
A Standardized Extract of Microalgae Phaeodactylum tricornutum (Mi136) Inhibit D-Gal Induced Cognitive Dysfunction in Mice

Abstract
:1. Introduction
2. Results
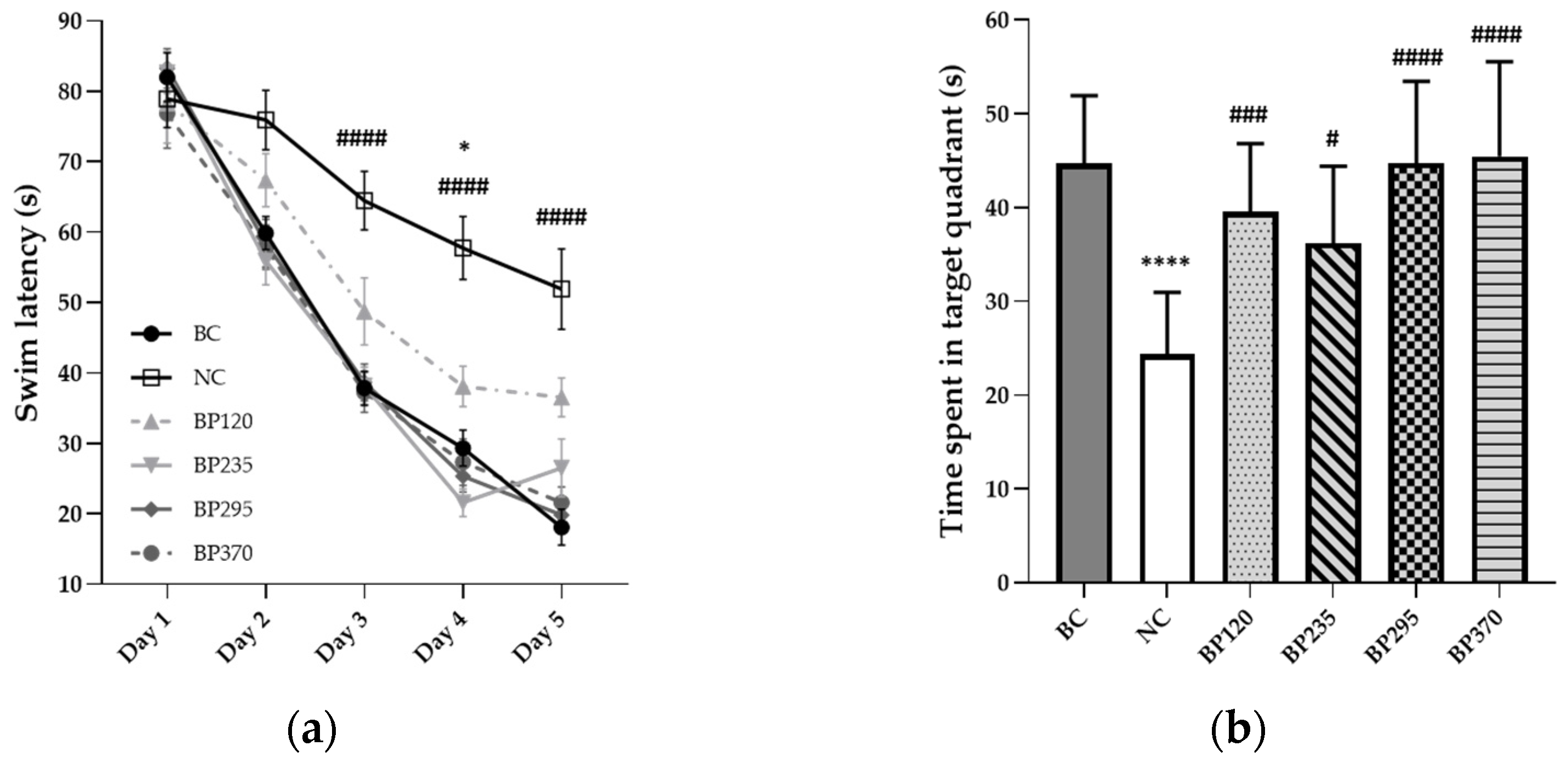
2.1. Effect on Y-Maze Spontaneous Alternation Performance
2.1.1. Y-Maze Spontaneous Alternation Performance
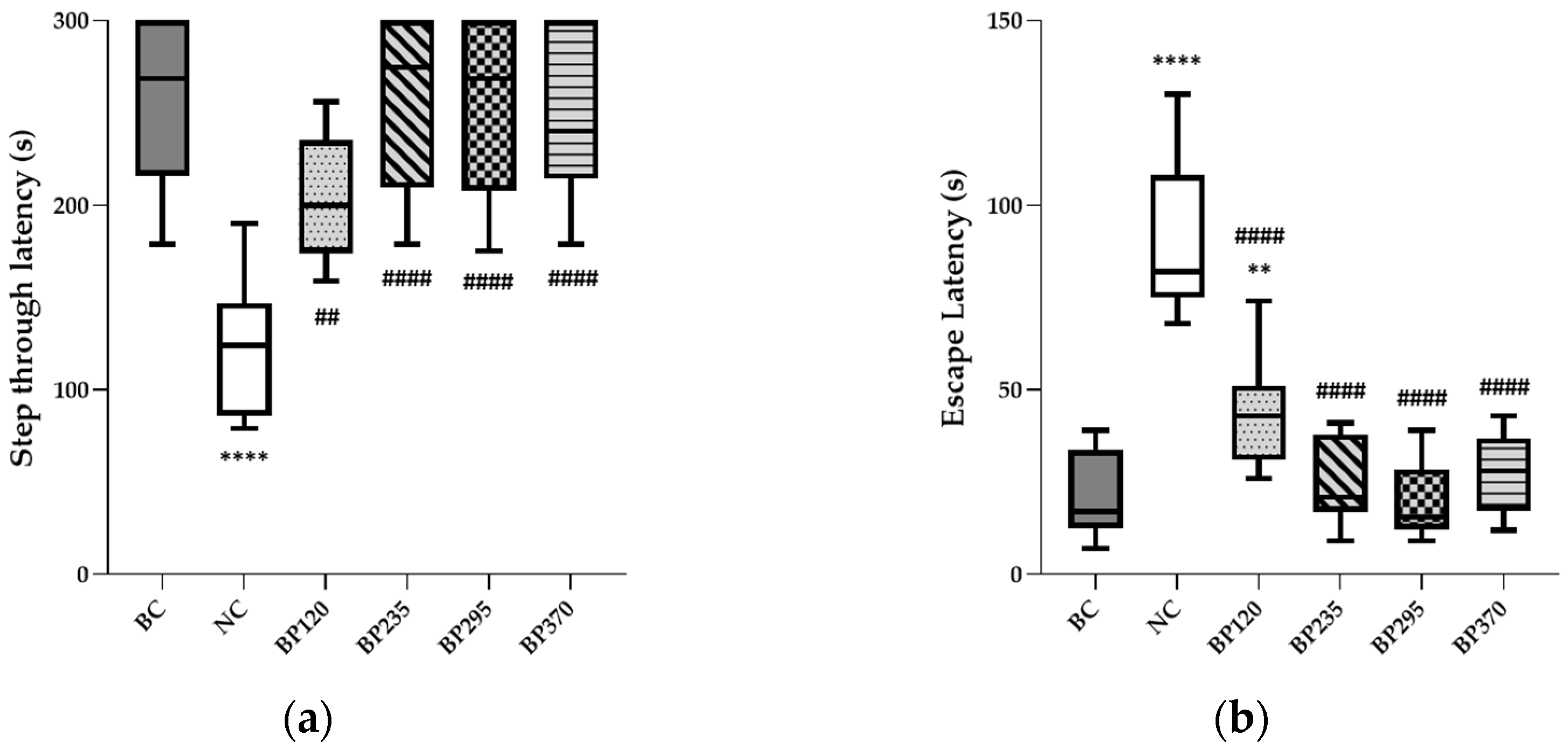
2.1.2. Place Learning in the Morris Water Maze
2.1.3. Passive Avoidance Test
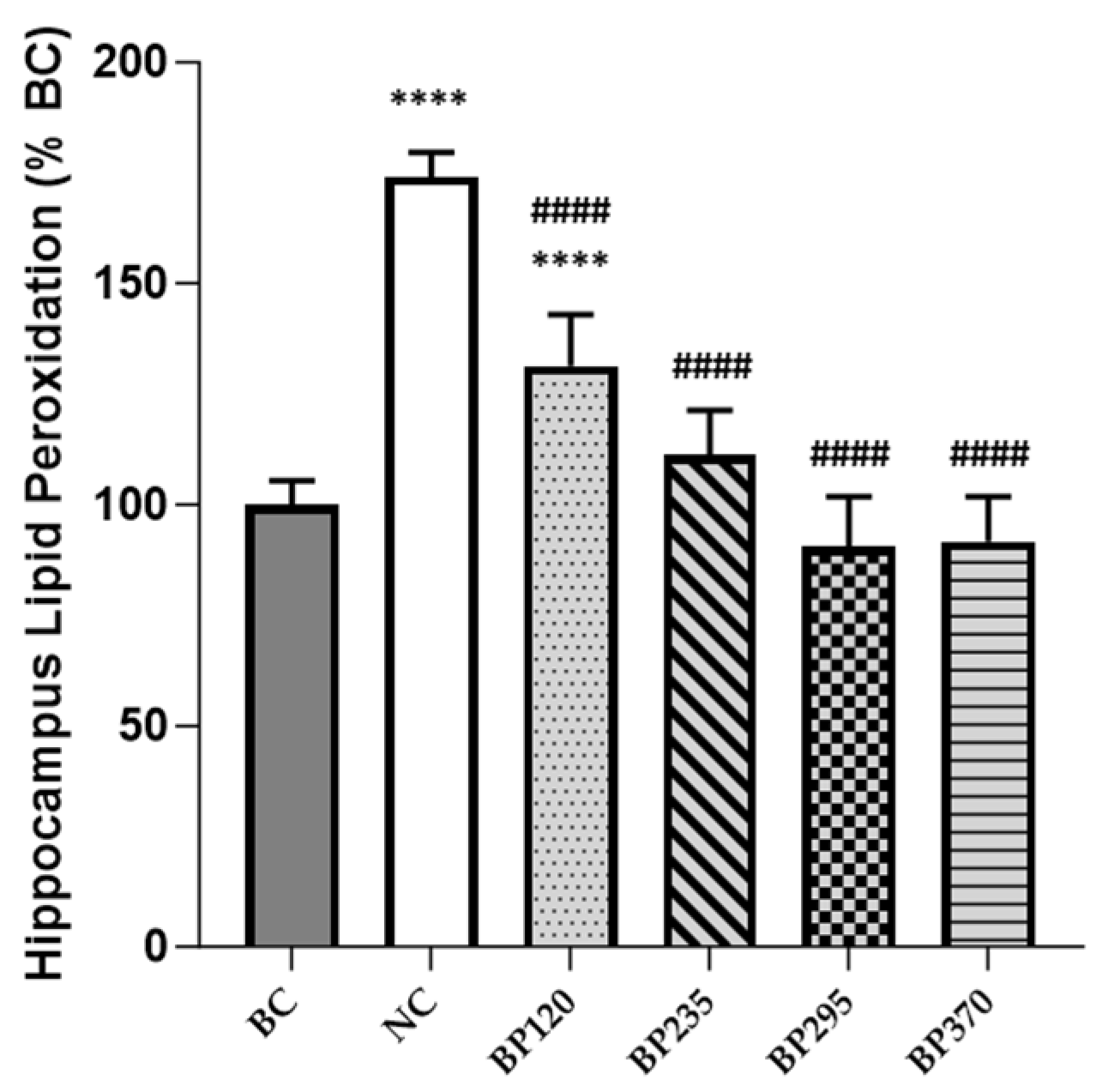
2.2. Effect on Brain Lipid Peroxidation Level
2.3. Effect on Brain and Plasma Inflammatory Markers Level
3. Discussion
4. Materials and Methods
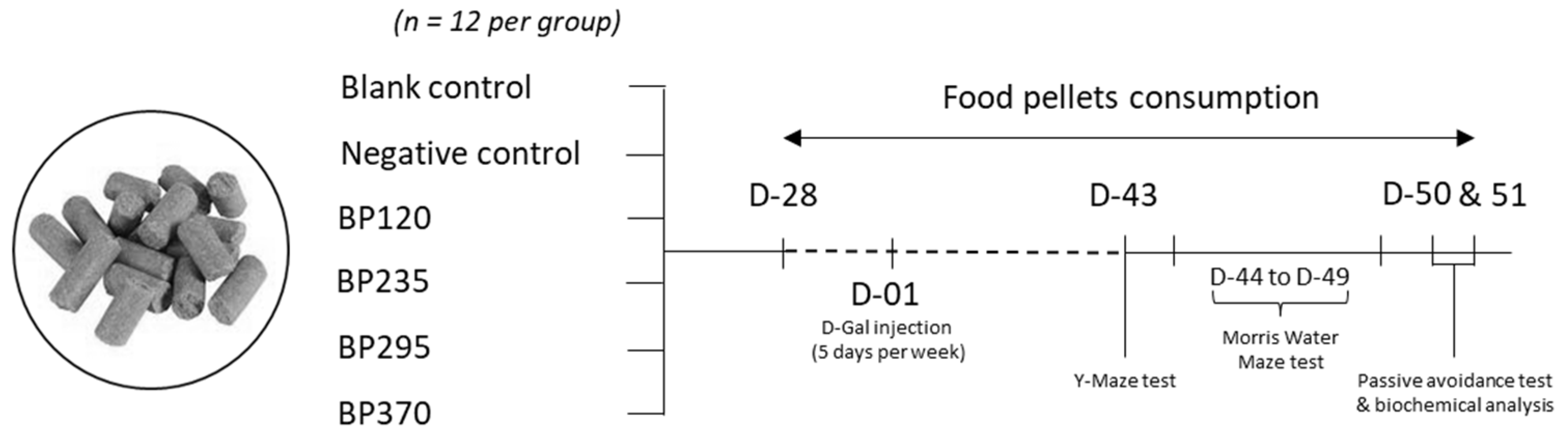
4.1. Animals, Diet and Treatment Groups
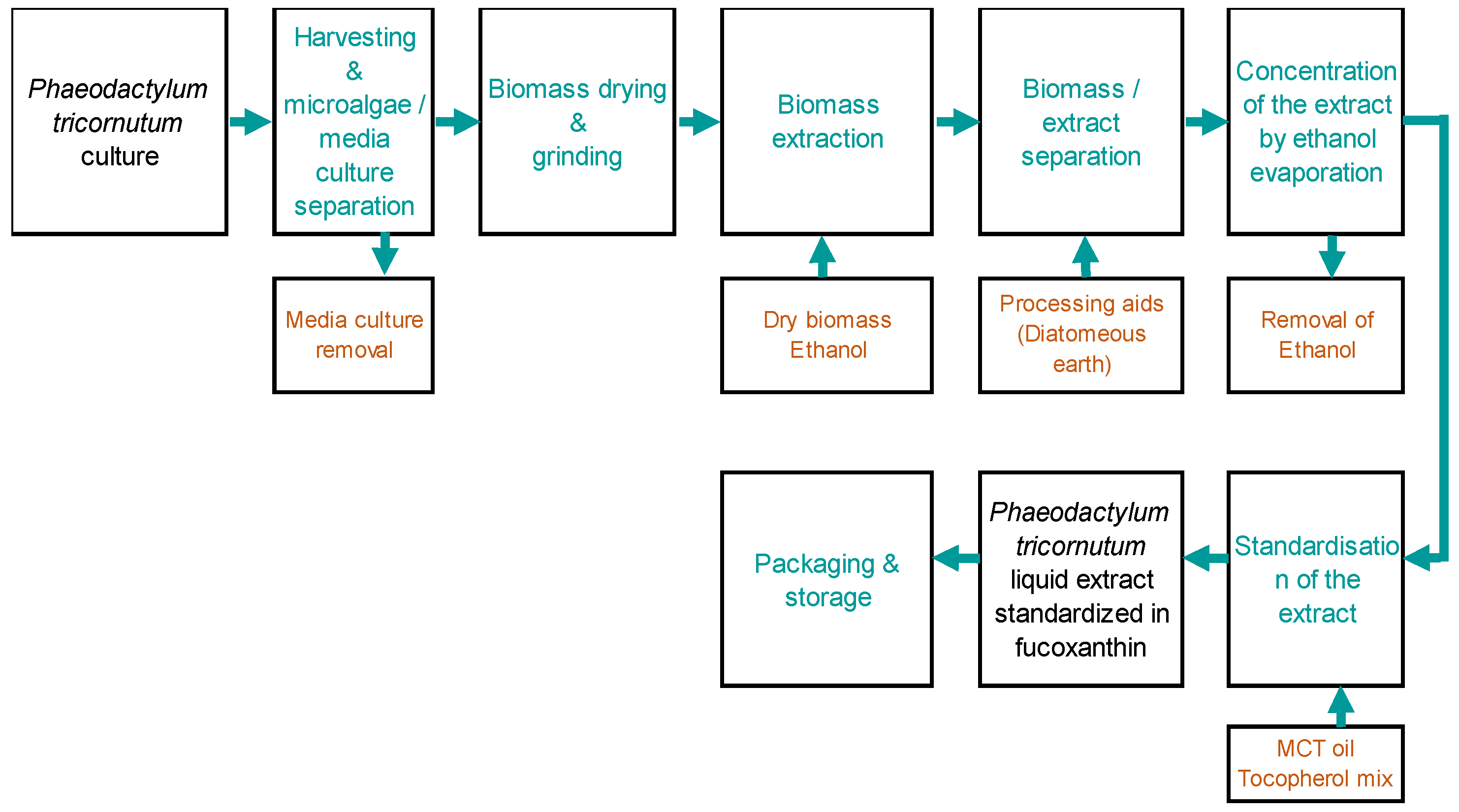
4.2. Microalgae Extract of PT Production and Characterization
4.3. Y-Maze Spontaneous Alternation Performance
4.4. Place Learning in the Morris Water Maze
4.5. Passive Avoidance Test
4.6. Biochemical Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- United Nations Department of Economic and Social Affairs. World Population Prospects, the 2012 Revision; Population division of the department of economic and social affairs of the United Nations Secretariat: New York, NY, USA, 2013. [Google Scholar]
- Salthouse, T. Consequences of Age-Related Cognitive Declines. Annu. Rev. Psychol. 2012, 63, 201–226. [Google Scholar] [CrossRef]
- Salthouse, T.A. When Does Age-Related Cognitive Decline Begin? Neurobiol. Aging 2009, 30, 507–514. [Google Scholar] [CrossRef]
- Singh-Manoux, A.; Kivimaki, M.; Glymour, M.M.; Elbaz, A.; Berr, C.; Ebmeier, K.P.; Ferrie, J.E.; Dugravot, A. Timing of Onset of Cognitive Decline: Results from Whitehall II Prospective Cohort Study. BMJ 2012, 344, d7622. [Google Scholar] [CrossRef]
- Hedden, T.; Gabrieli, J.D.E. Insights into the Ageing Mind: A View from Cognitive Neuroscience. Nat. Rev. Neurosci. 2004, 5, 87–96. [Google Scholar] [CrossRef]
- Freemantle, E.; Vandal, M.; Tremblay-Mercier, J.; Tremblay, S.; Blachère, J.-C.; Bégin, M.E.; Brenna, J.T.; Windust, A.; Cunnane, S.C. Omega-3 Fatty Acids, Energy Substrates, and Brain Function during Aging. Prostaglandins Leukot. Essent. Fat. Acids 2006, 75, 213–220. [Google Scholar] [CrossRef]
