Impact of Marine Drugs on Animal Reproductive Processes

Abstract
:1. Introduction
1.1. Biology of Reproduction
1.2. Marine Drugs
1.2.1. Marine Natural Products Affecting Reproduction
1.2.2. Potential Impact of Marine Drugs
Acknowledgments
References
- Yanagimachi, R. Mammalian fertilization. In The Physiology of Reproduction; Knobil, E, Neil, J, Eds.; Raven Press: New York, NY, USA, 1994; Volume 1, pp. 189–317. [Google Scholar]
- Eppig, JJ. Coordination of nuclear and cytoplasmic oocyte maturation in eutherian mammals. Reprod Fertil Dev 1996, 8, 485–489. [Google Scholar]
- Masui, Y. A quest for cytoplasmic factors that control the cell cycle. Prog Cell Cycle Res 1996, 2, 1–13. [Google Scholar]
- Whitaker, M; Patel, R. Calcium and cell cycle control. Development 1990, 108, 525–542. [Google Scholar]
- Whitaker, M. Control of meiotic arrest. Rev Reprod 1996, 1, 127–135. [Google Scholar]
- Kishimoto, T. Cell-cycle control during meiotic maturation. Curr Opin Cell Biol 2003, 15, 654–663. [Google Scholar]
- Haccard, O; Jessus, C. Oocyte maturation, Mos and cyclins-A matter of synthesis: Two functionally redundant ways to induce meiotic maturation. Cell Cycle 2006, 5, 1152–1159. [Google Scholar]
- Ducibella, TA; Anderson, DF; Albertini, F; Aalberg, J; Rangarajan, S. Quantitative studies of changes in cortical granule number and distribution in the mouse oocyte during maturation. Dev Biol 1988, 130, 184–197. [Google Scholar]
- Wessel, GM; Brooks, JM; Green, E; Haley, S; Voronina, E; Wong, J; Zaydfudim, V; Conner, S. The biology of cortical granules. Int Rev Cytol 2001, 209, 117–206. [Google Scholar]
- Prodon, F; Chenevert, J; Sardet, C. Establishment of animal-vegetal polarity during maturation in ascidian oocytes. Dev Biol 2006, 290, 297–311. [Google Scholar]
- Hake, LE; Richter, JD. Translational regulation of maternal mRNA. Biochem Biophys Acta 1997, 1332, 31–38. [Google Scholar]
- Carroll, J. Na+-Ca2+ exchange in mouse oocytes: Modifications in the regulation of intracellular free Ca2+ during oocyte maturation. J Reprod Fert 2000, 118, 337–342. [Google Scholar]
- Cuomo, A; Di Cristo, C; Di Cosmo, A; Paolucci, M; Tosti, E. Calcium currents correlate with oocyte maturation during the reproductive cycle in. Octopus vulgaris J Exp Zool A 2005, 303, 193–202. [Google Scholar]
- Cuomo, A; Silvestre, F; De Santis, R; Tosti, E. Ca2+ and Na+ current patterns during oocyte maturation, fertilization, and early developmental stages of Ciona intestinalis. Mol Reprod Dev 2006, 73, 501–511. [Google Scholar]
- Silvestre, F; Cuomo, A; Tosti, E. Ion current activity and molecules modulating maturation and growth stages of ascidian (Ciona intestinalis) oocytes. Mol Repodr Dev 2009, 76, 1084–1093. [Google Scholar]
- Homa, S. Calcium and meiotic maturation of the mammalian oocyte. Mol Reprod Dev 1995, 40, 122–134. [Google Scholar]
- Gilchrist, R; Thompson, JG. Oocyte maturation: Emerging concepts and technologies to improve developmental potential in vitro. Theriogenology 2007, 67, 6–15. [Google Scholar]
- Richard, FJ. Regulation of meiotic maturation. J Animal Sci 2007, 85, E4–E6. [Google Scholar]
- Vaccari, S; Horner, K; Mehlmann, LM; Conti, M. Generation of mouse oocytes defective in cAMP synthesis and degradation: Endogenous cyclic AMP is essential for meiotic arrest. Dev Biol 2008, 316, 124–134. [Google Scholar]
- Masui, Y. From oocyte maturation to the in vitro cell cycle: The history of discoveries of Maturation-Promoting Factor (MPF) and Cytostatic Factor (CSF). Differentiation 2001, 69, 1–17. [Google Scholar]
