
Induction of Autophagy by Extract from Corydalis heterocarpa for Skin Anti-Aging

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
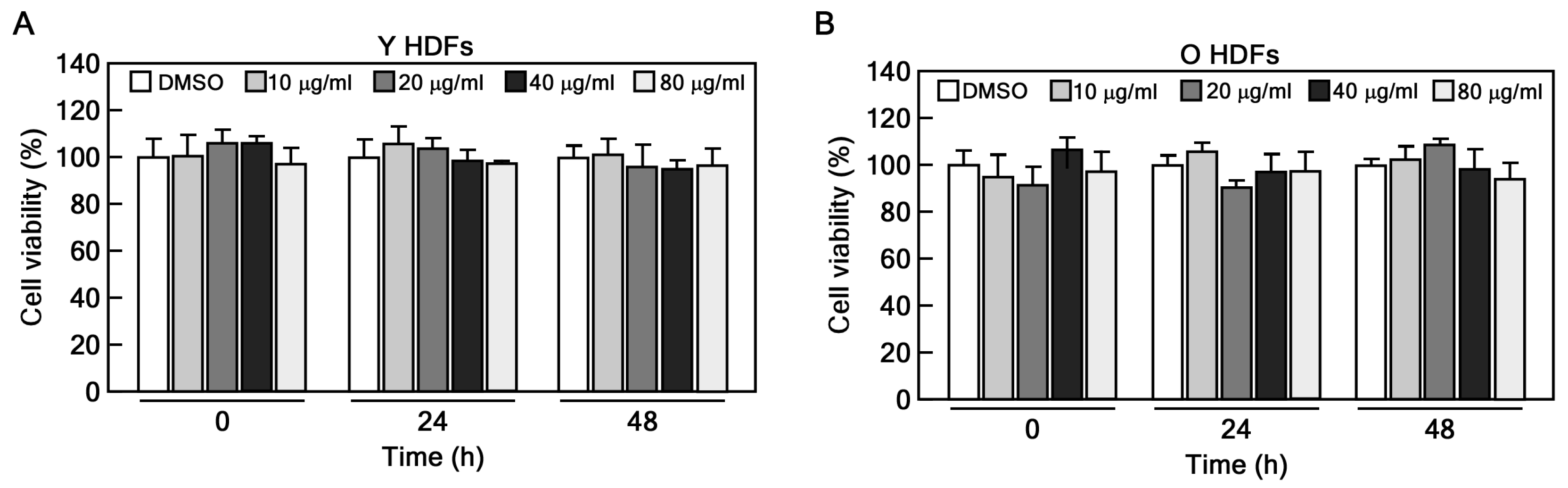
2.1. CHE Reverses the Cellular Senescence in Senescent HDFs
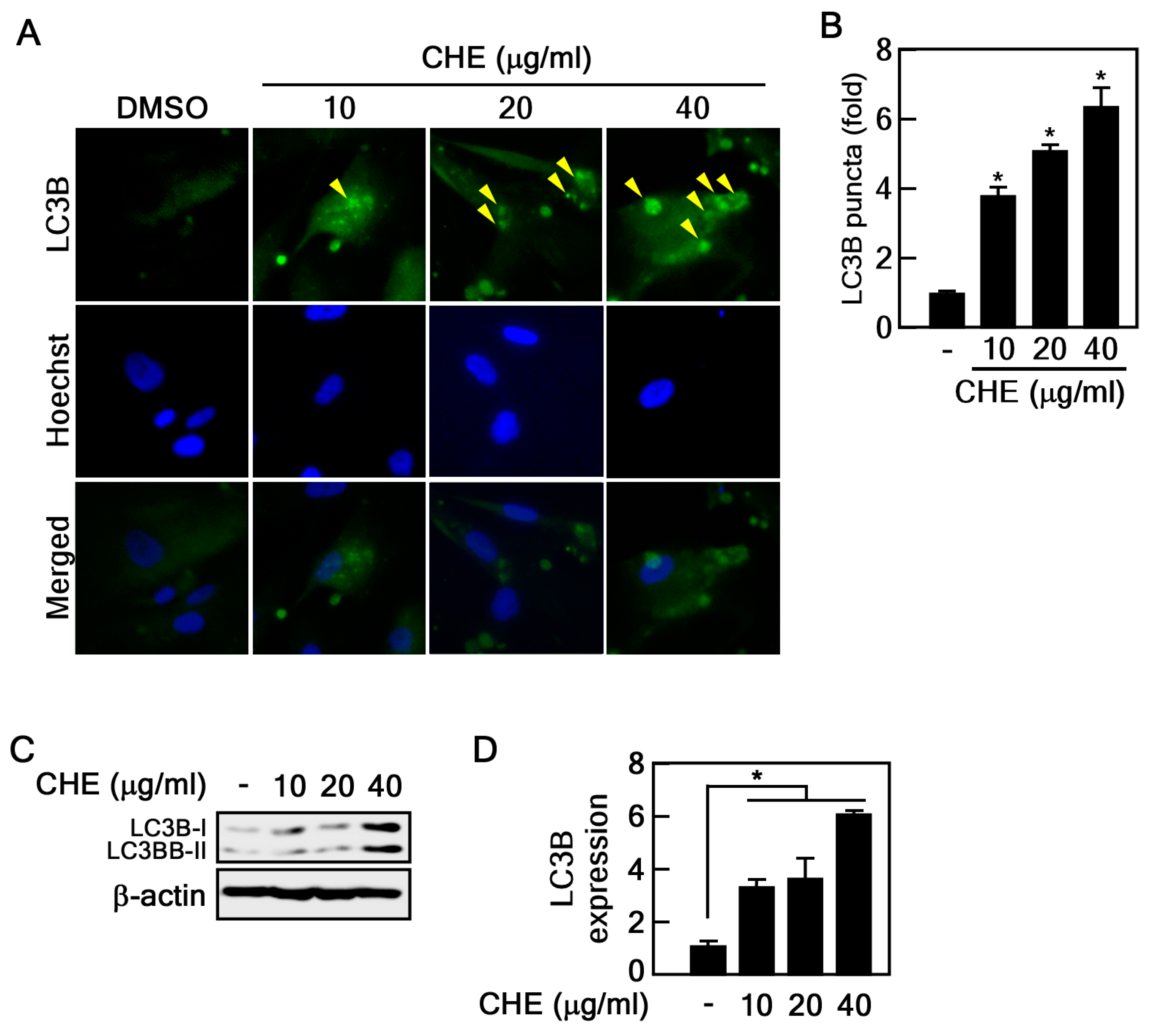
2.2. Regulation of Autophagy by CHE in Senescent HDFs
