K.; Akhtar, N.; Blomberg, A.; Adler, L. Purification and characterization of two isoenzymes of DL-glycerol-3-phosphatase from Saccharomyces cerevisiae: Identification of the corresponding GPP1 and GPP2 genes and evidence for osmotic regulation of Gpp2p expression by the osmosensing mitogen-activated protein kinase signal transduction pathway. J. Biol. Chem. 1996, 271, 13875–13881. [Google Scholar]
- Koornneef, A.; Leon-Reyes, A.; Ritsema, T.; Verhage, A.; Den Otter, F.C.; Van Loon, L.C.; Pieterse, C.M.J. Kinetics of salicylate-mediated suppression of jasmonate signaling reveal a role for redox modulation. Plant Physiol. 2008, 147, 1358–1368. [Google Scholar] [CrossRef] [PubMed]
- Driver, T.; Trivedi, D.K.; McIntosh, O.A.; Dean, A.P.; Goodacre, R.; Pittman, J.K. Two glycerol-3-phosphate dehydrogenases from Chlamydomonas have distinct roles in lipid metabolism. Plant Physiol. 2017, 174, 2083–2097. [Google Scholar] [CrossRef]
- Ansell, R.; Granath, K.; Hohmann, S.; Thevelein, J.M.; Adler, L. The two isoenzymes for yeast NAD+-dependent glycerol 3-phosphate dehydrogenase encoded by GPD1 and GPD2 have distinct roles in osmoadaptation and redox regulation. EMBO J. 1997, 16, 2179–2187. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.J.; Yang, R.; Luo, Q.J.; Chen, J.J.; Chen, H.M.; Yan, X.J. Glycerol-3-phosphate metabolism plays a role in stress response in the red alga Pyropia haitanensis. J. Phycol. 2015, 51, 321–331. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signaling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Weinberger, F.; Lion, U.; Delage, L.; Kloareg, B.; Potin, P.; Beltran, J.; Flores, V.; Faugeron, S.; Correa, J.; Pohnert, G. Up-regulation of lipoxygenase, phospholipase, and oxylipin-production in the induced chemical defense of the red alga Gracilaria chilensis against epiphytes. J. Chem. Ecol. 2011, 33, 677–686. [Google Scholar] [CrossRef]
- Higashi, Y.; Okazaki, Y.; Myouga, F.; Shinozaki, K.; Saito, K. Landscape of the lipidome and transcriptome under heat stress in Arabidopsis thaliana. Sci. Rep. 2015, 5, 10533. [Google Scholar] [CrossRef]
- Yuan, L.; Mao, X.; Zhao, K.; Ji, X.; Ji, C.; Xue, J.; Li, R. Characterisation of phospholipid: Diacylglycerol acyltransferases (PDATs) from Camelina sativa and their roles in stress responses. Biol. Open 2017, 6, 1024–1034. [Google Scholar]
- Luo, S.; Levine, R.L. Methionine in proteins defends against oxidative stress. FASEB J. 2009, 23, 464–472. [Google Scholar] [CrossRef]
- Garcia-Jimenez, P.; Brito-Romano, O.; Robaina, R.R. Production of volatiles by the red seaweed Gelidium arbuscula (Rhodophyta): Emission of ethylene and dimethyl sulfide. J. Phycol. 2013, 49, 661–669. [Google Scholar] [CrossRef] [PubMed]
- André, L.; Hemming, A.; Adler, L. Osmoregulation in Saccharomyces cerevisiae studies on the osmotic induction of glycerol production and glycerol 3-phosphate dehydrogenase (NAD+). FEBS Lett. 1991, 286, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Ericksson, P.; Andre, L.; Ansell, R.; Blomberg, A.; Adler, L. Molecular cloning of GPD2, a second gene encoding sn-glycerol-3-phosphate dehydrogenase (NAD+) in Saccharomyces cerevisiae. Mol. Microbiol. 1995, 17, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Liu, L.; Barkla, B.J. Membrane lipid remodeling in response to salinity. Int. J. Mol. Sci. 2019, 20, 4264. [Google Scholar] [CrossRef] [PubMed]