- Andlin-Sobocki, P.; Jönsson, B.; Wittchen, H.-U.; Olesen, J. Cost of Disorders of the Brain in Europe. Eur. J. Neurol. 2005, 12 (Suppl. S1), 1–27. [Google Scholar] [CrossRef]
- Harada, C.N.; Natelson Love, M.C.; Triebel, K.L. Normal Cognitive Aging. Clin. Geriatr. Med. 2013, 29, 737–752. [Google Scholar] [CrossRef] [PubMed]
- Scarmeas, N.; Anastasiou, C.A.; Yannakoulia, M. Nutrition and Prevention of Cognitive Impairment. Lancet Neurol. 2018, 17, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Tönnies, E.; Trushina, E. Oxidative Stress, Synaptic Dysfunction, and Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 57, 1105–1121. [Google Scholar] [CrossRef] [PubMed]
- McGrattan, A.M.; McGuinness, B.; McKinley, M.C.; Kee, F.; Passmore, P.; Woodside, J.V.; McEvoy, C.T. Diet and Inflammation in Cognitive Ageing and Alzheimer’s Disease. Curr. Nutr. Rep. 2019, 8, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Vorburger, R.; Scarmeas, N.; Luchsinger, J.A.; Manly, J.J.; Schupf, N.; Mayeux, R.; Brickman, A.M. Circulating Inflammatory Biomarkers in Relation to Brain Structural Measurements in a Non-Demented Elderly Population. Brain Behav. Immun. 2017, 65, 150–160. [Google Scholar] [CrossRef]
- Swomley, A.M.; Butterfield, D.A. Oxidative Stress in Alzheimer Disease and Mild Cognitive Impairment: Evidence from Human Data Provided by Redox Proteomics. Arch. Toxicol. 2015, 89, 1669–1680. [Google Scholar] [CrossRef] [PubMed]
- Valls-Pedret, C.; Sala-Vila, A.; Serra-Mir, M.; Corella, D.; de la Torre, R.; Martínez-González, M.Á.; Martínez-Lapiscina, E.H.; Fitó, M.; Pérez-Heras, A.; Salas-Salvadó, J.; et al. Mediterranean Diet and Age-Related Cognitive Decline: A Randomized Clinical Trial. JAMA Intern. Med. 2015, 175, 1094–1103. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.J.; Katuwal, S.; Anderson, G.A.; Gu, L.; Zhou, R.; Gibbons, W.R. Photobioreactor Cultivation Strategies for Microalgae and Cyanobacteria. Biotechnol. Prog. 2018, 34, 811–827. [Google Scholar] [CrossRef] [PubMed]
- Delbrut, A.; Albina, P.; Lapierre, T.; Pradelles, R.; Dubreucq, E. Fucoxanthin and Polyunsaturated Fatty Acids Co-Extraction by a Green Process. Molecules 2018, 23, E874. [Google Scholar] [CrossRef] [PubMed]