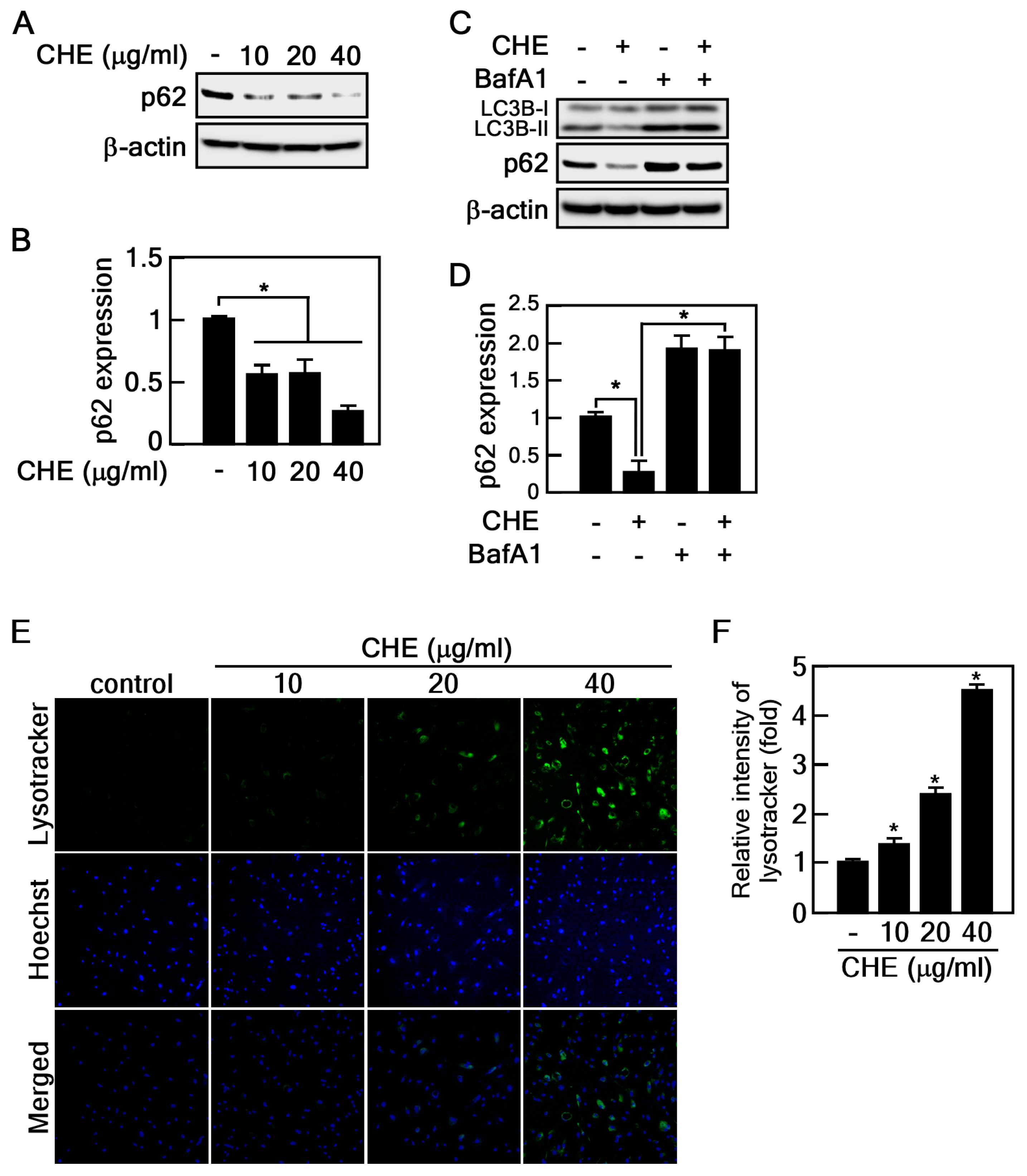
2.3. CHE Promotes Autophagic Flux
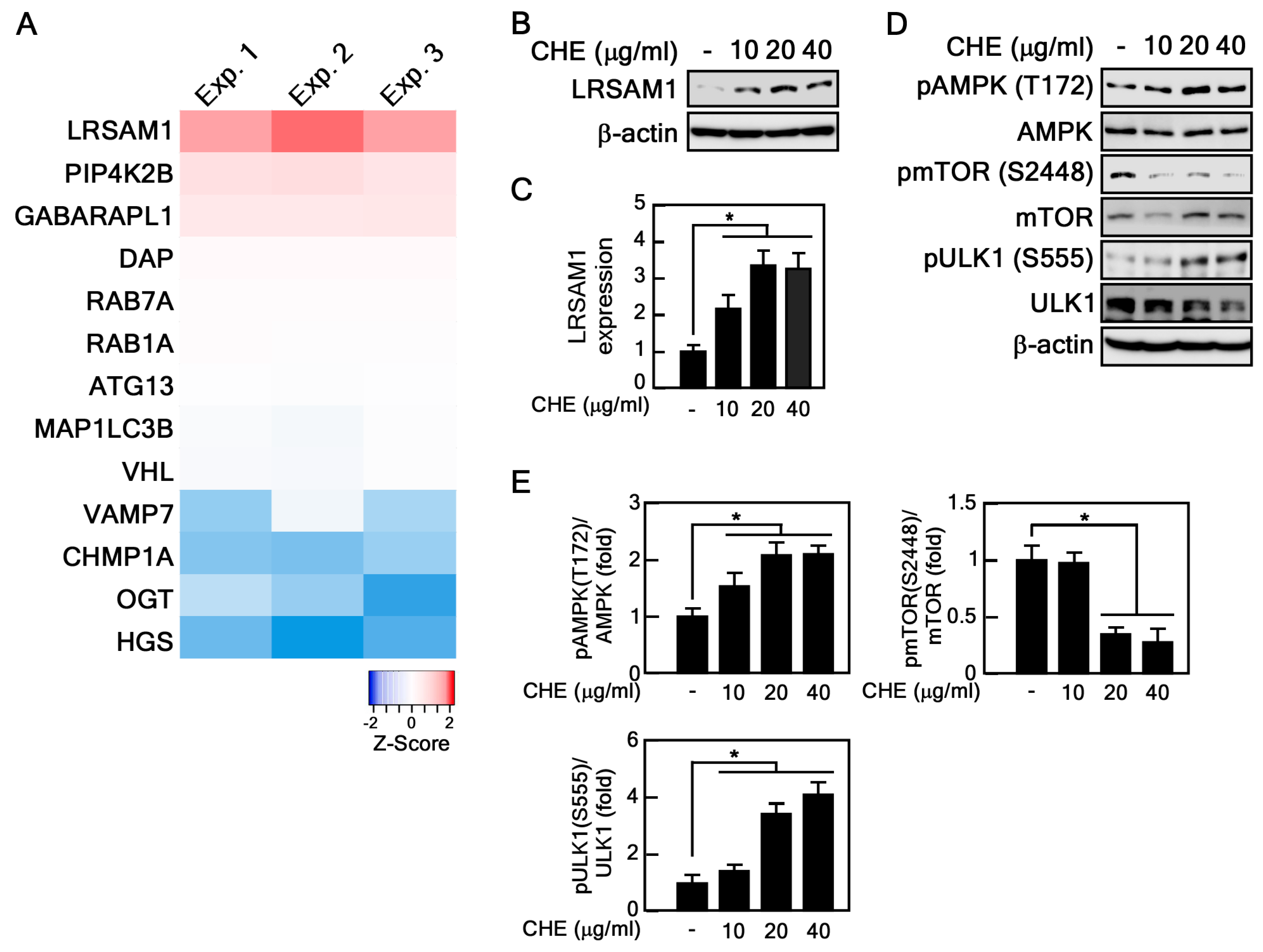
2.4. CHE Affects Leucine-Rich Repeat and Sterile Alpha Motif-Containing 1 (LRSAM1) Expression and Adenosine-Monophosphate Activated-Protein Kinase (AMPK)-Mammalian Target of Rapamycin (mTOR) Pathway
2.5. Depletion of LRSAM1 Suppresses the CHE-Induced Reversal of Cellular Senescence by Inhibiting Autophagy
3. Discussion
4. Materials and Methods
4.1. Corydalis Heterocarpa Extract
4.2. Cell Culture and Treatment
4.3. CCK-8 Assay
4.4. Senescence-Associated β-galactosidase Staining Assay
4.5. Western Blotting
4.6. LC3B Puncta Formation
4.7. Measuring Autophagy-Associated Lysosomal Activity
4.8. Identification of Compounds in CHE Using LC/MS
4.9. Cell Cycle Distribution Analysis
4.10. RNA-Sequencing
4.11. Materials
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ageing and Health. Available online: https://www.who.int/news-room/fact-sheets/detail/ageing-and-health (accessed on 1 January 2023).
