
Protein Hydrolysis as a Way to Valorise Squid-Processing Byproducts: Obtaining and Identification of ACE, DPP-IV and PEP Inhibitory Peptides



Abstract
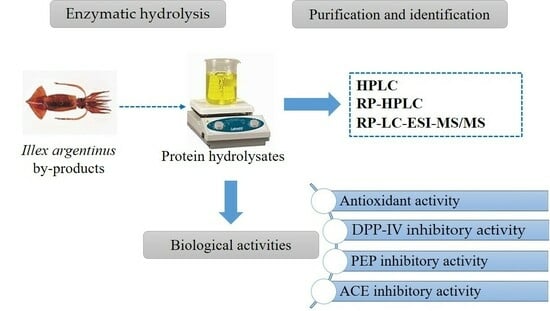
:
1. Introduction
2. Results and Discussion
2.1. Proximate Composition of the Squid Skin and Amino Acid Composition
2.2. Degree of Hydrolysis (DH) and Molecular Weight Distribution
2.3. Biological Properties of Protein Hydrolysates
2.3.1. Angiotensin-I-Converting Enzyme Inhibitory Activity
2.3.2. DPP-IV Inhibitory Activity
2.3.3. PEP Inhibitory Activity
2.3.4. Antioxidant Activity
2.4. Purification and Biological Activities of Fractions
2.5. Identification of the Peptide Composition of the Most Potent ACE-Inhibiting Fractions
2.6. Identification of the Peptide Composition of the Most Potent DPP-IV-Inhibiting Fractions
2.7. Identification of the Peptide Composition of the Most Potent PEP-Inhibiting Fractions
3. Materials and Methods
3.1. Enzyme and Chemicals
3.2. Raw Material
3.3. Determination of the Proximate Composition of Squid Skin
3.4. Enzymatic Hydrolysis
3.4.1. Measurement of Enzymatic Activity
3.4.2. Preparation of Protein Hydrolysates
3.5. Determination of the Degree of Hydrolysis
3.6. Molecular Weight Analysis
3.7. Biological Activities
3.7.1. ABTS Free-Radical-Scavenging Activity
3.7.2. Ferric-Reducing Power Assay (FRAP)
3.7.3. Angiotensin-I-Converting Enzyme Inhibitory Activity
3.7.4. DPP-IV Inhibitory Activity
3.7.5. Prolyl Endopeptidase Inhibitory Activity
3.8. Fractionation of the Protein Hydrolysates
3.9. Peptide Identification
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Navarro-Peraza, R.S.; Osuna-Ruiz, I.; Lugo-Sánchez, M.E.; Pacheco-Aguilar, R.; Ramírez-Suárez, J.C.; Burgos-Hernández, A.; Martínez-Montaño, E.; Salazar-Leyva, J.A. Structural and Biological Properties of Protein Hydrolysates from Seafood By-Products: A Review Focused on Fishery Effluents. Food Sci. Technol. 2020, 40, 1–5. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; The State of World Fisheries and Aquaculture (SOFIA); FAO: Rome, Italy, 2020; ISBN 978-92-5-132692-3. [Google Scholar]
- Cipriani, P.; Giulietti, L.; Palomba, M.; Mattiucci, S.; Bao, M.; Levsen, A. Occurrence of Larval Ascaridoid Nematodes in the Argentinean Short-Finned Squid Illex Argentinus from the Southwest Atlantic Ocean (off Falkland Islands). Int. J. Food Microbiol. 2019, 297, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Ezquerra-Brauer, J.M.; Aubourg, S.P. Recent Trends for the Employment of Jumbo Squid (Dosidicus gigas) by-Products as a Source of Bioactive Compounds with Nutritional, Functional and Preservative Applications: A Review. Int. J. Food Sci. Technol. 2019, 54, 987–998. [Google Scholar] [CrossRef]
- Camargo, T.R.; Mantoan, P.; Ramos, P.; Monserrat, J.M.; Prentice, C.; Fernandes, C.C.; Zambuzzi, W.F.; Valenti, W.C. Bioactivity of the Protein Hydrolysates Obtained from the Most Abundant Crustacean Bycatch. Mar. Biotechnol. 2021, 23, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Pratama, I.S.; Putra, Y.; Pangestuti, R.; Kim, S.-K.; Siahaan, E.A. Bioactive Peptides-Derived from Marine by-Products: Development, Health Benefits and Potential Application in Biomedicine. Fish. Aquat. Sci. 2022, 25, 357–379. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Jayawardena, T.U.; Wu, J.; Fernando, I.P.S.; Jayawardena, T.U.; Wu, J. Marine Proteins and Peptides: Production, Biological Activities, and Potential Applications. Food Innov. Adv. 2023, 2, 69–84. [Google Scholar] [CrossRef]
