
Porphyran Attenuates Neuronal Loss in the Hippocampal CA1 Subregion Induced by Ischemia and Reperfusion in Gerbils by Inhibiting NLRP3 Inflammasome-Mediated Neuroinflammation
 ,
,  , and
, and 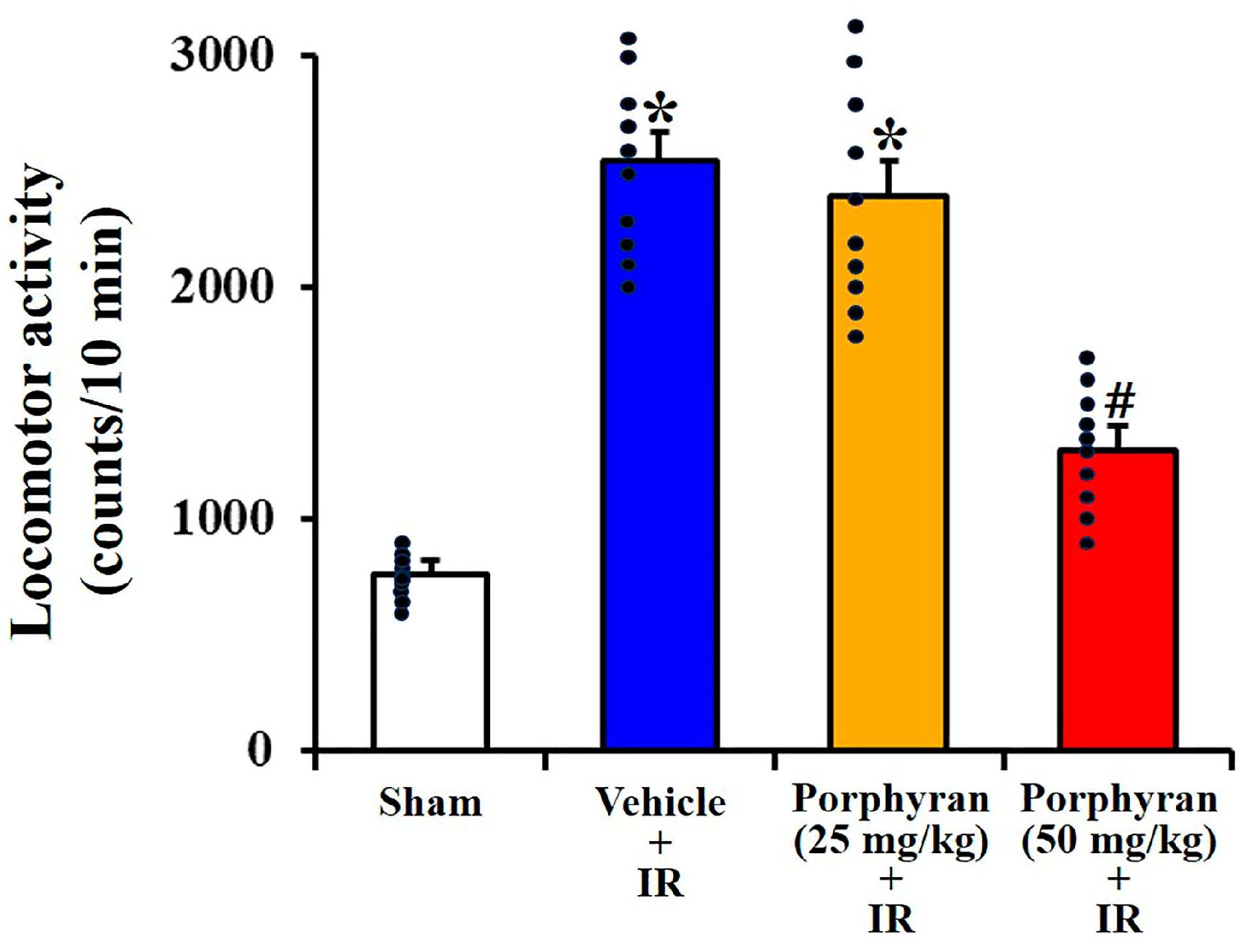{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Alleviation of IR-Induced Locomotor Hyperactivity by Porphyran
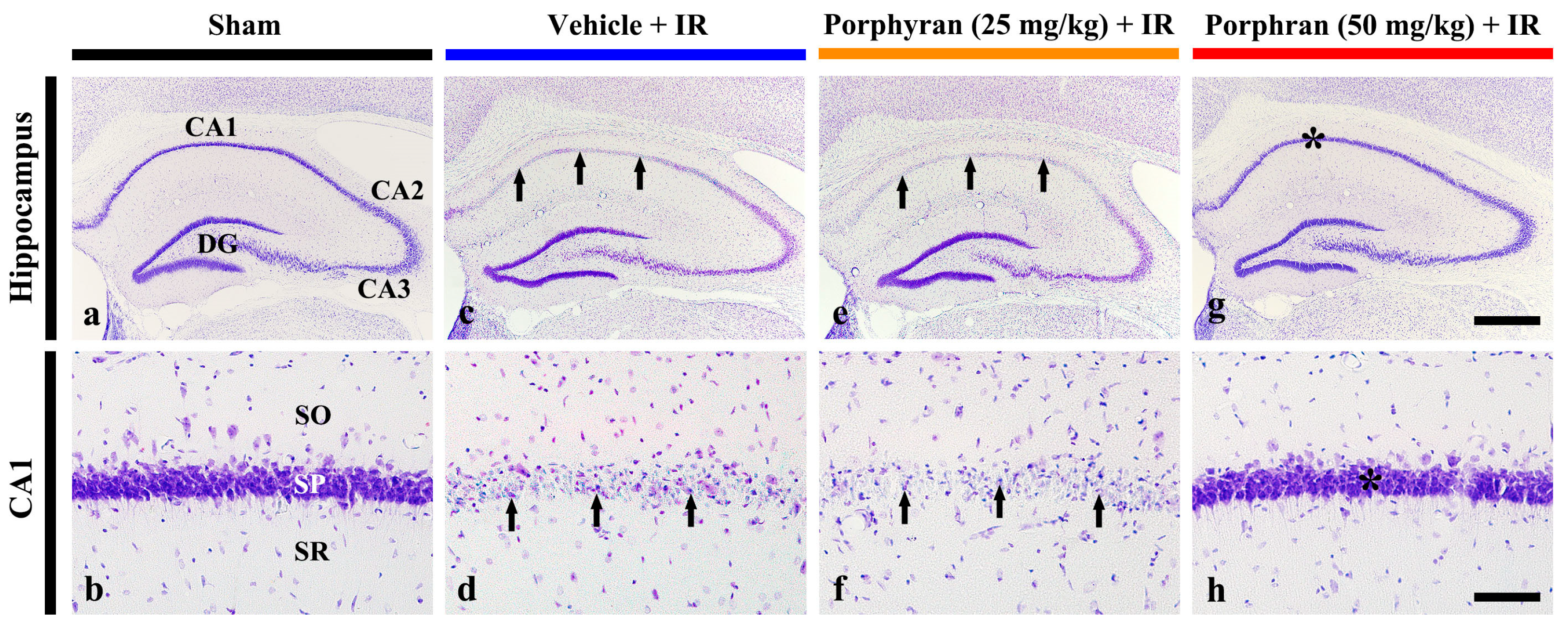
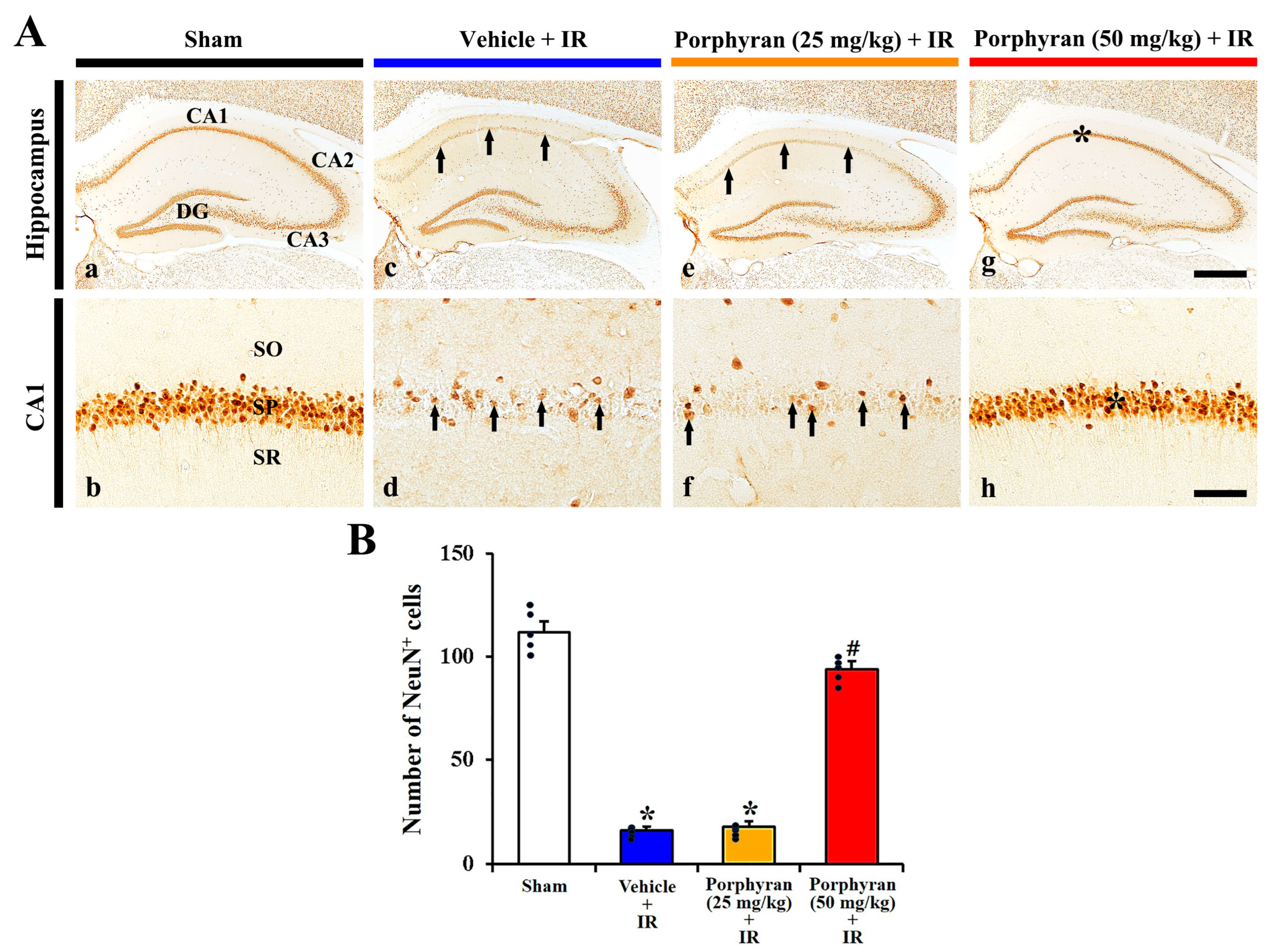
2.2. Reduction in IR-Induced Pyramidal Cell Death by Porphyran
2.2.1. Cresyl Violet (CV)-Stained Cells
2.2.2. Neuronal Nuclei (NeuN)+ Cells
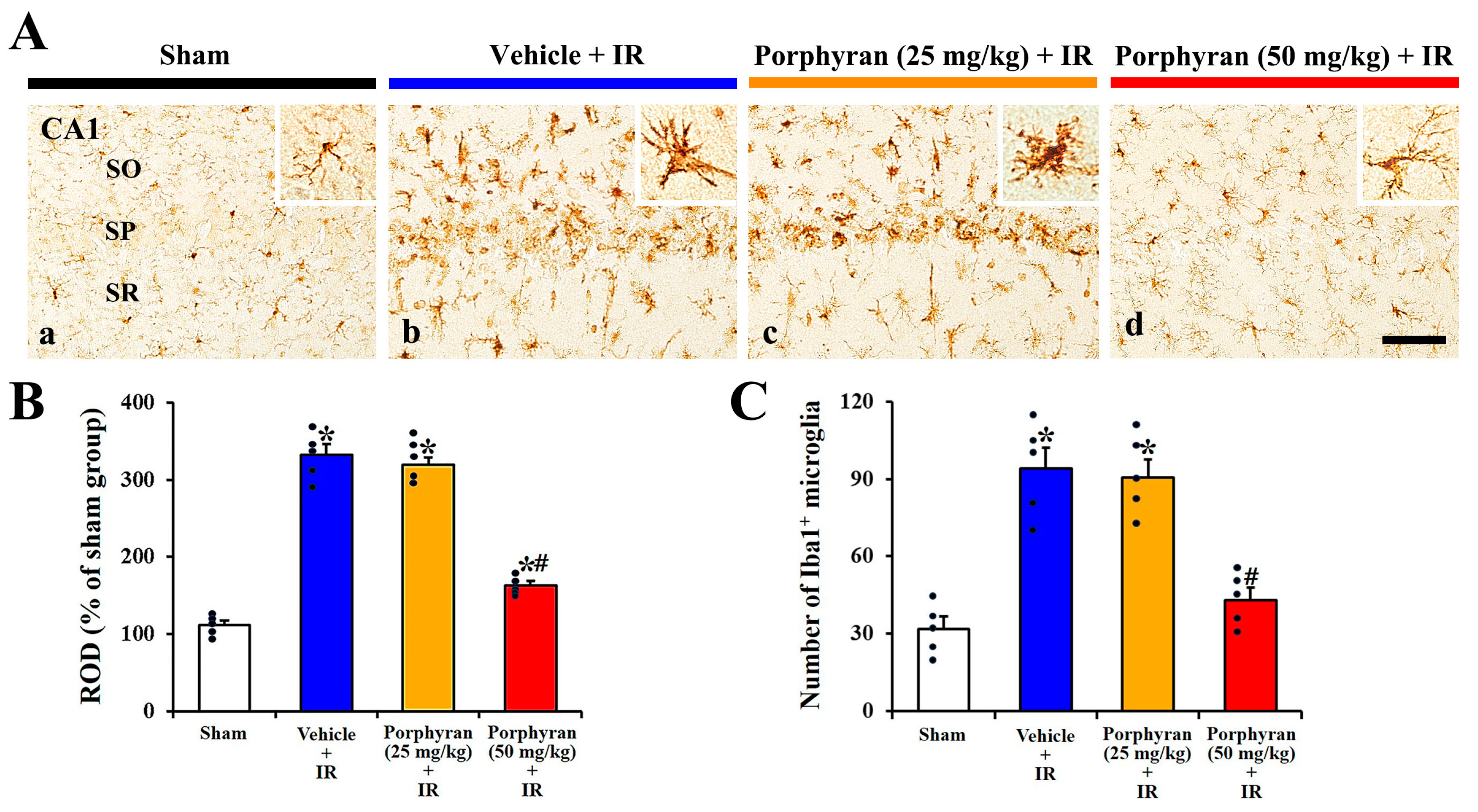
2.3. Inhibition of IR-Induced Microgliosis by Porphyran
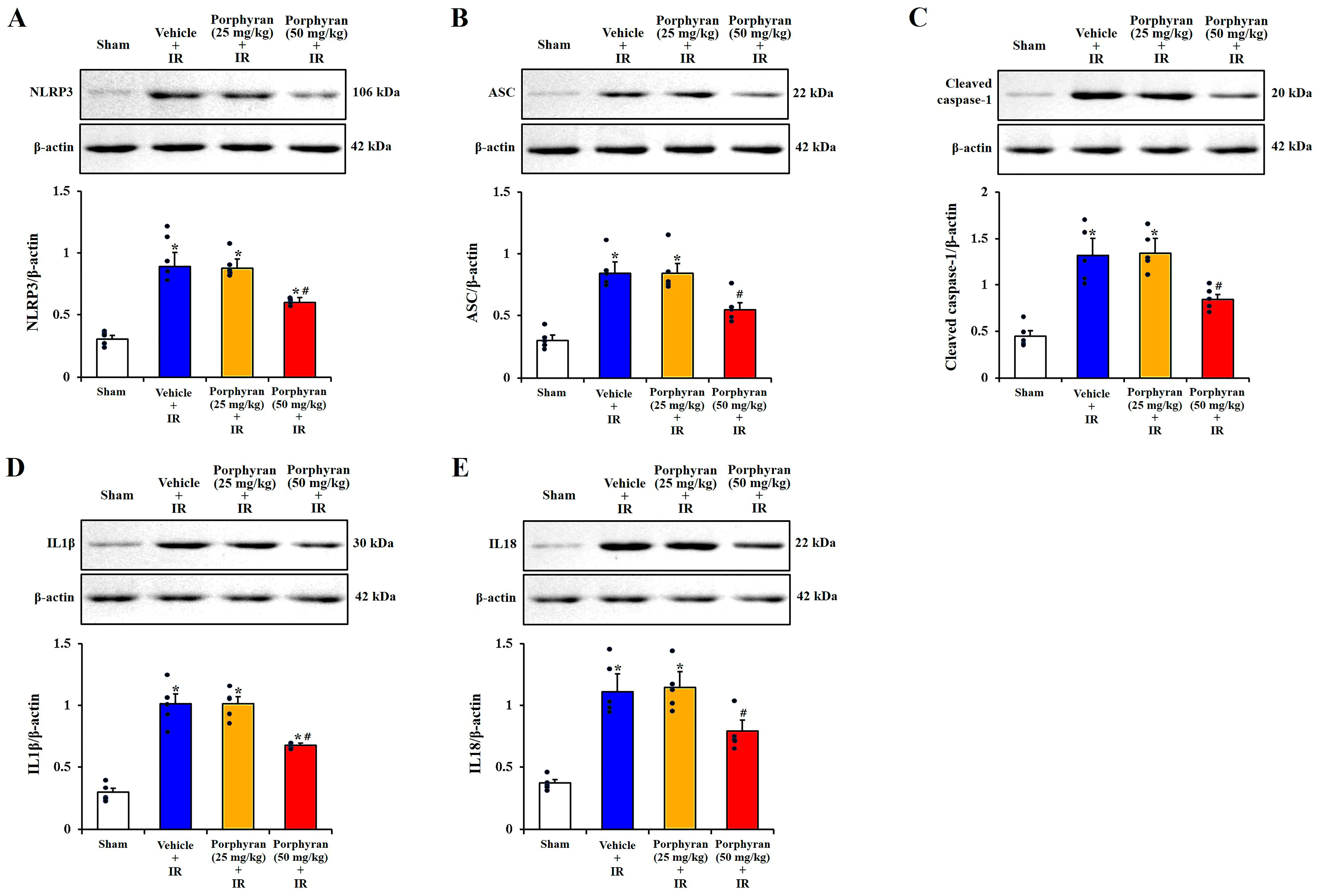
2.4. Attenuation of IR-Induced NLRP3 Inflammasome-Mediated Neuroinflammation by Porphyran
3. Discussion
4. Materials and Methods
4.1. Ethical Statement and Experimental Animals
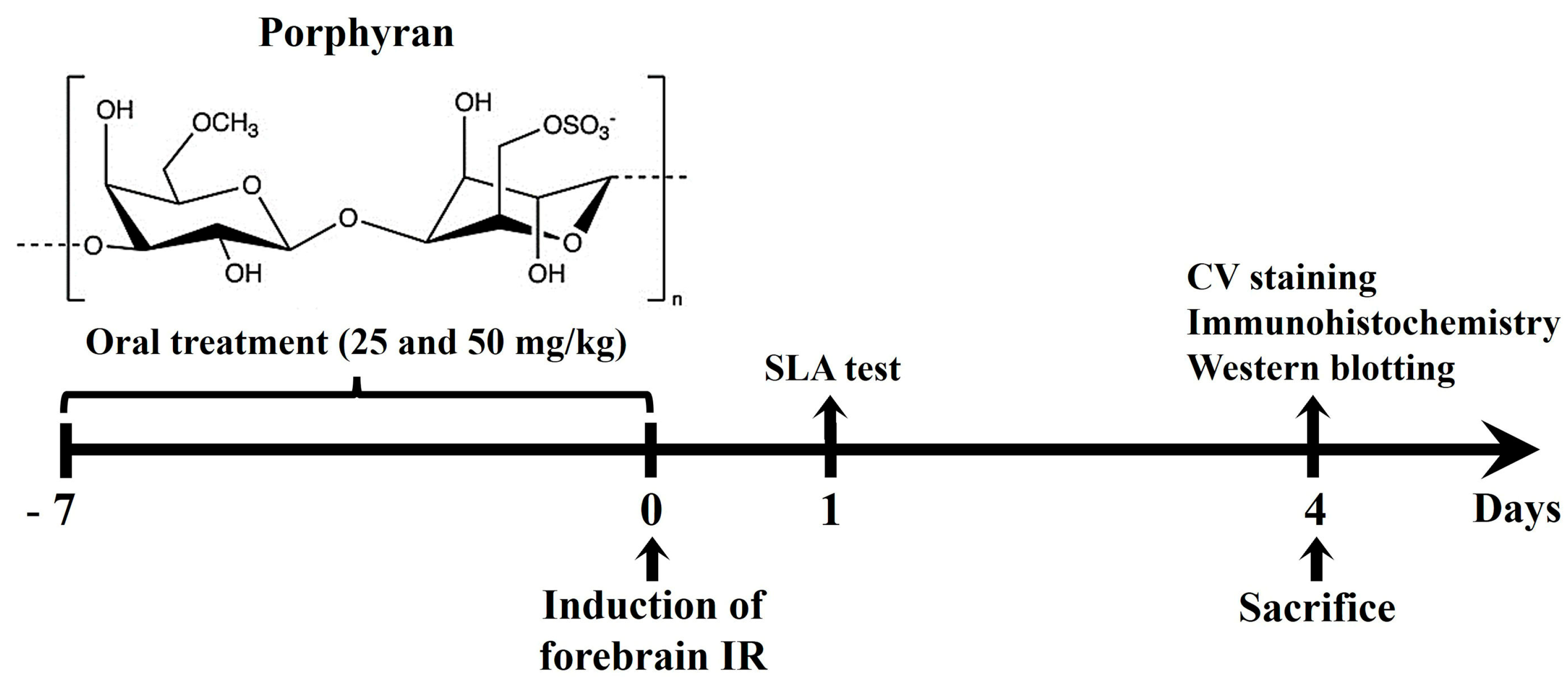
4.2. Administration of Porphyran
4.3. Induction of IR Injury
4.4. SLA Test
4.5. Tissue Preparation for Microscopic Observations
4.6. CV Staining
4.7. Immunohistochemistry
4.8. Western Blotting
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ding, X.D.; Zheng, N.N.; Cao, Y.Y.; Zhao, G.Y.; Zhao, P. Dexmedetomidine preconditioning attenuates global cerebral ischemic injury following asphyxial cardiac arrest. Int. J. Neurosci. 2016, 126, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Salazar, J.D.; Wityk, R.J.; Grega, M.A.; Borowicz, L.M.; Doty, J.R.; Petrofski, J.A.; Baumgartner, W.A. Stroke after cardiac surgery: Short- and long-term outcomes. Ann. Thorac. Surg. 2001, 72, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Sifat, A.E.; Nozohouri, S.; Archie, S.R.; Chowdhury, E.A.; Abbruscato, T.J. Brain Energy Metabolism in Ischemic Stroke: Effects of Smoking and Diabetes. Int. J. Mol. Sci. 2022, 23, 8512. [Google Scholar] [CrossRef] [PubMed]