- Russo, GL; Kyozuka, K; Antonazzo, L; Tosti, E; Dale, B. Maturation promoting factor in ascidian oocytes is regulated by different intracellular signals at meiosis I and II. Development 1996, 122, 1995–2003. [Google Scholar]
- Kishimoto, T. Cell cycle arrest and release in starfish oocytes and eggs. Semin Cell Dev Biol 1998, 9, 549–557. [Google Scholar]
- Yamashita, M; Mita, K; Yoshida, N; Kondo, T. Molecular mechanisms of the initiation of oocyte maturation: General and species-specific aspects. Prog Cell Cycle Res 2000, 4, 115–129. [Google Scholar]
- Tosti, E. Calcium ion currents mediating oocyte maturation events. Reprod Biol Endocrinol 2006, 4, 26. [Google Scholar]
- Whitaker, M. Calcium at fertilization and in early development. Physiol Rev 2006. [Google Scholar]
- Carroll, J; Swann, K; Whittingham, D; Whitaker, M. Spatiotemporal dynamics of intracellular [Ca2+]i oscillations during the growth and meiotic maturation of mouse oocytes. Development 1994, 120, 3507–3517. [Google Scholar]
- Tombes, RM; Simerly, C; Borisy, GG; Schatten, G. Meiosis, egg activation, and nuclear envelope breakdown are differentially reliant on Ca2+, whereas germinal vesicle breakdown is Ca2+ independent in the mouse oocyte. J Cell Biol 1992, 117, 799–811. [Google Scholar]
- Roth, Z; Hansen, PJ. Disruption of nuclear maturation and rearrangement of cytoskeletal elements in bovine oocytes exposed to heat shock during maturation. Reproduction 2005, 129, 235–244. [Google Scholar]
- Sun, QY; Schatten, H. Regulation of dynamic events by microfilaments during oocyte maturation and fertilization. Reproduction 2006, 131, 193–205. [Google Scholar]
- Tosti, E. Sperm activation in species with external fertilization. Zygote 1994, 2, 359–361. [Google Scholar]
- Stratmhann, RR. Why life histories evolve differently in the sea. Soc Integ Comp Biol 1990, 30, 197–207. [Google Scholar]
- Levitan, DR; Young, CM. Reproductive success in large populations: Empirical measures and theoretical predictions of fertilization in the sea biscuit Clypeaster rosaceus. J Exp Mar Biol Ecol 1995, 190, 221–241. [Google Scholar]
- Sewell, MA; Levitan, DR. Fertilization success during a natural spawning of the dendrochirote sea cucumber Cucumaria miniata. Bull Mar Sci 1992, 51, 161–166. [Google Scholar]
- Mead, KS; Denny, MW. The effects of hydrodynamic shear stress on fertilization and early development of the purple sea urchin Strongylocentrotus purpuratus. Biol Bull 1995, 188, 46–56. [Google Scholar]
- Thomas, F. Physical properties of gametes in three sea urchin species. J Exp Biol 1994, 194, 263–284. [Google Scholar]
- Dale, B. Fertilization in animals. Arnold, Edward, Ed.; The Camelot Pres Ltd: London, UK, 1983. [Google Scholar]
- Aitken, RJ. Sperm function tests and fertility. Int J Androl 2006, 29, 69–75. [Google Scholar]
- Breitbart, H. Signaling pathways in sperm capacitation and acrosome reaction. Cell Mol Biol 2003, 49, 321–327. [Google Scholar]
- Stein, KK; Primakoff, P; Myles, D. Sperm-egg fusion: Events at the plasma membrane. J Cell Sci 2004, 1117, 6269–6274. [Google Scholar]
- Tosti, E; Boni, R. Electrical events during gamete maturation and fertilisation in animals and human. Hum Reprod Update 2004, 10, 53–65. [Google Scholar]
- Dale, B; DeFelice, LJ; Taglietti, V. Membrane noise and conductance increase during single spermatozoon-egg interaction. Nature 1978, 275, 217–219. [Google Scholar]