- Gelse, K.; Pöschl, E.; Aigner, T. Collagens—Structure, Function, and Biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef] [PubMed]
- Aleman, A.; Martinez-Alvarez, O. Marine Collagen as a Source of Bioactive Molecules: A Review. Nat. Prod. J. 2013, 3, 105–114. [Google Scholar] [CrossRef]
- Arkhipkin, A.I.; Winter, A.G.; Nigmatullin, C.M. Heavy Fishery Exploitation Does Not Affect Sizes at Recruitment and Maturity of Squid Illex Argentinus and Doryteuthis Gahi, in the Southwest Atlantic. ICES J. Mar. Sci. 2022, 79, 182–192. [Google Scholar] [CrossRef]
- Nam, K.A.; You, S.G.; Kim, S.M. Molecular and Physical Characteristics of Squid (Todarodes Pacificus) Skin Collagens and Biological Properties of Their Enzymatic Hydrolysates. J. Food Sci. 2008, 73, C249–C255. [Google Scholar] [CrossRef]
- Tolano-Villaverde, I.J.; Ocaño-Higuera, V.; Ezquerra-Brauer, J.; Santos-Sauceda, I.; Santacruz-Ortega, H.; Cárdenas-López, J.L.; Rodríguez-Olibarria, G.; Márquez-Ríos, E. Physicochemical Characterization of Actomyosin–Paramyosin from Giant Squid Mantle (Dosidicus gigas). J. Sci. Food Agric. 2018, 98, 1787–1793. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Aluko, R.E. Chemometric Analysis of the Amino Acid Requirements of Antioxidant Food Protein Hydrolysates. Int. J. Mol. Sci. 2011, 12, 3148–3161. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Guillén, M.C.; Giménez, B.; López-Caballero, M.E.; Montero, M.P. Functional and Bioactive Properties of Collagen and Gelatin from Alternative Sources: A Review. Food Hydrocoll. 2011, 25, 1813–1827. [Google Scholar] [CrossRef]
- Akinyede, A.I.; Fagbemi, T.N.; Osundahunsi, O.F.; Aluko, R.E. Amino Acid Composition and Antioxidant Properties of the Enzymatic Hydrolysate of Calabash Nutmeg (Monodora myristica) and Its Membrane Ultrafiltration Peptide Fractions. J. Food Biochem. 2021, 45, e13437. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, J.A.; Blanco, M.; Massa, A.E.; Amado, I.R.; Pérez-Martín, R.I. Production of Fish Protein Hydrolysates from Scyliorhinus Canicula Discards with Antihypertensive and Antioxidant Activities by Enzymatic Hydrolysis and Mathematical Optimization Using Response Surface Methodology. Mar. Drugs 2017, 15, 306. [Google Scholar] [CrossRef] [PubMed]
- Segura Campos, M.R.; Peralta González, F.; Chel Guerrero, L.; Betancur Ancona, D. Angiotensin I-Converting Enzyme Inhibitory Peptides of Chia (Salvia Hispanica) Produced by Enzymatic Hydrolysis. Int. J. Food Sci. 2013, 2013, e158482. [Google Scholar] [CrossRef]
- Georgieva, D.N.; Stoeva, S.; Voelter, W.; Genov, N.; Betzel, C. Substrate Specificity of the Highly Alkalophilic Bacterial Proteinase Esperase: Relation to the X-Ray Structure. Curr. Microbiol. 2001, 42, 368–371. [Google Scholar] [CrossRef] [PubMed]
- García Arteaga, V.; Apéstegui Guardia, M.; Muranyi, I.; Eisner, P.; Schweiggert-Weisz, U. Effect of Enzymatic Hydrolysis on Molecular Weight Distribution, Techno-Functional Properties and Sensory Perception of Pea Protein Isolates. Innov. Food Sci. Emerg. Technol. 2020, 65, 102449. [Google Scholar] [CrossRef]