- Reed, R.H.; Collins, J.C.; Russell, G. The effects of salinity upon galactosyl-glycerol content and concentration of the marine red alga Porphyra purpurea (Roth) C. Ag. J. Exp. Bot. 1980, 31, 1539–1554. [Google Scholar] [CrossRef]
- Kirst, G.O. Low molecular weight carbohydrates and ions in Rhodophyceae: Quantitative measurement of floridoside and digeneaside. Phytochemistry 1980, 19, 1107–1110. [Google Scholar] [CrossRef]
- Ekman, P.; Yu, S.; Pedersen, M. Effects of altered salinity, darkness and algal nutrient status on floridoside and starch content, α-galactosidase activity and agar yield of cultivated Gracilaria sordida. Brit. Phycol. J. 1991, 26, 123–131. [Google Scholar] [CrossRef]
- Arunraj, R.; Skori, L.; Kumar, A.; Hickerson, N.M.; Shoma, N.; Samuel, M.A. Spatial regulation of alpha-galactosidase activity and its influence on raffinose family oligosaccharides during seed maturation and germination in Cicer arietinum. Plant Signal. Behav. 2020, 15, 1709707. [Google Scholar] [CrossRef]
- Giordano, M.; Raven, J.A. Nitrogen and sulfur assimilation in plants and algae. Aquat. Bot. 2014, 118, 45–61. [Google Scholar] [CrossRef]
- Garcia-Jimenez, P.; Montero-Fernández, M.; Robaina, R.R. Analysis of ethylene-induced gene regulation during carposporogenesis in the red seaweed Grateloupia imbricata (Rhodophyta). J. Phycol. 2018, 54, 681–689. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Name | Sequence (5′-3′) |
|---|---|---|
| Synthesis of hexose pool | ||
| alpha-galactosidase | AG-2468F AG-2468R | CTGTCAAGTTCCCGGATTCTC TTCCTGCTGAAAGTCCCATTAG |
| galactose-1-phosphate uridylyl transferase (GALT) | G1PU-1681F G1PU-1681R | GTAGTAGATGCCTGGTGTGATG CATATCTGGCCATGAGGATGAG |
| Synthesis of galactosyl–glycerol pool | ||
| glycerol 3-phosphate dehydrogenase (G3PD) | G3PD-7275F G3PD-7275R | ACCTATCGGGTCCTTCATTTG GGATGAGAACATGTCACCTAGAG |
| monogalactosyl diacylglyceride synthase (MGDGS) | MGS-2948F MGS-2948R | TCCCGTTTAATCACTTCCCTTC ACTAAACGCGGTCTCAGTAATC |
| digalactosyl diacylglyceride synthase (DGDGS) | DGS-3637F DGS-3637R | GTCCAATCCCAATCGAAGAGAG CTCAGCCAGACAATTCCGATAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Jimenez, P.; del Rosario-Santana, D.; Robaina, R.R. Jasmonates and Ethylene Shape Floridoside Synthesis during Carposporogenesis in the Red Seaweed Grateloupia imbricata. Mar. Drugs 2024, 22, 115. https://doi.org/10.3390/md22030115
Garcia-Jimenez P, del Rosario-Santana D, Robaina RR. Jasmonates and Ethylene Shape Floridoside Synthesis during Carposporogenesis in the Red Seaweed Grateloupia imbricata. Marine Drugs. 2024; 22(3):115. https://doi.org/10.3390/md22030115
Chicago/Turabian StyleGarcia-Jimenez, Pilar, Diana del Rosario-Santana, and Rafael R. Robaina. 2024. "Jasmonates and Ethylene Shape Floridoside Synthesis during Carposporogenesis in the Red Seaweed Grateloupia imbricata" Marine Drugs 22, no. 3: 115. https://doi.org/10.3390/md22030115
APA StyleGarcia-Jimenez, P., del Rosario-Santana, D., & Robaina, R. R. (2024). Jasmonates and Ethylene Shape Floridoside Synthesis during Carposporogenesis in the Red Seaweed Grateloupia imbricata. Marine Drugs, 22(3), 115. https://doi.org/10.3390/md22030115