- Globus, M.Y.; Busto, R.; Martinez, E.; Valdes, I.; Dietrich, W.D.; Ginsberg, M.D. Comparative effect of transient global ischemia on extracellular levels of glutamate, glycine, and gamma-aminobutyric acid in vulnerable and nonvulnerable brain regions in the rat. J. Neurochem. 1991, 57, 470–478. [Google Scholar] [CrossRef]
- Lin, C.S.; Polsky, K.; Nadler, J.V.; Crain, B.J. Selective neocortical and thalamic cell death in the gerbil after transient ischemia. Neuroscience 1990, 35, 289–299. [Google Scholar] [CrossRef]
- Wahul, A.B.; Joshi, P.C.; Kumar, A.; Chakravarty, S. Transient global cerebral ischemia differentially affects cortex, striatum and hippocampus in Bilateral Common Carotid Arterial occlusion (BCCAo) mouse model. J. Chem. Neuroanat. 2018, 92, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Park, J.H.; Ahn, J.H.; Kim, I.H.; Cho, J.H.; Choi, J.H.; Yoo, K.Y.; Lee, C.H.; Hwang, I.K.; Cho, J.H.; et al. New GABAergic Neurogenesis in the Hippocampal CA1 Region of a Gerbil Model of Long-Term Survival after Transient Cerebral Ischemic Injury. Brain Pathol. 2016, 26, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Sato-Maeda, M.; Fujimura, M.; Rashad, S.; Morita-Fujimura, Y.; Niizuma, K.; Sakata, H.; Ikawa, S.; Tominaga, T. Transient Global Cerebral Ischemia Induces RNF213, a Moyamoya Disease Susceptibility Gene, in Vulnerable Neurons of the Rat Hippocampus CA1 Subregion and Ischemic Cortex. J. Stroke Cerebrovasc. Dis. 2017, 26, 1904–1911. [Google Scholar] [CrossRef]
- Chi, X.X.; Xu, Z.C. Differential changes of potassium currents in CA1 pyramidal neurons after transient forebrain ischemia. J. Neurophysiol. 2000, 84, 2834–2843. [Google Scholar] [CrossRef]
- Niizuma, K.; Endo, H.; Nito, C.; Myer, D.J.; Kim, G.S.; Chan, P.H. The PIDDosome mediates delayed death of hippocampal CA1 neurons after transient global cerebral ischemia in rats. Proc. Natl. Acad. Sci. USA 2008, 105, 16368–16373. [Google Scholar] [CrossRef]