- Hedstrom, L. Trypsin: A Case Study in the Structural Determinants of Enzyme Specificity. Biol. Chem. 1996, 377, 465–470. [Google Scholar]
- Choonpicharn, S.; Jaturasitha, S.; Rakariyatham, N.; Suree, N.; Niamsup, H. Antioxidant and Antihypertensive Activity of Gelatin Hydrolysate from Nile Tilapia Skin. J. Food Sci. Technol. 2015, 52, 3134–3139. [Google Scholar] [CrossRef]
- Shavandi, A.; Hu, Z.; Teh, S.; Zhao, J.; Carne, A.; Bekhit, A.; Bekhit, A.E.-D.A. Antioxidant and Functional Properties of Protein Hydrolysates Obtained from Squid Pen Chitosan Extraction Effluent. Food Chem. 2017, 227, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Mendonça Diniz, F.; Martin, A.M. Influence of Process Variables on the Hydrolysis of Shark Muscle Protein/Influencia de Las Variables de Proceso En La Hidrólisis de Proteína Del Músculo de Tiburón. Food Sci. Technol. Int. 1998, 4, 91–98. [Google Scholar] [CrossRef]
- Lin, L.; Lv, S.; Li, B. Angiotensin-I-Converting Enzyme (ACE)-Inhibitory and Antihypertensive Properties of Squid Skin Gelatin Hydrolysates. Food Chem. 2012, 131, 225–230. [Google Scholar] [CrossRef]
- Zheng, Z.; Li, J.; Li, J.; Sun, H.; Liu, Y. Physicochemical and Antioxidative Characteristics of Black Bean Protein Hydrolysates Obtained from Different Enzymes. Food Hydrocoll. 2019, 97, 105222. [Google Scholar] [CrossRef]
- Alemán, A.; Pérez-Santín, E.; Bordenave-Juchereau, S.; Arnaudin, I.; Gómez-Guillén, M.C.; Montero, P. Squid Gelatin Hydrolysates with Antihypertensive, Anticancer and Antioxidant Activity. Food Res. Int. 2011, 44, 1044–1051. [Google Scholar] [CrossRef]
- Nuchprapha, A.; Paisansak, S.; Sangtanoo, P.; Srimongkol, P.; Saisavoey, T.; Reamtong, O.; Choowongkomon, K.; Karnchanatat, A. Two Novel ACE Inhibitory Peptides Isolated from Longan Seeds: Purification, Inhibitory Kinetics and Mechanisms. RSC Adv. 2020, 10, 12711–12720. [Google Scholar] [CrossRef]
- Dai, C.; Ma, H.; Luo, L.; Yin, X. Angiotensin I-Converting Enzyme (ACE) Inhibitory Peptide Derived from Tenebrio molitor (L.) Larva Protein Hydrolysate. Eur. Food Res. Technol. 2013, 236, 681–689. [Google Scholar] [CrossRef]
- Barbana, C.; Boye, J.I. Angiotensin I-Converting Enzyme Inhibitory Properties of Lentil Protein Hydrolysates: Determination of the Kinetics of Inhibition. Food Chem. 2011, 127, 94–101. [Google Scholar] [CrossRef]
- Wongtay, P.; Sangtanoo, P.; Sangvanich, P.; Karnchanatat, A. Variation in the Protein Composition and Biological Activity of King Cobra (Ophiophagus hannah) Venoms. Protein J. 2019, 38, 565–575. [Google Scholar] [CrossRef]
- Pujiastuti, D.Y.; Ghoyatul Amin, M.N.; Alamsjah, M.A.; Hsu, J.-L. Marine Organisms as Potential Sources of Bioactive Peptides That Inhibit the Activity of Angiotensin I-Converting Enzyme: A Review. Molecules 2019, 24, 2541. [Google Scholar] [CrossRef]
- Liu, Q.; Yao, Y.; Ibrahim, M.A.A.; Halawany, A.M.E.; Yang, L.; Zhang, X. Production of Dual Inhibitory Hydrolysate by Enzymatic Hydrolysis of Squid Processing By-Product. Mar. Biotechnol. 2022, 24, 293–302. [Google Scholar] [CrossRef] [PubMed]