- Kennedy, B.K.; Berger, S.L.; Brunet, A.; Campisi, J.; Cuervo, A.M.; Epel, E.S.; Franceschi, C.; Lithgow, G.J.; Morimoto, R.I.; Pessin, J.E.; et al. Geroscience: Linking aging to chronic disease. Cell 2014, 159, 709–713. [Google Scholar] [CrossRef]
- Guo, J.; Huang, X.; Dou, L.; Yan, M.; Shen, T.; Tang, W.; Li, J. Aging and aging-related diseases: From molecular mechanisms to interventions and treatments. Signal Transduct. Target. Ther. 2022, 7, 391. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Hayflick, L.; Moorhead, P.S. The serial cultivation of human diploid cell strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef]
- Beauséjour, C.M.; Krtolica, A.; Galimi, F.; Narita, M.; Lowe, S.W.; Yaswen, P.; Campisi, J. Reversal of human cellular senescence: Roles of the p53 and p16 pathways. EMBO J. 2003, 22, 4212–4222. [Google Scholar] [CrossRef]
- Liu, B.; Chen, Y.; St. Clair, D.K. ROS and p53: A versatile partnership. Free Radic. Biol. Med. 2008, 44, 1529–1535. [Google Scholar] [CrossRef]
- Serrano, M.; Lin, A.W.; McCurrach, M.E.; Beach, D.; Lowe, S.W. Oncogenic ras provokes premature cell senescence associated with accumulation of p53 and p16INK4a. Cell 1997, 88, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Segura, A.; Nehme, J.; Demaria, M. Hallmarks of Cellular Senescence. Trends Cell Biol. 2018, 28, 436–453. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.E.; Nam, S.-B.; Jang, M.; Park, J.; Lee, G.-E.; Cho, Y.-Y.; Jang, B.-C.; Lee, C.-J.; Choi, J.-S. Ginsenoside Rb2 suppresses cellular senescence of human dermal fibroblasts by inducing autophagy. J. Ginseng Res. 2023, 47, 337–346. [Google Scholar] [CrossRef]
- Cho, K.A.; Ryu, S.J.; Oh, Y.S.; Park, J.H.; Lee, J.W.; Kim, H.P.; Kim, K.T.; Jang, I.S.; Park, S.C. Morphological adjustment of senescent cells by modulating caveolin-1 status. J. Biol. Chem. 2004, 279, 42270–42278. [Google Scholar] [CrossRef]
- Zhang, J.; Lazarenko, O.P.; Blackburn, M.L.; Badger, T.M.; Ronis, M.J.J.; Chen, J.-R. Soy protein isolate down-regulates caveolin-1 expression to suppress osteoblastic cell senescence pathways. FASEB J. 2014, 28, 3134–3145. [Google Scholar] [CrossRef]
- Slobodnyuk, K.; Radic, N.; Ivanova, S.; Llado, A.; Trempolec, N.; Zorzano, A.; Nebreda, A.R. Autophagy-induced senescence is regulated by p38alpha signaling. Cell Death Dis. 2019, 10, 376. [Google Scholar] [CrossRef]
- Garcia-Prat, L.; Martinez-Vicente, M.; Perdiguero, E.; Ortet, L.; Rodriguez-Ubreva, J.; Rebollo, E.; Ruiz-Bonilla, V.; Gutarra, S.; Ballestar, E.; Serrano, A.L.; et al. Autophagy maintains stemness by preventing senescence. Nature 2016, 529, 37–42. [Google Scholar] [CrossRef]
- Chun, Y.; Kim, J. Autophagy: An Essential Degradation Program for Cellular Homeostasis and Life. Cells 2018, 7, 278. [Google Scholar] [CrossRef]