- Salem, N.; Vandal, M.; Calon, F. The Benefit of Docosahexaenoic Acid for the Adult Brain in Aging and Dementia. Prostaglandins Leukot. Essent. Fat. Acids 2015, 92, 15–22. [Google Scholar] [CrossRef]
- Yurko-Mauro, K.; McCarthy, D.; Rom, D.; Nelson, E.B.; Ryan, A.S.; Blackwell, A.; Salem, N.; Stedman, M.; MIDAS Investigators. Beneficial Effects of Docosahexaenoic Acid on Cognition in Age-Related Cognitive Decline. Alzheimer’s Dement. 2010, 6, 456–464. [Google Scholar] [CrossRef]
- Aziz, E.; Batool, R.; Akhtar, W.; Rehman, S.; Shahzad, T.; Malik, A.; Shariati, M.A.; Laishevtcev, A.; Plygun, S.; Heydari, M.; et al. Xanthophyll: Health Benefits and Therapeutic Insights. Life Sci. 2020, 240, 117104. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Chen, W.; Tian, F.; Yuan, C.; Wang, H.; Yue, H. Neuroprotective Role of Fucoxanthin against Cerebral Ischemic/Reperfusion Injury through Activation of Nrf2/HO-1 Signaling. Biomed. Pharmacother. 2018, 106, 1484–1489. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Chen, L.; Cheng, W.; Liu, T. Combined Production of Fucoxanthin and EPA from Two Diatom Strains Phaeodactylum Tricornutum and Cylindrotheca Fusiformis Cultures. Bioprocess Biosyst. Eng. 2018, 41, 1061–1071. [Google Scholar] [CrossRef]
- Minghetti, L.; Salvi, R.; Lavinia Salvatori, M.; Ajmone-Cat, M.A.; De Nuccio, C.; Visentin, S.; Bultel-Poncé, V.; Oger, C.; Guy, A.; Galano, J.-M.; et al. Nonenzymatic Oxygenated Metabolites of α-Linolenic Acid B1- and L1-Phytoprostanes Protect Immature Neurons from Oxidant Injury and Promote Differentiation of Oligodendrocyte Progenitors through PPAR-γ Activation. Free Radic. Biol. Med. 2014, 73, 41–50. [Google Scholar] [CrossRef]
- Fotuhi, M.; Mohassel, P.; Yaffe, K. Fish Consumption, Long-Chain Omega-3 Fatty Acids and Risk of Cognitive Decline or Alzheimer Disease: A Complex Association. Nat. Clin. Pract. Neurol. 2009, 5, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Dyall, S.C. Long-Chain Omega-3 Fatty Acids and the Brain: A Review of the Independent and Shared Effects of EPA, DPA and DHA. Front. Aging Neurosci. 2015, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Li, L.; Song, Q.; Ai, H.; Chu, J.; Li, W. Behavioural Study of the D-Galactose Induced Aging Model in C57BL/6J Mice. Behav. Brain Res. 2005, 157, 245–251. [Google Scholar] [CrossRef]