- He, H.-L.; Chen, X.-L.; Wu, H.; Sun, C.-Y.; Zhang, Y.-Z.; Zhou, B.-C. High Throughput and Rapid Screening of Marine Protein Hydrolysates Enriched in Peptides with Angiotensin-I-Converting Enzyme Inhibitory Activity by Capillary Electrophoresis. Bioresour. Technol. 2007, 98, 3499–3505. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.-Y.; Kang, N.; Lee, J.-H.; Kim, J.-S.; Kim, W.-S.; Park, S.-J.; Kim, Y.-T.; Jeon, Y.-J. Angiotensin I-Converting Enzyme Inhibitory Peptides from an Enzymatic Hydrolysate of Flounder Fish (Paralichthys olivaceus) Muscle as a Potent Anti-Hypertensive Agent. Process Biochem. 2016, 51, 535–541. [Google Scholar] [CrossRef]
- Mosquera, M.; Giménez, B.; Ramos, S.; López-Caballero, M.E.; Gómez-Guillén, M.d.C.; Montero, P. Antioxidant, ACE-Inhibitory, and Antimicrobial Activities of Peptide Fractions Obtained From Dried Giant Squid Tunics. J. Aquat. Food Prod. Technol. 2016, 25, 444–455. [Google Scholar] [CrossRef]
- Nong, N.T.P.; Hsu, J.-L. Characteristics of Food Protein-Derived Antidiabetic Bioactive Peptides: A Literature Update. Int. J. Mol. Sci. 2021, 22, 9508. [Google Scholar] [CrossRef]
- Jin, R.; Teng, X.; Shang, J.; Wang, D.; Liu, N. Identification of Novel DPP–IV Inhibitory Peptides from Atlantic Salmon (Salmo salar) Skin. Food Res. Int. 2020, 133, 109161. [Google Scholar] [CrossRef]
- Taraszkiewicz, A.; Sinkiewicz, I.; Sommer, A.; Staroszczyk, H. The Biological Role of Prolyl Oligopeptidase and the Procognitive Potential of Its Peptidic Inhibitors from Food Proteins. Crit. Rev. Food Sci. Nutr. 2023, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.; Hayes, M.; Carney, B. Angiotensin-I-Converting Enzyme and Prolyl Endopeptidase Inhibitory Peptides from Natural Sources with a Focus on Marine Processing by-Products. Food Chem. 2011, 129, 235–244. [Google Scholar] [CrossRef]
- Sila, A.; Alvarez, O.M.; Haddar, A.; Frikha, F.; Dhulster, P.; Nedjar-Arroume, N.; Bougatef, A. Purification, Identification and Structural Modelling of DPP-IV Inhibiting Peptides from Barbel Protein Hydrolysate. J. Chromatogr. B 2016, 1008, 260–269. [Google Scholar] [CrossRef]
- Lajmi, K.; Gómez-Estaca, J.; Hammami, M.; Martínez-Alvarez, O. Upgrading Collagenous Smooth Hound By-Products: Effect of Hydrolysis Conditions, in Vitro Gastrointestinal Digestion and Encapsulation on Bioactive Properties. Food Biosci. 2019, 28, 99–108. [Google Scholar] [CrossRef]
- Martínez-Alvarez, O.; Batista, I.; Ramos, C.; Montero, P. Enhancement of ACE and Prolyl Oligopeptidase Inhibitory Potency of Protein Hydrolysates from Sardine and Tuna By-Products by Simulated Gastrointestinal Digestion. Food Funct. 2016, 7, 2066–2073. [Google Scholar] [CrossRef] [PubMed]
- Chalamaiah, M.; Dinesh Kumar, B.; Hemalatha, R.; Jyothirmayi, T. Fish Protein Hydrolysates: Proximate Composition, Amino Acid Composition, Antioxidant Activities and Applications: A Review. Food Chem. 2012, 135, 3020–3038. [Google Scholar] [CrossRef]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural Requirements of Angiotensin I-Converting Enzyme Inhibitory Peptides: Quantitative Structure-Activity Relationship Modeling of Peptides Containing 4–10 Amino Acid Residues. QSAR Comb. Sci. 2006, 25, 873–880. [Google Scholar] [CrossRef]