- Mohd Sairazi, N.S.; Sirajudeen, K.N.S. Natural Products and Their Bioactive Compounds: Neuroprotective Potentials against Neurodegenerative Diseases. Evid. Based Complement. Alternat Med. 2020, 2020, 6565396. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.F.; Zhang, Z.; Wang, F.; Chen, J.G. Natural compounds from traditional medicinal herbs in the treatment of cerebral ischemia/reperfusion injury. Acta Pharmacol. Sin. 2010, 31, 1523–1531. [Google Scholar] [CrossRef] [PubMed]
- Che, N.; Ma, Y.; Xin, Y. Protective Role of Fucoidan in Cerebral Ischemia-Reperfusion Injury through Inhibition of MAPK Signaling Pathway. Biomol. Ther. 2017, 25, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Ahn, J.H.; Song, M.; Kim, D.W.; Lee, T.K.; Lee, J.C.; Kim, Y.M.; Kim, J.D.; Cho, J.H.; Hwang, I.K.; et al. Pretreated fucoidan confers neuroprotection against transient global cerebral ischemic injury in the gerbil hippocampal CA1 area via reducing of glial cell activation and oxidative stress. Biomed. Pharmacother. 2019, 109, 1718–1727. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.K.; Ahn, J.H.; Park, C.W.; Kim, B.; Park, Y.E.; Lee, J.C.; Park, J.H.; Yang, G.E.; Shin, M.C.; Cho, J.H.; et al. Pre-Treatment with Laminarin Protects Hippocampal CA1 Pyramidal Neurons and Attenuates Reactive Gliosis Following Transient Forebrain Ischemia in Gerbils. Mar. Drugs 2020, 18, 52. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Ahn, J.H.; Lee, T.K.; Park, C.W.; Kim, B.; Lee, J.C.; Kim, D.W.; Shin, M.C.; Cho, J.H.; Lee, C.H.; et al. Laminarin Pretreatment Provides Neuroprotection against Forebrain Ischemia/Reperfusion Injury by Reducing Oxidative Stress and Neuroinflammation in Aged Gerbils. Mar. Drugs 2020, 18, 213. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, Q.; Wang, J.; Zhang, H.; Niu, X.; Li, P. Preparation of the different derivatives of the low-molecular-weight porphyran from Porphyra haitanensis and their antioxidant activities in vitro. Int. J. Biol. Macromol. 