- Wu, D.; Lu, J.; Zheng, Y.; Zhou, Z.; Shan, Q.; Ma, D. Purple Sweet Potato Color Repairs D-Galactose-Induced Spatial Learning and Memory Impairment by Regulating the Expression of Synaptic Proteins. Neurobiol. Learn. Mem. 2008, 90, 19–27. [Google Scholar] [CrossRef]
- Grimmig, B.; Hudson, C.; Moss, L.; Peters, M.; Subbarayan, M.; Weeber, E.J.; Bickford, P.C. Astaxanthin Supplementation Modulates Cognitive Function and Synaptic Plasticity in Young and Aged Mice. Geroscience 2019, 41, 77–87. [Google Scholar] [CrossRef]
- Yook, J.S.; Okamoto, M.; Rakwal, R.; Shibato, J.; Lee, M.C.; Matsui, T.; Chang, H.; Cho, J.Y.; Soya, H. Astaxanthin Supplementation Enhances Adult Hippocampal Neurogenesis and Spatial Memory in Mice. Mol. Nutr. Food Res. 2016, 60, 589–599. [Google Scholar] [CrossRef]
- Farr, S.A.; Niehoff, M.L.; Ceddia, M.A.; Herrlinger, K.A.; Lewis, B.J.; Feng, S.; Welleford, A.; Butterfield, D.A.; Morley, J.E. Effect of Botanical Extracts Containing Carnosic Acid or Rosmarinic Acid on Learning and Memory in SAMP8 Mice. Physiol. Behav. 2016, 165, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Abdelmeguid, N.E.; Khalil, M.I.M.; Elhabet, R.; Sultan, A.S.; Salam, S.A. Combination of Docosahexaenoic Acid and Ginko Biloba Extract Improves Cognitive Function and Hippocampal Tissue Damages in a Mouse Model of Alzheimer’s Disease. J. Chem. Neuroanat. 2021, 116, 101995. [Google Scholar] [CrossRef]
- Ren, Q.; Sun, J.; Xu, D.; Xie, H.; Ye, M.; Zhao, Y. A Dietary Supplement Containing Micronutrients, Phosphatidylserine, and Docosahexaenoic Acid Counteracts Cognitive Impairment in D-Galactose-Induced Aged Rats. Front. Nutr. 2022, 9, 931734. [Google Scholar] [CrossRef]
- Wei, H.; Gao, Z.; Zheng, L.; Zhang, C.; Liu, Z.; Yang, Y.; Teng, H.; Hou, L.; Yin, Y.; Zou, X. Protective Effects of Fucoidan on Aβ25–35 and d-Gal-Induced Neurotoxicity in PC12 Cells and d-Gal-Induced Cognitive Dysfunction in Mice. Mar. Drugs 2017, 15, 77. [Google Scholar] [CrossRef] [PubMed]