- Matecic, M.; Smith, D.L.; Pan, X.; Maqani, N.; Bekiranov, S.; Boeke, J.D.; Smith, J.S. A microarray-based genetic screen for yeast chronological aging factors. PLoS Genet. 2010, 6, e1000921. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.; Rubinsztein, D.C.; Walker, D.W. Autophagy as a promoter of longevity: Insights from model organisms. Nat. Rev. Mol. Cell Biol. 2018, 19, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Tóth, M.L.; Sigmond, T.; Borsos, E.; Barna, J.; Erdélyi, P.; Takács-Vellai, K.; Orosz, L.; Kovács, A.L.; Csikós, G.; Sass, M.; et al. Longevity pathways converge on autophagy genes to regulate life span in Caenorhabditis elegans. Autophagy 2008, 4, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Pyo, J.-O.; Yoo, S.-M.; Ahn, H.-H.; Nah, J.; Hong, S.-H.; Kam, T.-I.; Jung, S.; Jung, Y.-K. Overexpression of Atg5 in mice activates autophagy and extends lifespan. Nat. Commun. 2013, 4, 2300. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.N. Flora of Korea; Kyo-Hak Publishing: Seoul, Republic of Korea, 2002; pp. 218–219. [Google Scholar]
- Kim, Y.A.; Kong, C.-S.; Yea, S.S.; Seo, Y. Constituents of Corydalis heterocarpa and their anti-proliferative effects on human cancer cells. Food Chem. Toxicol. 2010, 48, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.-H.; Kong, C.-S.; Seo, Y.; Kim, M.-M.; Kim, S.-K. Anti-inflammatory effect of coumarins isolated from Corydalis heterocarpa in HT-29 human colon carcinoma cells. Food Chem. Toxicol. 2009, 47, 2129–2134. [Google Scholar] [CrossRef]
- Oh, J.H.; Karadeniz, F.; Seo, Y.; Kong, C.S. Hyunganol II Exerts Antiadipogenic Properties via MAPK-Mediated Suppression of PPARgamma Expression in Human Bone Marrow-Derived Mesenchymal Stromal Cells. Evid.-Based Complement. Altern. Med. 2022, 2022, 4252917. [Google Scholar] [CrossRef]
- Ahn, B.-N.; Kim, J.-A.; Kong, C.-S.; Seo, Y.; Kim, S.-K. Protective effect of (2′S)-columbianetin from Corydalis heterocarpa on UVB-induced keratinocyte damage. J. Photochem. Photobiol. B 2012, 109, 20–27. [Google Scholar] [CrossRef]
- Galbiati, F.; Volonte, D.; Liu, J.; Capozza, F.; Frank, P.G.; Zhu, L.; Pestell, R.G.; Lisanti, M.P. Caveolin-1 expression negatively regulates cell cycle progression by inducing G(0)/G(1) arrest via a p53/p21(WAF1/Cip1)-dependent mechanism. Mol. Biol. Cell 2001, 12, 2229–2244. [Google Scholar] [CrossRef]
- Wang, L.; Chen, M.; Yang, J.; Zhang, Z. LC3 fluorescent puncta in autophagosomes or in protein aggregates can be distinguished by FRAP analysis in living cells. Autophagy 2013, 9, 756–769. [Google Scholar] [CrossRef]
- Yoshii, S.R.; Mizushima, N. Monitoring and Measuring Autophagy. Int. J. Mol. Sci. 2017, 18, 1865. [Google Scholar] [CrossRef]