2009, 45, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Isaka, S.; Cho, K.; Nakazono, S.; Abu, R.; Ueno, M.; Kim, D.; Oda, T. Antioxidant and anti-inflammatory activities of porphyran isolated from discolored nori (Porphyra yezoensis). Int. J. Biol. Macromol. 2015, 74, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, X.; Pan, Y.; Wang, G.; Mao, G. The degraded polysaccharide from Pyropia haitanensis represses amyloid beta peptide-induced neurotoxicity and memory in vivo. Int. J. Biol. Macromol. 2020, 146, 725–729. [Google Scholar] [CrossRef]
- Liu, Y.; Geng, L.; Zhang, J.; Wang, J.; Zhang, Q.; Duan, D.; Zhang, Q. Oligo-Porphyran Ameliorates Neurobehavioral Deficits in Parkinsonian Mice by Regulating the PI3K/Akt/Bcl-2 Pathway. Mar. Drugs 2018, 16, 82. [Google Scholar] [CrossRef]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [PubMed]
- Clifton, G.L.; Taft, W.C.; Blair, R.E.; Choi, S.C.; DeLorenzo, R.J. Conditions for pharmacologic evaluation in the gerbil model of forebrain ischemia. Stroke 1989, 20, 1545–1552. [Google Scholar] [CrossRef]
- Islam, M.S.; Shin, H.Y.; Yoo, Y.J.; Kim, R.; Jang, Y.J.; Akanda, M.R.; Tae, H.J.; Kim, I.S.; Ahn, D.; Park, B.Y. Olanzapine Ameliorates Ischemic Stroke-like Pathology in Gerbils and H2O2-Induced Neurotoxicity in SH-SY5Y Cells via Inhibiting the MAPK Signaling Pathway. Antioxidants 2022, 11, 1697. [Google Scholar] [CrossRef] [PubMed]
- Hannan, M.A.; Dash, R.; Haque, M.N.; Mohibbullah, M.; Sohag, A.A.M.; Rahman, M.A.; Uddin, M.J.; Alam, M.; Moon, I.S. Neuroprotective Potentials of Marine Algae and Their Bioactive Metabolites: Pharmacological Insights and Therapeutic Advances. Mar. Drugs 2020, 18, 347. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Yuan, Y.; Zheng, Y.; Sheng, R.; Liu, L.; Xie, F.; Tan, J. Anti-cerebral ischemia reperfusion injury of polysaccharides: A review of the mechanisms. Biomed. Pharmacother. 2021, 137, 111303. [Google Scholar] [CrossRef] [PubMed]