- Sorrenti, V.; Castagna, D.A.; Fortinguerra, S.; Buriani, A.; Scapagnini, G.; Willcox, D.C. Spirulina Microalgae and Brain Health: A Scoping Review of Experimental and Clinical Evidence. Mar. Drugs 2021, 19, 293. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, X.; Xiao, J.; Song, M.; Cao, Y.; Xiao, H.; Liu, X. Astaxanthin Attenuates D-Galactose-Induced Brain Aging in Rats by Ameliorating Oxidative Stress, Mitochondrial Dysfunction, and Regulating Metabolic Markers. Food Funct. 2020, 11, 4103–4113. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Xuan, Z.; Wang, Q.; Yan, S.; Zhou, D.; Naman, C.B.; Zhang, J.; He, S.; Yan, X.; Cui, W. Fucoxanthin Has Potential for Therapeutic Efficacy in Neurodegenerative Disorders by Acting on Multiple Targets. Nutr. Neurosci. 2022, 25, 2167–2180. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef]
- Andronie-Cioara, F.L.; Ardelean, A.I.; Nistor-Cseppento, C.D.; Jurcau, A.; Jurcau, M.C.; Pascalau, N.; Marcu, F. Molecular Mechanisms of Neuroinflammation in Aging and Alzheimer’s Disease Progression. Int. J. Mol. Sci. 2023, 24, 1869. [Google Scholar] [CrossRef]
- Peters, M.M.; Hill, K.E.; Burk, R.F.; Weeber, E.J. Altered Hippocampus Synaptic Function in Selenoprotein P Deficient Mice. Mol. Neurodegener. 2006, 1, 12. [Google Scholar] [CrossRef]
- Papp, L.; Vizi, E.S.; Sperlágh, B. P2X7 Receptor Mediated Phosphorylation of p38MAP Kinase in the Hippocampus. Biochem. Biophys. Res. Commun. 2007, 355, 568–574. [Google Scholar] [CrossRef]
- Sanada, F.; Taniyama, Y.; Muratsu, J.; Otsu, R.; Shimizu, H.; Rakugi, H.; Morishita, R. Source of Chronic Inflammation in Aging. Front. Cardiovasc. Med. 2018, 5, 12. [Google Scholar] [CrossRef]
- Kobylarek, D.; Iwanowski, P.; Lewandowska, Z.; Limphaibool, N.; Szafranek, S.; Labrzycka, A.; Kozubski, W. Advances in the Potential Biomarkers of Epilepsy. Front. Neurol. 2019, 10, 685. [Google Scholar] [CrossRef]