- Cushman, D.W.; Wang, F.L.; Fung, W.C.; Harvey, C.M.; DeForrest, J.M. Differentiation of Angiotensin–Converting Enzyme (ACE) Inhibitors by Their Selective Inhibition of ACE in Physiologically Important Target Organs. Am. J. Hypertens. 1989, 2, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Pihlanto-Leppälä, A. Bioactive Peptides Derived from Bovine Whey Proteins: Opioid and Ace-Inhibitory Peptides. Trends Food Sci. Technol. 2000, 11, 347–356. [Google Scholar] [CrossRef]
- Rho, S.J.; Lee, J.-S.; Chung, Y.I.; Kim, Y.-W.; Lee, H.G. Purification and Identification of an Angiotensin I-Converting Enzyme Inhibitory Peptide from Fermented Soybean Extract. Process Biochem. 2009, 44, 490–493. [Google Scholar] [CrossRef]
- Wang, R.; Lu, X.; Sun, Q.; Gao, J.; Ma, L.; Huang, J. Novel ACE Inhibitory Peptides Derived from Simulated Gastrointestinal Digestion in Vitro of Sesame (Sesamum Indicum L.) Protein and Molecular Docking Study. Int. J. Mol. Sci. 2020, 21, 1059. [Google Scholar] [CrossRef]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural Requirements of Angiotensin I-Converting Enzyme Inhibitory Peptides: Quantitative Structure−Activity Relationship Study of Di- and Tripeptides. J. Agric. Food Chem. 2006, 54, 732–738. [Google Scholar] [CrossRef]
- Chen, H.; Chen, Y.; Zheng, H.; Xiang, X.; Xu, L. A Novel Angiotensin-I-Converting Enzyme Inhibitory Peptide from Oyster: Simulated Gastro-Intestinal Digestion, Molecular Docking, Inhibition Kinetics and Antihypertensive Effects in Rats. Front. Nutr. 2022, 9, 981163. [Google Scholar] [CrossRef]
- Asoodeh, A.; Haghighi, L.; Chamani, J.; Ansari-Ogholbeyk, M.A.; Mojallal-Tabatabaei, Z.; Lagzian, M. Potential Angiotensin I Converting Enzyme Inhibitory Peptides from Gluten Hydrolysate: Biochemical Characterization and Molecular Docking Study. J. Cereal Sci. 2014, 60, 92–98. [Google Scholar] [CrossRef]
- Ling, Y.; Liping, S.; Yongliang, Z. Preparation and Identification of Novel Inhibitory Angiotensin-I-Converting Enzyme Peptides from Tilapia Skin Gelatin Hydrolysates: Inhibition Kinetics and Molecular Docking. Food Funct. 2018, 9, 5251–5259. [Google Scholar] [CrossRef] [PubMed]
- Olalere, O.A.; Yap, P.-G.; Gan, C.-Y. Comprehensive Review on Some Food-Derived Bioactive Peptides with Anti-Hypertension Therapeutic Potential for Angiotensin-Converting Enzyme (ACE) Inhibition. J. Proteins Proteom. 2023, 14, 129–161. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Virtual—A Novel Database of Food-Derived Peptides with In Silico-Predicted Biological Activity. Appl. Sci. 2022, 12, 7204. [Google Scholar] [CrossRef]
- Ding, Q.; Shults, N.V.; Gychka, S.G.; Harris, B.T.; Suzuki, Y.J. Protein Expression of Angiotensin-Converting Enzyme 2 (ACE2) Is Upregulated in Brains with Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 1687. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Wang, R.; Cheng, C.; Ma, Y.; Zhang, Y.; Lu, W. Preparation, Structural Properties, and in Vitro and in Vivo Activities of Peptides against Dipeptidyl Peptidase IV (DPP-IV) and α-Glucosidase: A General Review. Crit. Rev. Food Sci. Nutr. 2023, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lorey, S.; Stöckel-Maschek, A.; Faust, J.; Brandt, W.; Stiebitz, B.; Gorrell, M.D.; Kähne, T.; Mrestani-Klaus, C.; Wrenger, S.; Reinhold, D.; et al. Different Modes of Dipeptidyl Peptidase IV (CD26) Inhibition by Oligopeptides Derived from the N-Terminus of HIV-1 Tat Indicate at Least Two Inhibitor Binding Sites. Eur. J. Biochem. 2003, 270, 2147–2156. [Google Scholar] [CrossRef] [PubMed]