- Janac, B.; Radenovic, L.; Selakovic, V.; Prolic, Z. Time course of motor behavior changes in Mongolian gerbils submitted to different durations of cerebral ischemia. Behav. Brain Res. 2006, 175, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.B.; Zimmer, J.; Sams-Dodd, F. Postischemic hyperactivity in the Mongolian gerbil correlates with loss of hippocampal neurons. Behav. Neurosci. 1997, 111, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Katsuta, K.; Umemura, K.; Ueyama, N.; Matsuoka, N. Pharmacological evidence for a correlation between hippocampal CA1 cell damage and hyperlocomotion following global cerebral ischemia in gerbils. Eur. J. Pharmacol. 2003, 467, 103–109. [Google Scholar] [CrossRef]
- Li, X.J.; Ye, Q.F. Fucoidan reduces inflammatory response in a rat model of hepatic ischemia-reperfusion injury. Can. J. Physiol. Pharmacol. 2015, 93, 999–1005. [Google Scholar] [CrossRef]
- Muzio, L.; Viotti, A.; Martino, G. Microglia in Neuroinflammation and Neurodegeneration: From Understanding to Therapy. Front. Neurosci. 2021, 15, 742065. [Google Scholar] [CrossRef]
- Streit, W.J.; Mrak, R.E.; Griffin, W.S. Microglia and neuroinflammation: A pathological perspective. J. Neuroinflamm. 2004, 1, 14. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.K.; Kim, H.; Song, M.; Lee, J.C.; Park, J.H.; Ahn, J.H.; Yang, G.E.; Kim, H.; Ohk, T.G.; Shin, M.C.; et al. Time-course pattern of neuronal loss and gliosis in gerbil hippocampi following mild, severe, or lethal transient global cerebral ischemia. Neural Regen. Res. 2019, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, J.R.; Koerner, I.P.; Moller, T. Microglia in ischemic brain injury. Future Neurol. 2010, 5, 227–246. [Google Scholar] [CrossRef] [PubMed]
- Uno, H.; Matsuyama, T.; Akita, H.; Nishimura, H.; Sugita, M. Induction of tumor necrosis factor-alpha in the mouse hippocampus following transient forebrain ischemia. J. Cereb. Blood Flow. Metab. 1997, 17, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Su, X.; Li, S.; Zhao, F.; Mu, D.; Qu, Y. Microglia and Their Promising Role in Ischemic Brain Injuries: An Update. Front. Cell Neurosci. 2020, 14, 211. [Google Scholar] [CrossRef] [PubMed]