- Koyama, A.; O’Brien, J.; Weuve, J.; Blacker, D.; Metti, A.L.; Yaffe, K. The Role of Peripheral Inflammatory Markers in Dementia and Alzheimer’s Disease: A Meta-Analysis. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 433–440. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 Fatty Acids and Inflammatory Processes: From Molecules to Man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, H.; Fan, Y.; Gao, Y.; Li, X.; Hu, Z.; Ding, K.; Wang, Y.; Wang, X. Fucoxanthin Provides Neuroprotection in Models of Traumatic Brain Injury via the Nrf2-ARE and Nrf2-Autophagy Pathways. Sci. Rep. 2017, 7, 46763. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Liu, F.; Lin, J.; Chen, H.; Huang, C.; Chen, L.; Zhou, Y.; Ye, L.; Zhang, K.; Jin, J.; et al. Fucoxanthin Inhibits β-Amyloid Assembly and Attenuates β-Amyloid Oligomer-Induced Cognitive Impairments. J. Agric. Food Chem. 2017, 65, 4092–4102. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-N.; Heo, S.-J.; Yoon, W.-J.; Kang, S.-M.; Ahn, G.; Yi, T.-H.; Jeon, Y.-J. Fucoxanthin Inhibits the Inflammatory Response by Suppressing the Activation of NF-κB and MAPKs in Lipopolysaccharide-Induced RAW 264.7 Macrophages. Eur. J. Pharmacol. 2010, 649, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Subhramanyam, C.S.; Wang, C.; Hu, Q.; Dheen, S.T. Microglia-Mediated Neuroinflammation in Neurodegenerative Diseases. Semin. Cell Dev. Biol. 2019, 94, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Dighriri, I.M.; Alsubaie, A.M.; Hakami, F.M.; Hamithi, D.M.; Alshekh, M.M.; Khobrani, F.A.; Dalak, F.E.; Hakami, A.A.; Alsueaadi, E.H.; Alsaawi, L.S.; et al. Effects of Omega-3 Polyunsaturated Fatty Acids on Brain Functions: A Systematic Review. Cureus 2022, 14, e30091. [Google Scholar] [CrossRef] [PubMed]
- Leonard, M.; Maury, J.; Dickerson, B.; Gonzalez, D.E.; Kendra, J.; Jenkins, V.; Nottingham, K.; Yoo, C.; Xing, D.; Ko, J.; et al. Effects of Dietary Supplementation of a Microalgae Extract Containing Fucoxanthin Combined with Guarana on Cognitive Function and Gaming Performance. Nutrients 2023, 15, 1918. [Google Scholar] [CrossRef]
- Fakhri, S.; Yarmohammadi, A.; Yarmohammadi, M.; Farzaei, M.H.; Echeverria, J. Marine Natural Products: Promising Candidates in the Modulation of Gut-Brain Axis towards Neuroprotection. Mar. Drugs 2021, 19, 165. [Google Scholar] [CrossRef] [PubMed]