- Dale, B; de Santis, A. Maturation and fertilization of the sea urchin oocyte: An electrophysiological study. Dev Biol 1981, 85, 474–484. [Google Scholar]
- Dale, B; DeFelice, LJ. Sperm-activated channels in ascidian oocytes. Dev Biol 1984, 101, 235–239. [Google Scholar]
- DeFelice, LJ; Kell, MJ. Sperm-activated currents in ascidian oocytes. Dev Biol 1987, 119, 123–128. [Google Scholar]
- Jaffe, LA; Gould-Somero, M; Holland, L. Ionic mechanism of the fertilization potential of the marine worm, Urechis caupo (Echiura). J Gen Physiol 1979, 73, 469–492. [Google Scholar]
- Glahn, D; Nuccitelli, R. Voltage-clamp study of the activation currents and fast block to polyspermy in the egg of Xenopus laevis. Dev Growth Diff 2003, 45, 187–197. [Google Scholar]
- Miyazaki, S; Igusa, Y. Fertilization potential in golden hamster eggs consists of recurring hyperpolarization. Nature 1981, 290, 702–704. [Google Scholar]
- McCulloh, DH; Rexroad, CE, Jr; Levitan, H. Insemination of rabbit eggs is associated with slow depolarization and repetitive diphasic membrane potentials. Dev Biol 1983, 95, 372–377. [Google Scholar]
- Igusa, Y; Miyazaki, S; Yamashita, N. Periodic hyperpolarizing responses in hamster and mouse eggs fertilized with mouse sperm. J Physiol 1983, 340, 633–647. [Google Scholar]
- Gianaroli, L; Tosti, E; Magli, C; Iaccarino, M; Ferraretti, AP; Dale, B. Fertilization current in the human oocyte. Mol Reprod Dev 1994, 38, 209–214. [Google Scholar]
- Tosti, E; Boni, R; Cuomo, A. Fertilization and activation currents in bovine oocytes. Reproduction 2002, 124, 835–846. [Google Scholar]
- Jaffe, LA. Fast block to polyspermy in sea urchin eggs is electrically mediated. Nature 1976, 261, 68–71. [Google Scholar]
- Gould-Somero, M; Jaffe, LA; Holland, LZ. Electrically mediated fast polyspermy block in eggs of the marine worm. Urechis caupo J Cell Biol 1979, 82, 426–440. [Google Scholar]
- Goudeau, H; Depresle, Y; Rosa, A; Goudeau, M. Evidence by a voltage clamp study of an electrically mediated block to polyspermy in the egg of the ascidian Phallusia mammillata. Dev Biol 1994, 166, 489–501. [Google Scholar]
- Satoh, N. Developmental Biology of Ascidians; Cambridge University Press: New York, NY, USA, 1994. [Google Scholar]
- Turner, PR; Jaffe, LA; Fein, A. Regulation of cortical vesicle exocytosis in sea urchin eggs by inositol 1,4,5-trisphosphate and GTP-binding protein. J Cell Biol 1986, 102, 70–76. [Google Scholar]
- Stith, BJ; Espinoza, R; Roberts, D; Smart, T. Sperm increase inositol 1,4,5-trisphosphate mass in Xenopus laevis eggs preinjected with calcium buffers or heparin. Dev Biol 1994, 165, 206–215. [Google Scholar]
- Dupont, G; McGuinness, OM; Johnson, MH; Berridge, MJ; Borgese, F. Phospholipase C in mouse oocytes: Characterization of beta and gamma isoforms and their possible involvement in sperm-induced Ca2+ spiking. Biochem J 1996, 316, 583–591. [Google Scholar]
- Lee, SJ; Madden, PJ; Shen, SS. U73122 blocked the cGMP-induced calcium release in sea urchin eggs. Exp Cell Res 1998, 242, 328–334. [Google Scholar]
- Kline, D; Kline, JT. Repetitive calcium transients and the role of calcium in exocytosis and cell cycle activation in the mouse egg. Dev Biol 1992, 149, 80–89. [Google Scholar]
- Miyazaki, S; Shirakawa, H; Nakada, K; Honda, Y. Essential role of the inositol 1,4,5-trisphosphate receptor/Ca2+ release channel in Ca2+ waves and Ca2+ oscillations at fertilization of mammalian eggs. Dev Biol 1993, 158, 62–78. [Google Scholar]