- Gaire, B.P. Microglia as the Critical Regulators of Neuroprotection and Functional Recovery in Cerebral Ischemia. Cell. Mol. Neurobiol. 2022, 42, 2505–2525. [Google Scholar] [CrossRef]
- Yrjanheikki, J.; Keinanen, R.; Pellikka, M.; Hokfelt, T.; Koistinaho, J. Tetracyclines inhibit microglial activation and are neuroprotective in global brain ischemia. Proc. Natl. Acad. Sci. USA 1998, 95, 15769–15774. [Google Scholar] [CrossRef] [PubMed]
- Chiarini, A.; Gui, L.; Viviani, C.; Armato, U.; Dal Pra, I. NLRP3 Inflammasome’s Activation in Acute and Chronic Brain Diseases-An Update on Pathogenetic Mechanisms and Therapeutic Perspectives with Respect to Other Inflammasomes. Biomedicines 2023, 11, 999. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Guo, M.; Ma, R.; Wu, M.; Zhang, Y. Tetrandrine alleviates cerebral ischemia/reperfusion injury by suppressing NLRP3 inflammasome activation via Sirt-1. PeerJ 2020, 8, e9042. [Google Scholar] [CrossRef]
- Zhao, J.; Piao, X.; Wu, Y.; Liang, S.; Han, F.; Liang, Q.; Shao, S.; Zhao, D. Cepharanthine attenuates cerebral ischemia/reperfusion injury by reducing NLRP3 inflammasome-induced inflammation and oxidative stress via inhibiting 12/15-LOX signaling. Biomed. Pharmacother. 2020, 127, 110151. [Google Scholar] [CrossRef]
- Shim, D.W.; Lee, K.H. Posttranslational Regulation of the NLR Family Pyrin Domain-Containing 3 Inflammasome. Front. Immunol. 2018, 9, 1054. [Google Scholar] [CrossRef]
- Chen, J.; Chen, Z.J. PtdIns4P on dispersed trans-Golgi network mediates NLRP3 inflammasome activation. Nature 2018, 564, 71–76. [Google Scholar] [CrossRef]
- Iyer, S.S.; Pulskens, W.P.; Sadler, J.J.; Butter, L.M.; Teske, G.J.; Ulland, T.K.; Eisenbarth, S.C.; Florquin, S.; Flavell, R.A.; Leemans, J.C.; et al. Necrotic cells trigger a sterile inflammatory response through the Nlrp3 inflammasome. Proc. Natl. Acad. Sci. USA 2009, 106, 20388–20393. [Google Scholar] [CrossRef]