- Wang, Z.; Hu, J.; Li, G.; Qu, L.; He, Q.; Lou, Y.; Song, Q.; Ma, D.; Chen, Y. PHF23 (plant homeodomain finger protein 23) negatively regulates cell autophagy by promoting ubiquitination and degradation of E3 ligase LRSAM1. Autophagy 2014, 10, 2158–2170. [Google Scholar] [CrossRef]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K.-L. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Ahn, B.N.; Kim, J.A.; Kong, C.S.; Seo, Y.; Kim, S.K. Photoprotective effect of libanoridin isolated from Corydalis heterocarpa on UVB stressed human keratinocyte cells. Exp. Dermatol. 2013, 22, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.; Oresajo, C.; Hayward, J. Ultraviolet radiation and skin aging: Roles of reactive oxygen species, inflammation and protease activation, and strategies for prevention of inflammation-induced matrix degradation—A review. Int. J. Cosmet. Sci. 2005, 27, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, Y.; Zhao, Z.; Qiu, J. Oxidative stress in the skin: Impact and related protection. Int. J. Cosmet. Sci. 2021, 43, 495–509. [Google Scholar] [CrossRef] [PubMed]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef]
- Choi, S.J.; Lee, S.N.; Kim, K.; Joo, D.H.; Shin, S.; Lee, J.; Lee, H.K.; Kim, J.; Kwon, S.B.; Kim, M.J.; et al. Biological effects of rutin on skin aging. Int. J. Mol. Med. 2016, 38, 357–363. [Google Scholar] [CrossRef]
- Han, T.; Cheng, G.; Liu, Y.; Yang, H.; Hu, Y.-T.; Huang, W. In vitro evaluation of tectoridin, tectorigenin and tectorigenin sodium sulfonate on antioxidant properties. Food Chem. Toxicol. 2012, 50, 409–414. [Google Scholar] [CrossRef]
- Dong, T.; Fan, X.; Zheng, N.; Yan, K.; Hou, T.; Peng, L.; Ci, X. Activation of Nrf2 signalling pathway by tectoridin protects against ferroptosis in particulate matter-induced lung injury. Br. J. Pharmacol. 2023, 180, 2532–2549. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.-L.; Zou, L.-B.; Lin, S.; Shi, J.-G.; Zhu, H.-B. Anti-apoptotic effect of esculin on dopamine-induced cytotoxicity in the human neuroblastoma SH-SY5Y cell line. Neuropharmacology 2007, 53, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Shen, Y.; Du, Y.; Chen, J.; Pei, F.; Fu, W.; Qiao, J. Esculin prevents Lipopolysaccharide/D-Galactosamine-induced acute liver injury in mice. Microb. Pathog. 2018, 125, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Verdú, E.; Ceballos, D.; Vilches, J.J.; Navarro, X. Influence of aging on peripheral nerve function and regeneration. J. Peripher. Nerv. Syst. 2000, 5, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Yang, X.; Wang, L.; Zhang, Q.; Ma, W.; Huang, Z.; Bao, Y.; Zhong, L.; Sun, H.; Ding, F. Isoquercitrin promotes peripheral nerve regeneration through inhibiting oxidative stress following sciatic crush injury in mice. Ann. Transl. Med. 2019, 7, 680. [Google Scholar] [CrossRef] [PubMed]
- Shui, L.; Wang, W.; Xie, M.; Ye, B.; Li, X.; Liu, Y.; Zheng, M. Isoquercitrin induces apoptosis and autophagy in hepatocellular carcinoma cells via AMPK/mTOR/p70S6K signaling pathway. Aging 2020, 12, 24318–24332. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Retzlaff, M.; Roos, T.; Frydman, J. Cellular strategies of protein quality control. Cold Spring Harb. Perspect. Biol. 2011, 3, a004374. [Google Scholar] [CrossRef] [PubMed]
- Cheon, S.Y.; Kim, H.; Rubinsztein, D.C.; Lee, J.E. Autophagy, Cellular Aging and Age-related Human Diseases. Exp. Neurobiol. 2019, 28, 643–657. [Google Scholar] [CrossRef] [PubMed]