- Swann, K; Ozil, JP. Dynamics of the calcium signal that triggers mammalian egg activation. Int Rev Cytol 1994, 152, 183–222. [Google Scholar]
- Xu, Z; Kopf, GS; Schultz, RM. Involvement of inositol 1,4,5-trisphosphate-mediated Ca2+ release in early and late events of mouse egg activation. Development 1994, 120, 1851–1859. [Google Scholar]
- Abbott, AL; Ducibella, T. Calcium and the control of mammalian cortical granule exocytosis. Front Biosci 2001, 6, D792–D806. [Google Scholar]
- Malcuit, C; Kurokawa, M; Fissore, RA. Calcium oscillations and mammalian egg activation. J Cell Physiol 2006, 206, 565–573. [Google Scholar]
- Busa, WB; Nuccitelli, R. An elevated free cytosolic Ca2+ wave follows fertilization in eggs of the frog, Xenopus laevis. J Cell Biol 1985, 100, 1325–329. [Google Scholar]
- Jaffe, LA; Giusti, AF; Carroll, DJ; Foltz, KR. Ca2+ signalling during fertilization of echinoderm eggs. seminars in Cell & Dev. Biol 2001, 12, 45–51. [Google Scholar]
- Dumollard, R; McDougall, A; Rouvière, C; Sardet, C. Fertilisation calcium signals in the ascidian egg. Biol Cell 2004, 96, 29–36. [Google Scholar]
- He, CL; Damiani, P; Parys, JB; Fissore, RA. Calcium, calcium release receptors, and meiotic resumption in bovine oocytes. Biol Reprod 1997, 57, 1245–1255. [Google Scholar]
- Mehlmann, LM; Kline, D. Regulation of intracellular calcium in the mouse egg: Calcium release in response to sperm or inositol trisphosphate is enhanced after meiotic maturation. Biol Reprod 1994, 51, 1088–1098. [Google Scholar]
- Ducibella, T; Huneau, D; Angelichio, E; Xu, Z; Schultz, RM; Kopf, GS; Fissore, RA; Madoux, S; Ozil, JP. Egg-to-embryo transition is driven by differential responses to Ca2+ oscillation number. Dev Biol 2002, 250, 280–291. [Google Scholar]
- Miyazaki, S; Ito, M. Calcium signals for egg activation in mammals. J Pharmacol Sci 2006, 100, 545–552. [Google Scholar]
- Swann, K; Yu, Y. The dynamics of calcium oscillations that activate mammalian eggs. Int J Dev Biol 2008, 52, 585–594. [Google Scholar]
- Dale, B. Oocyte activation in invertebrates and humans. Zygote 1994, 2, 373–377. [Google Scholar]
- Runft, LL; Jaffe, LA; Mehlmann, LM. Egg activation at fertilization: Where it all begins. Dev Biol 2002, 245, 237–54. [Google Scholar]
- Wilding, M; Dale, B. Sperm factor: What is and what I does it do? Mol Human Reprod 1997, 3, 269–273. [Google Scholar]
- Swann, K; Larman, MG; Saunders, CM; Lai, FA. The cytosolic sperm factor that triggers Ca2+ oscillations and egg activation in mammals is a novel phospholipase C: PLCzeta. Reproduction 2004, 127, 431–439. [Google Scholar]
- Parrington, J; Swann, K; Shevchenko, VI; Sesay, AK; Lai, FA. Calcium oscillations in mammalian eggs triggered by a soluble sperm protein. Nature 1996, 379, 364–368. [Google Scholar]
- Sette, C; Bevilacqua, A; Bianchini, A; Mangia, F; Geremia, R; Rossi, P. Parthenogenetic activation of mouse eggs by microinjection of a truncated c-kit tyrosine kinase present in spermatozoa. Development 1997, 124, 2267–2274. [Google Scholar]
- Saunders, CM; Larman, MG; Parrington, J; Cox, LJ; Royse, J; Blayney, LM; Swann, K; Lai, FA. PLC zeta: A sperm-specific trigger of Ca(2+) oscillations in eggs and embryo development. Development 2002, 129, 3533–3544. [Google Scholar]
- Tosti, E; Menezo, Y. Sperm induced oocyte activation. In Human Spermatozoa: Maturation, Capacitation and Abnormalities; Nova Science Publishers: Hauppage, NY, USA; , in press.