- He, Q.; Li, Z.; Wang, Y.; Hou, Y.; Li, L.; Zhao, J. Resveratrol alleviates cerebral ischemia/reperfusion injury in rats by inhibiting NLRP3 inflammasome activation through Sirt1-dependent autophagy induction. Int. Immunopharmacol. 2017, 50, 208–215. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Ye, Y.; Zhu, H.; Zhang, J.; Wang, H.; Lei, J.; Gu, L.; Zhan, L. NLRP3 inflammasome deficiency attenuates cerebral ischemia-reperfusion injury by inhibiting ferroptosis. Brain Res. Bull. 2023, 193, 37–46. [Google Scholar] [CrossRef]
- Chu, X.; Zhang, L.; Zhou, Y.; Fang, Q. Cucurbitacin B alleviates cerebral ischemia/reperfusion injury by inhibiting NLRP3 inflammasome-mediated inflammation and reducing oxidative stress. Biosci. Biotechnol. Biochem. 2022, 86, 846–854. [Google Scholar] [CrossRef]
- Wang, Y.; Meng, C.; Zhang, J.; Wu, J.; Zhao, J. Inhibition of GSK-3beta alleviates cerebral ischemia/reperfusion injury in rats by suppressing NLRP3 inflammasome activation through autophagy. Int. Immunopharmacol. 2019, 68, 234–241. [Google Scholar] [CrossRef]
- Lee, T.K.; Lee, J.C.; Kim, J.D.; Kim, D.W.; Ahn, J.H.; Park, J.H.; Kim, H.I.; Cho, J.H.; Choi, S.Y.; Won, M.H.; et al. Populus tomentiglandulosa Extract Is Rich in Polyphenols and Protects Neurons, Astrocytes, and the Blood-Brain Barrier in Gerbil Striatum Following Ischemia-Reperfusion Injury. Molecules 2021, 26, 5430. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, T.K.; Kim, D.W.; Ahn, J.H.; Lee, C.H.; Kim, J.D.; Shin, M.C.; Cho, J.H.; Lee, J.C.; Won, M.H.; et al. Astaxanthin Confers a Significant Attenuation of Hippocampal Neuronal Loss Induced by Severe Ischemia-Reperfusion Injury in Gerbils by Reducing Oxidative Stress. Mar. Drugs 2022, 20, 267. [Google Scholar] [CrossRef]