- Huett, A.; Heath, R.J.; Begun, J.; Sassi, S.O.; Baxt, L.A.; Vyas, J.M.; Goldberg, M.B.; Xavier, R.J. The LRR and RING domain protein LRSAM1 is an E3 ligase crucial for ubiquitin-dependent autophagy of intracellular Salmonella Typhimurium. Cell Host Microbe 2012, 12, 778–790. [Google Scholar] [CrossRef] [PubMed]
- Dubey, A.R.; Mishra, R.; Sundaria, N.; Jagtap, Y.A.; Kumar, P.; Kinger, S.; Choudhary, A.; Jha, H.C.; Prasad, A.; Gutti, R.K.; et al. Resveratrol Promotes LRSAM1 E3 Ubiquitin Ligase-Dependent Degradation of Misfolded Proteins Linked with Neurodegeneration. Cell Physiol. Biochem. 2022, 56, 530–545. [Google Scholar] [PubMed]
- Alers, S.; Löffler, A.S.; Wesselborg, S.; Stork, B. Role of AMPK-mTOR-Ulk1/2 in the regulation of autophagy: Cross talk, shortcuts, and feedbacks. Mol. Cell. Biol. 2012, 32, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Zachari, M.; Ganley, I.G. The mammalian ULK1 complex and autophagy initiation. Essays Biochem. 2017, 61, 585–596. [Google Scholar]
- Koren, I.; Reem, E.; Kimchi, A. DAP1, a novel substrate of mTOR, negatively regulates autophagy. Curr. Biol. 2010, 20, 1093–1098. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies | Company | Catalog |
|---|---|---|
| p53 | Santa Cruz Biotechnology | sc-126 |
| p21 | Cell Signaling Technology | 2946 |
| caveolin-1 | Cell Signaling Technology | 3238 |
| β-actin | Santa Cruz Biotechnology | sc-47778 |
| LC3B | Novus Biologicals | NB100-2220 |
| p62 | Novus Biologicals | NBP1-48320 |
| p-AMPK | Cell Signaling Technology | 2531 |
| AMPK | Cell Signaling Technology | 2532 |
| p-mTOR | Cell Signaling Technology | 5536 |
| mTOR | Cell Signaling Technology | 2972 |
| p-ULK1 | Cell Signaling Technology | 6888 |
| ULK1 | Cell Signaling Technology | 4773 |
| LRSAM1 | Cell Signaling Technology | 28405 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, K.E.; Nam, S.-B.; Lee, G.-E.; Yang, G.; Lee, M.-H.; Bang, G.; Choi, J.H.; Cho, Y.-Y.; Lee, C.-J. Induction of Autophagy by Extract from Corydalis heterocarpa for Skin Anti-Aging. Mar. Drugs 2024, 22, 127. https://doi.org/10.3390/md22030127
Yang KE, Nam S-B, Lee G-E, Yang G, Lee M-H, Bang G, Choi JH, Cho Y-Y, Lee C-J. Induction of Autophagy by Extract from Corydalis heterocarpa for Skin Anti-Aging. Marine Drugs. 2024; 22(3):127. https://doi.org/10.3390/md22030127
Chicago/Turabian StyleYang, Kyeong Eun, Soo-Bin Nam, Ga-Eun Lee, Gabsik Yang, Mee-Hyun Lee, Geul Bang, Jung Hoon Choi, Yong-Yeon Cho, and Cheol-Jung Lee. 2024. "Induction of Autophagy by Extract from Corydalis heterocarpa for Skin Anti-Aging" Marine Drugs 22, no. 3: 127. https://doi.org/10.3390/md22030127
APA StyleYang, K. E., Nam, S.-B., Lee, G.-E., Yang, G., Lee, M.-H., Bang, G., Choi, J. H., Cho, Y.-Y., & Lee, C.-J. (2024). Induction of Autophagy by Extract from Corydalis heterocarpa for Skin Anti-Aging. Marine Drugs, 22(3), 127. https://doi.org/10.3390/md22030127