- Mou, Y.; Du, Y.; Zhou, L.; Yue, J.; Hu, X.; Liu, Y.; Chen, S.; Lin, X.; Zhang, G.; Xiao, H.; et al. Gut Microbiota Interact with the Brain Through Systemic Chronic Inflammation: Implications on Neuroinflammation, Neurodegeneration, and Aging. Front. Immunol. 2022, 13, 796288. [Google Scholar] [CrossRef]
- Hiramatsu, M.; Inoue, K. Nociceptin/Orphanin FQ and Nocistatin on Learning and Memory Impairment Induced by Scopolamine in Mice. Br. J. Pharmacol. 1999, 127, 655–660. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M.T. Morris Water Maze: Procedures for Assessing Spatial and Related Forms of Learning and Memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Othman, M.Z.; Hassan, Z.; Che Has, A.T. Morris Water Maze: A Versatile and Pertinent Tool for Assessing Spatial Learning and Memory. Exp. Anim. 2022, 71, 264–280. [Google Scholar] [CrossRef] [PubMed]
- D’Hooge, R.; De Deyn, P.P. Applications of the Morris Water Maze in the Study of Learning and Memory. Brain Res. Rev. 2001, 36, 60–90. [Google Scholar] [CrossRef] [PubMed]
- Labban, S.; Alghamdi, B.S.; Alshehri, F.S.; Kurdi, M. Effects of Melatonin and Resveratrol on Recognition Memory and Passive Avoidance Performance in a Mouse Model of Alzheimer’s Disease. Behav. Brain Res. 2021, 402, 113100. [Google Scholar] [CrossRef] [PubMed]
- Tabrizian, K.; Yazdani, A.; Baheri, B.; Payandemehr, B.; Sanati, M.; Hashemzaei, M.; Miri, A.; Zandkarimi, M.; Belaran, M.; Fanoudi, S.; et al. Zinc Chloride and Lead Acetate-Induced Passive Avoidance Memory Retention Deficits Reversed by Nicotine and Bucladesine in Mice. Biol. Trace Elem. Res. 2016, 169, 106–113. [Google Scholar] [CrossRef]
- Hermes-Lima, M.; Willmore, W.G.; Storey, K.B. Quantification of Lipid Peroxidation in Tissue Extracts Based on Fe(III)Xylenol Orange Complex Formation. Free Radic. Biol. Med. 1995, 19, 271–280. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition (%) | |
|---|---|
| Phaeodactylum tricornutum extract | 40–70 (w:w) |
| MCT oil based on coconut oil | 30–60 (w:w) |
| Mix on non-GMO tocopherols | 0.45–0.55 (w:w) |
| Total lipids | 60–90 (w:w) |
| Proteins | 5–15 (w:w) |
| Humidity | <2 (w:w) |
| Ashes | <10 (w:w) |