- Alberts, B; Bray, D; Lewis, J; Raff, M; Roberts, K; Watson, JD. Cellular mechanisms of development. In Molecular Biology of the Cell; Garland Publishing: New York, NY, USA, 1983; pp. 813–890. [Google Scholar]
- Menezo, Y; Renard, JP. The life of egg before implantation. In Reproduction in Mammals and Man; Thibault, C, Levassouf, MC, Eds.; AHF Unter Elipses: Paris, France, 1993; pp. 350–366. [Google Scholar]
- Vacelet, J; Donadey, C. Electron microscope study of the association between some sponges and bacteria. J Exp Mar Ecol 1977, 30, 301–314. [Google Scholar]
- Wilkinson, CR. Symbiotic interactions between marine sponges and algae. In Algae and Symbioses; Reisser, W, Ed.; Biopress: Bristol, UK, 1992; pp. 112–151. [Google Scholar]
- Hentschel, U; Steinert, M; Hacker, J. Common molecular mechanisms of symbiosis and pathogenesis. Trends Microbiol 2000, 8, 226–231. [Google Scholar]
- Steinert, M; Hentschel, U; Hacker, J. Symbiosis and pathogenesis: Evolution of the microbe-host interaction. Naturwissenschaft 2000, 7, 1–11. [Google Scholar]
- Proksch, P; Edrada, RA; Ebel, R. Drugs from the seas - Current status and microbiological implications. Appl Microbiol Biotechnol 2002, 59, 125–134. [Google Scholar]
- Berry, JP; Gantar, M; Perez, MH; Berry, G; Noriega, FG. Cyanobacterial toxins as allelochemicals with potential applications as algaecides, herbicides and insecticides. Mar Drugs 2008, 6, 117–146. [Google Scholar]
- van Apeldoorn, ME; van Egmond, HP; Speijers, GJ; Bakker, GJ. Toxins of cyanobacteria. Mol Nutr Food Res 2007, 51, 7–60. [Google Scholar]
- Duy, TN; Lam, PKS; Shaw, G; Connell, DW. Toxicology and risk assessment of freshwater cyanobacterial (bluegreen algal) toxins in water. Rev Environ Contam Toxicol 2000, 163, 113–186. [Google Scholar]
- Miyazawa, K; Noguchi, T. Distribution and origin of tetrodotoxin. J ToxicolTox Rev 2001, 20, 11–33. [Google Scholar]
- Noguchi, T; Jeon, JK; Arakawa, O; Sugita, H; Deguchi, Y; Shida, Y; Hashimoto, K. Occurrence of Tetrodotoxin and Anhydrotetrodotoxin in Vibrio sp. Isolated from the Intestines of a Xanthid Crab, Atergatis floridus. J Biochem 1986, 99, 311–314. [Google Scholar]
- Narita, H; Matsubara, S; Miwa, N; Akahane, S; Murakami, M; Goto, T; Nara, M; Noguchi, T; Saito, T; Shida, Y; Hashimoto, K. Vibrio alginolyticus, a TTX-producing bacterium isolated from the starfish Astropecten polyacanthus. Bull Japan Soc Sci Fish 1987, 47, 935–941. [Google Scholar]
- Hashimoto, K; Noguchi, T; Watabe, S. Microbial Toxins in Foods and Feeds; Pohland, AE, Dowell, VR, Jr, Richard, JL, Eds.; Plenum Press: New York, NY, USA, 1990; pp. 159–172. [Google Scholar]
- Naharashi, T. Tetrodotoxin: A brief history. Proc Jpn Acad Ser B Phys Biol Sci 2008, 84, 147–154. [Google Scholar]
- Yasumoto, T; Murata, M. Marine toxins. Chem Rev 1993, 93, 1897–1909. [Google Scholar]
- Yasumoto, T; Satake, M. Chemistry, Etiology and Determination Methods of Ciguatera Toxins. J Toxicol Tox Rev 1996, 15, 91–107. [Google Scholar]
- Yasumoto, T. The Chemistry and Biological Function of Natural Marine Toxins. Chem Rec 2001, 1, 228–242. [Google Scholar]
- Takahashi, M; Ohizumi, Y; Yasumoto, T. Maitotoxin, a Ca2+ channel activator candidate. J Biol Chem 1982, 257, 7287–7289. [Google Scholar]
- Pesando, D; Girard, JP; Durand-Clément, M; Payan, P; Puiseux-Dao, S. Effect of maitotoxin on sea urchin egg fertilization and on Ca2+ permeabilities of eggs and intracellular stores. Biol Cell 1991, 72, 269–273. [Google Scholar]