- Deacon, C.F. Physiology and Pharmacology of DPP-4 in Glucose Homeostasis and the Treatment of Type 2 Diabetes. Front. Endocrinol. 2019, 10, 440649. [Google Scholar]
- Liu, R.; Zheng, W.; Li, J.; Wang, L.; Wu, H.; Wang, X.; Shi, L. Rapid Identification of Bioactive Peptides with Antioxidant Activity from the Enzymatic Hydrolysate of Mactra Veneriformis by UHPLC–Q-TOF Mass Spectrometry. Food Chem. 2015, 167, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Chanajon, P.; Tian, F.; Noisa, P.; Roytrakul, S.; Yongsawatdigul, J. Corn Gluten Meal Peptides Inhibit Prolyl Oligopeptidase and Modulate α-Synuclein Aggregation in KCl-Treated SH-SY5Y Cells. J. Funct. Foods 2023, 104, 105501. [Google Scholar] [CrossRef]
- Fülöp, V.; Böcskei, Z.; Polgár, L. Prolyl Oligopeptidase: An Unusual β-Propeller Domain Regulates Proteolysis. Cell 1998, 94, 161–170. [Google Scholar] [CrossRef]
- Pripp, A.H. Quantitative Structure−Activity Relationship of Prolyl Oligopeptidase Inhibitory Peptides Derived from β-Casein Using Simple Amino Acid Descriptors. J. Agric. Food Chem. 2006, 54, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Thiex, N.; Novotny, L.; Crawford, A. Determination of Ash in Animal Feed: AOAC Official Method 942.05 Revisited. J. AOAC Int. 2012, 95, 1392–1397. [Google Scholar] [CrossRef] [PubMed]
- Ebeling, M.E. The Dumas Method for Nitrogen in Feeds. J. Assoc. Off. Anal. Chem. 1968, 51, 766–770. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bougatef, H.; de la Vega-Fernández, C.; Sila, A.; Bougatef, A.; Martínez-Alvarez, O. Identification of ACE I-Inhibitory Peptides Released by the Hydrolysis of Tub Gurnard (Chelidonichthys lucerna) Skin Proteins and the Impact of Their In Silico Gastrointestinal Digestion. Mar. Drugs 2023, 21, 131. [Google Scholar] [CrossRef] [PubMed]
- Adler-Nissen, J. Enzymic Hydrolysis of Food Proteins; Elsevier Applied Science Publishers: Amsterdam, The Netherlands, 1986; ISBN 0-85334-386-1. [Google Scholar]
- Martínez-Alvarez, O.; Ravallec, R.; Cudennec, B.; Guimas, L.; Delannoy, C.; Fouchereau-Peron, M. Effect of Daily Gavage with a Collagen Hydrolysate Containing Calcitonin Gene-Related Peptide (CGRP)-like Molecules on Plasma CGRP-Levels in Rats. J. Funct. Foods 2012, 4, 767–775. [Google Scholar] [CrossRef]
- Djellouli, M.; López-Caballero, M.E.; Arancibia, M.Y.; Karam, N.; Martínez-Alvarez, O. Antioxidant and Antimicrobial Enhancement by Reaction of Protein Hydrolysates Derived from Shrimp By-Products with Glucosamine. Waste Biomass Valorization 2020, 11, 2491–2505. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Types | Amino Acids | ‰ |
|---|---|---|
| Essential amino acid (EAA) | Leu (L) | 55 |
| Arg (R) | 53 | |
| Thr (T) | 42 | |
| Lys (K) | 42 | |
| Val (V) | 37 | |
| Ile (I) | 33 | |
| Phe (F) | 28 | |
| Met (M) | 22 | |
| His (H) | 14 | |
| Non-essential amino acids (NEAA) | Gly (G) | 215 |
| Glx (E + Q) | 98 | |
| Asx (D + N) | 87 | |
| Ala (A) | 84 | |
| Ser (S) | 55 | |
| Pro (P) | 42 | |
| Tyr (Y) | 18 | |
| Cys (C) | 6 | |
| Hyp | 58 | |
| Hyl | 12 | |
| THAA * | 516 | |
| TEAA * | 325 | |
| Pro + Hyp | 100 |
| Enzyme | MW (Da) | ACE Inhibition (IC50) | PEP Inhibition (IC50) | DPP-IV Inhibition (IC50) | ABTS (mg Ascorbic acid/g) | FRAP (μM FeSO4/mg) |