- Radtke-Schuller, S.; Schuller, G.; Angenstein, F.; Grosser, O.S.; Goldschmidt, J.; Budinger, E. Brain atlas of the Mongolian gerbil (Meriones unguiculatus) in CT/MRI-aided stereotaxic coordinates. Brain Struct. Funct. 2016, 221 (Suppl. S1), 1–272. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, T.K.; Kim, D.W.; Sim, H.; Lee, J.C.; Kim, J.D.; Ahn, J.H.; Lee, C.H.; Kim, Y.M.; Won, M.H.; et al. Neuroprotective Effects of Salicin in a Gerbil Model of Transient Forebrain Ischemia by Attenuating Oxidative Stress and Activating PI3K/Akt/GSK3beta Pathway. Antioxidants 2021, 10, 629. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.W.; Lee, T.-K.; Ahn, J.H.; Yang, S.-R.; Shin, M.C.; Cho, J.H.; Won, M.-H.; Kang, I.J.; Park, J.H. Porphyran Attenuates Neuronal Loss in the Hippocampal CA1 Subregion Induced by Ischemia and Reperfusion in Gerbils by Inhibiting NLRP3 Inflammasome-Mediated Neuroinflammation. Mar. Drugs 2024, 22, 170. https://doi.org/10.3390/md22040170
Kim DW, Lee T-K, Ahn JH, Yang S-R, Shin MC, Cho JH, Won M-H, Kang IJ, Park JH. Porphyran Attenuates Neuronal Loss in the Hippocampal CA1 Subregion Induced by Ischemia and Reperfusion in Gerbils by Inhibiting NLRP3 Inflammasome-Mediated Neuroinflammation. Marine Drugs. 2024; 22(4):170. https://doi.org/10.3390/md22040170
Chicago/Turabian StyleKim, Dae Won, Tae-Kyeong Lee, Ji Hyeon Ahn, Se-Ran Yang, Myoung Cheol Shin, Jun Hwi Cho, Moo-Ho Won, Il Jun Kang, and Joon Ha Park. 2024. "Porphyran Attenuates Neuronal Loss in the Hippocampal CA1 Subregion Induced by Ischemia and Reperfusion in Gerbils by Inhibiting NLRP3 Inflammasome-Mediated Neuroinflammation" Marine Drugs 22, no. 4: 170. https://doi.org/10.3390/md22040170
APA StyleKim, D. W., Lee, T. -K., Ahn, J. H., Yang, S. -R., Shin, M. C., Cho, J. H., Won, M. -H., Kang, I. J., & Park, J. H. (2024). Porphyran Attenuates Neuronal Loss in the Hippocampal CA1 Subregion Induced by Ischemia and Reperfusion in Gerbils by Inhibiting NLRP3 Inflammasome-Mediated Neuroinflammation. Marine Drugs, 22(4), 170. https://doi.org/10.3390/md22040170