| Carbohydrates | 0.5–20 (w:w) |
| Total PUFAs-w3 | ≥4 (w:w) |
| All-trans-fucoxanthin | 2.0 ± 0.4% (w:w) |
| Concentration | |
|---|---|
| Chlorophylls- g equivalent chloropyll A/100 g) | |
| Chlorophyll c | 3.46 ± 0.45 |
| Σ Chlorophyll b | ND |
| Σ Chlorophyll a | 0.24 ± 0.25 |
| Total carotenoids | |
| Β-carotene - g equivalent Β-carotene/100 g | 0.09 ± 0.08 |
| All trans Fucoxanthin - g equivalent fucoxanthin/100 g | 1.99 ± 1.01 |
| Total unsaturated fatty acids omega 3—g/100 g of product | 6.51 ± 1.17 |
| Eicosapentaenoic acid (EPA) | 7.99 ± 6.14 |
| Docosahexaenoic acid (DHA) | 0.23 ± 0.20 |
| Alpha-linolenic acid (ALA) | 0.24 ± 0.17 |
| Stearidonic acid (SDA) | 0.07 ± 0.08 |
| Total unsaturated fatty acids omega 6—g/100 g of product | 2.56 ± 0.19 |
| Hexadecadienoic acid | 2.01 ± 1.38 |
| Linoleic acid | 0.83 ± 0.66 |
| Arachidonic acid | 0.52 ± 0.44 |
| Eicosatetraenoic acid | 0.14 ± 0.12 |
| Total saturated fatty acids—g/100 g of product | 45.42 ± 8.49 |
| Caprylic acid | 23.24 ± 5.35 |
| Capric acid | 18.23 ± 4.06 |
| Palmitic acid | 2.53 ± 0.60 |
| Myristic acid | 0.90 ± 0.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maury, J.; Delbrut, A.; Villard, V.; Pradelles, R. A Standardized Extract of Microalgae Phaeodactylum tricornutum (Mi136) Inhibit D-Gal Induced Cognitive Dysfunction in Mice. Mar. Drugs 2024, 22, 99. https://doi.org/10.3390/md22030099
Maury J, Delbrut A, Villard V, Pradelles R. A Standardized Extract of Microalgae Phaeodactylum tricornutum (Mi136) Inhibit D-Gal Induced Cognitive Dysfunction in Mice. Marine Drugs. 2024; 22(3):99. https://doi.org/10.3390/md22030099
Chicago/Turabian StyleMaury, Jonathan, Antoine Delbrut, Vanessa Villard, and Rémi Pradelles. 2024. "A Standardized Extract of Microalgae Phaeodactylum tricornutum (Mi136) Inhibit D-Gal Induced Cognitive Dysfunction in Mice" Marine Drugs 22, no. 3: 99. https://doi.org/10.3390/md22030099
APA StyleMaury, J., Delbrut, A., Villard, V., & Pradelles, R. (2024). A Standardized Extract of Microalgae Phaeodactylum tricornutum (Mi136) Inhibit D-Gal Induced Cognitive Dysfunction in Mice. Marine Drugs, 22(3), 99. https://doi.org/10.3390/md22030099