|---|---|---|---|---|---|---|
| Esperase | 720 | 53 ± 6 a | 38 ± 3 a | 673 ± 5 a | 16.61 ± 0.03 a | 41.62 ± 0.56 a |
| Alcalase | 1100 | 37 ± 3 b | 163 ± 21b | 702 ± 3 b | 10.76 ± 0.03 b | 47.41 ± 2.77 b |
| Trypsin | 2500 | 74 ± 11 c | 129 ± 15 b | 863 ± 9 c | 18.68 ± 0.01 c | 41.95 ±1.30 a |
| Enzyme Used | Fraction | ACE (25 µg/mL) | DPP-IV (800 µg/mL) | PEP (400 µg/mL) |
|---|---|---|---|---|
| Esperase | 2 | 84 ± 3% | ||
| Esperase | 9 | 66 ± 4% | 86 ± 9% | 55 ± 0% |
| Alcalase | 8 | 52 ± 8% | 59 ± 5% |
| Enzyme Used | Fraction and Subfraction | ACE | DPP-IV | PEP |
|---|---|---|---|---|
| Esperase | 2 (3) | NA | 97 ± 6 | NA |
| Esperase | 9 (1) | 16 ± 2 | NA | NA |
| Esperase | 9 (6) | NA | NA | 55 ± 4 |
| Alcalase | 8 (1) | 66 ± 10 | NA | NA |
| Peptide | −10lgP | ALC Score | Mass (Da) | Error (ppm) | Abundance (%) |
|---|---|---|---|---|---|
| EGRGLP(+15.99)GL | 87 | 813.433 | −0.3 | 12.6 | |
| LGRGEP(+15.99)GL | 84 | 813.433 | −0.3 | 12.6 | |
| GEAGPP(+15.99)GLVGLP(+15.99)GER | 96 | 1436.724 | 0.3 | 6.1 | |
| AGDDAPRAVFPS | 77 | 1201.572 | 0.7 | 3.6 | |
| GTLGPDGPRGPSGL | 94 | 1279.652 | 0.6 | 3.5 | |
| KIKIIAPPERKY | 72 | 1454.897 | 0.7 | 2.5 | |
| TGPRTP(+15.99)GLAGPE | 84 | 1167.587 | 0.4 | 2.2 | |
| AEREIVRDIKEKL | 65 | 1597.915 | −1.4 | 2.1 | |
| GATGP(+15.99)P(+15.99)GPIGHL | 70 | 1104.554 | 1.3 | 2.1 | |
| EIK(+14.02)DIVTDVGM(+15.99)N | 72 | 1362.670 | 2 | 1.7 | |
| NLLLEPH | 93 | 834.459 | −0.2 | 1.7 | |
| QGEQGIL(subA)GIP(+15.99)GPM(sub V)GPPGP(+15.99)KGH | 53 | 2055.018 | −7.5 | 1.6 | |
| NWDDM(+15.99)EKIWHH | 82 | 1525.640 | −0.7 | 1.6 | |
| LDVKLPN | 95 | 797.464 | −0.3 | 1.0 | |
| DIRPEVPEMVNQ | 74 | 1425.692 | 1.4 | 1.0 | |
| LARKGVL | 87 | 755.501 | 2.7 | 0.9 | |
| RAKLGVL | 85 | 755.501 | 2.7 | 0.9 | |
| RVAPEEHPVL | 77 | 1145.619 | 0.3 | 0.9 | |
| MYPGIADRM(+15.99) | 61 | 1068.473 | 1.2 | 0.8 | |
| GDDAPRAVFPS | 74 | 1130.535 | 1.1 | 0.8 |
| Peptide | −10lgP | ALC Score | Mass (Da) | Error (ppm) | Abundance (%) |
|---|---|---|---|---|---|
| SHPDLRLA | 93 | 907.487 | 0 | 8.59 | |
| LGMP(+15.99)GRL | 92 | 758.410 | −0.4 | 8.48 | |
| GNK(+15.99)GPVGPL | 88 | 853.464 | 1.6 | 8.29 | |
| KYPIEH(+14.02)GIVT | 52.83 | 1169.644 | 0.8 | 7.40 | |
| NGK(+15.99)GPVGPL | 90 | 853.464 | 1.6 | 4.15 | |
| HN(+.98)KIFIN | 50.35 | 885.470 | 0.5 | 2.42 | |
| GERGPAGPVGVP(+15.99)GESGPP(+15.99)GPIGSR | 84.89 | 2215.096 | 2.2 | 2.33 | |
| AVDGVP(+15.99)GNDGPVGSV | 80 | 1354.635 | 2.1 | 1.89 | |
| LLLEPH | 95 | 720.41 | 0.3 | 1.73 | |
| GTP(+15.99)GELGLP(+15.99)GELQK(+15.99) | 83 | 1442.722 | 3 | 1.47 | |
| KVKDDWELR | 90 | 1187.629 | 0.3 | 1.14 | |
| GHP(+15.99)GKDGLPT(sub G)FPGERGETG | 43.58 | 1923.906 | 1.6 | 0.99 | |
| WDDM(+15.99)EKIWHH | 56.47 | 1411.597 | −0.1 | 0.81 | |
| LGPDGPRGPSGL | 86 | 1121.582 | 1 | 0.81 | |
| LGPDGPRGAP(+15.99)GL | 84 | 1121.582 | 1.6 | 0.81 | |
| KWDKVEKL | 97 | 1044.596 | 1.6 | 0.80 | |
| TERGYSF | 47.93 | 858.387 | −0.4 | 0.79 | |
| M(+15.99)VPFPR | 90 | 761.389 | −0.4 | 0.77 | |
| KVGDLLR | 98 | 799.491 | −0.9 | 0.75 | |
| APRAVFPS | 91 | 843.460 | 0.5 | 0.72 |
| Peptide | −10lgP | Mass (Da) | Error (ppm) | Abundance (%) |
|---|---|---|---|---|
| GEP(+15.99)GEQGPP(+15.99)GSP(+15.99)GPR | 60.08 | 1465.641 | 6.7 | 21.62 |
| K(+15.99)GPDGEHGRDGNSGSR | 54.03 | 1640.724 | 1.9 | 16.85 |
| GAP(+15.99)GPSGP(+15.99)P(+15.99)G | 34.45 | 840.358 | 5.5 | 11.02 |
| GNTGPAGP(+15.99)P(+15.99)GPQ | 48.02 | 1080.482 | 6.3 | 10.62 |
| P(+57.02)SGPP(+15.99)GQPGN | 45.81 | 979.435 | 4.7 | 7.48 |
| KGPDGEHGRDGNSGSR | 47.62 | 1624.730 | 4.2 | 6.13 |
| GADGERGPRGNRGDTGAS | 40.86 | 1728.789 | 4.3 | 4.74 |
| GPAGP(+15.99)P(+15.99)GAPG | 42.73 | 808.369 | 4.7 | 4.43 |
| GPTGP(+15.99)AGPAG | 40.53 | 796.370 | 4.1 | 2.97 |
| RHPEYAVSVLLR | 52.09 | 1438.804 | 3.2 | 1.63 |
| GAP(+15.99)GFP(+15.99)GPR | 34.34 | 886.427 | 5.8 | 1.51 |
| GPDGEHGRDGNSGSR | 65.38 | 1496.635 | 2.4 | 1.51 |
| IIAPPERKY | 31.82 | 1085.623 | 3.8 | 1.30 |
| GPP(+15.99)GN(+.98)PGP(+15.99)K | 31.21 | 852.395 | 6.3 | 1.25 |
| K(+15.99)GPDGEHGRDGD(sub N)SGSR | 55.43 | 1641.708 | 8.7 | 0.99 |
| GEHGRDGNSGSR | 39.96 | 1227.534 | 2.1 | 0.94 |
| SLLEGQDAK | 33.12 | 959.492 | 4.2 | 0.75 |
| GPDGDRGAP(+15.99)GPQ(+.98)G | 37.07 | 1196.505 | 2.0 | 0.74 |
| Peptide | −10lgP | ALC Score | Mass (Da) | Error (ppm) | Abundance (%) |
|---|---|---|---|---|---|
| SHPDLRLA | 93 | 907.487 | 0 | 8.59 | |
| LGMP(+15.99)GRL | 92 | 758.410 | −0.4 | 8.48 | |
| GNK(+15.99)GPVGPL | 88 | 853.464 | 1.6 | 8.29 | |
| KYPIEH(+14.02)GIVT | 52.83 | 1169.644 | 0.8 | 7.40 | |
| NGK(+15.99)GPVGPL | 90 | 853.464 | 1.6 | 4.15 | |
| HN(+.98)KIFIN | 50.35 | 885.470 | 0.5 | 2.42 | |
| GERGPAGPVGVP(+15.99)GESGPP(+15.99)GPIGSR | 84.89 | 2215.096 | 2.2 | 2.33 | |
| AVDGVP(+15.99)GNDGPVGSV | 80 | 1354.635 | 2.1 | 1.89 | |
| LLLEPH | 95 | 720.41 | 0.3 | 1.73 | |
| GTP(+15.99)GELGLP(+15.99)GELQK(+15.99) | 83 | 1442.722 | 3 | 1.47 | |
| KVKDDWELR | 90 | 1187.629 | 0.3 | 1.14 | |
| GHP(+15.99)GKDGLPT(sub G)FPGERGETG | 43.58 | 1923.906 | 1.6 | 0.99 | |
| WDDM(+15.99)EKIWHH | 56.47 | 1411.597 | −0.1 | 0.81 | |
| LGPDGPRGPSGL | 86 | 1121.582 | 1 | 0.81 | |
| LGPDGPRGAP(+15.99)GL | 84 | 1121.582 | 1.6 | 0.81 | |
| KWDKVEKL | 97 | 1044.596 | 1.6 | 0.80 | |
| TERGYSF | 47.93 | 858.387 | −0.4 | 0.79 | |
| M(+15.99)VPFPR | 90 | 761.389 | −0.4 | 0.77 | |
| KVGDLLR | 98 | 799.491 | −0.9 | 0.75 | |
| APRAVFPS | 91 | 843.460 | 0.5 | 0.72 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bougatef, H.; Sila, A.; Bougatef, A.; Martínez-Alvarez, O. Protein Hydrolysis as a Way to Valorise Squid-Processing Byproducts: Obtaining and Identification of ACE, DPP-IV and PEP Inhibitory Peptides. Mar. Drugs 2024, 22, 156. https://doi.org/10.3390/md22040156
Bougatef H, Sila A, Bougatef A, Martínez-Alvarez O. Protein Hydrolysis as a Way to Valorise Squid-Processing Byproducts: Obtaining and Identification of ACE, DPP-IV and PEP Inhibitory Peptides. Marine Drugs. 2024; 22(4):156. https://doi.org/10.3390/md22040156
Chicago/Turabian StyleBougatef, Hajer, Assaad Sila, Ali Bougatef, and Oscar Martínez-Alvarez. 2024. "Protein Hydrolysis as a Way to Valorise Squid-Processing Byproducts: Obtaining and Identification of ACE, DPP-IV and PEP Inhibitory Peptides" Marine Drugs 22, no. 4: 156. https://doi.org/10.3390/md22040156
APA StyleBougatef, H., Sila, A., Bougatef, A., & Martínez-Alvarez, O. (2024). Protein Hydrolysis as a Way to Valorise Squid-Processing Byproducts: Obtaining and Identification of ACE, DPP-IV and PEP Inhibitory Peptides. Marine Drugs, 22(4), 156. https://doi.org/10.3390/md22040156