- Escobar, LI; Salvador, C; Martinez, M; Vaca, L. Maitotoxin, a cationic channel activator. Neurobiology 1998, 6, 59–74. [Google Scholar]
- Treviño, CL; De la Vega-Beltrán, JL; Nishigaki, T; Felix, R; Darszon, A. Maitotoxin potently promotes Ca2+ influx in mouse spermatogenic sells and sperm, and snduces the acrosome reaction. J Cell Physiol 2006, 206, 449–456. [Google Scholar]
- Yasumoto, T; Seino, N; Murakami, Y; Murata, M. Toxins produced by benthic dinoflagellates. Biol Bull 1987, 172, 128–131. [Google Scholar]
- Dounay, AB; Forsyth, CJ. Okadaic acid: The archetypal serine/threonine protein phosphatase inhibitor. Curr Med Chem 2002, 9, 1939–1980. [Google Scholar]
- Cohen, P; Holmes, CFB; Tsukitani, Y. Okadaic acid: A new probe for studying cellular regulation. Trends Biochem Sci 1990, 15, 98–102. [Google Scholar]
- Tachibana, K; Scheuer, P; Tsukitani, Y; Kikuchi, H; van Engen, D; Clardy, J; Gopichand, Y; Schmitz, FJ. Okadaic acid, a cytotoxic polyether from two marine sponges of the genus Halichondria. J Am Chem Soc 1981, 103, 2469–2471. [Google Scholar]
- Murakami, Y; Oshima, Y; Yasumoto, T. Identification of okadaic acid as a toxic component of a marine dinoflagellate. Prorocentrum lima Bull Jpn Soc Sci Fish 1982, 48, 69–72. [Google Scholar]
- Zhou, J; Fritz, L. Okadaic acid antibody localizes to chloroplasts in the DSP-toxin-producing dinoflagellates Prorocentrum lima and Prorocentrum maculosum. Phycologia 1994, 33, 455–461. [Google Scholar]
- McLachlan, JL; Marr, JC; Conlon-Kelly, A; Adamson, A. Effects of nitrogen concentration and cold temperature on DSP-toxin concentrations in the dinoflagellate Prorocentrum lima (Prorocentrales, Dinophyceae). J Nat Toxins 1994, 2, 263–270. [Google Scholar]
- Wiens, M; Luckas, B; Brümmer, F; Shokry, M; Ammar, A; Steffen, R; Batel, R; Diehl-Seifert, B; Schröder, HC; Müller, WEG. Okadaic acid: A potential defense molecule for the sponge Suberites domuncula. Mar Biol 2003, 142, 213–223. [Google Scholar]
- Schröder, HC; Breter, HJ; Fattorusso, E; Ushijima, H; Wiens, M; Steffen, R; Batel, R; Müller, WEG. Okadaic acid, an apoptogenic toxin for symbiotic/parasitic annelids in the demosponge. Suberites domuncula Appl Environ Microbiol 2006, 72, 4907–4916. [Google Scholar]
- Paul, VJ; Arthur, KE; Ritson-Williams, R; Ross, C; Sharp, K. Chemical defenses: From compounds to communities. Biol Bull 2007, 213, 226–251. [Google Scholar]
- Pondaven, P; Cohen, P. Identification of protein phosphatases-l and 2A and inhibitor-2 in oocytes of the starfish Asterias rubens and Marthasterias glacialis. Eur J Biochem 1987, 167, 135–140. [Google Scholar]
- Goris, J; Hermann, J; Hendrix, P; Ozon, R; Merlevede, W. Okadaic acid, a specific protein phosphatase inhibitor, induces maturation and MPF formation in Xenopus laevis oocytes. FEBS Lett 1989, 245, 91–94. [Google Scholar]
- Rime, H; Ozon, R. Protein phosphatases are involved in the in vivo activation of histone HI kinase in mouse oocyte. Dev Biol 1990, 141, 115–122. [Google Scholar]
- Alexandre, H; van Cauwenberge, A; Tsukitani, Y; Mulnard, J. Pleiotropic effect of okadaic acid on maturing mouse oocytes. Development 1991, 112, 971–980. [Google Scholar]
- Gavin, AC; Tsukitani, Y; Schorderet-Slatkine, S. Induction of M-phase entry of prophase-blocked mouse oocytes through microinjection of okadaic acid, a specific phosphatase inhibitor. Exp Cell Res 1991, 192, 75–81. [Google Scholar]
- Schwartz, DA; Schultz, RM. Stimulatory effect of okadaic acid, an inhibitor of protein phosphatases, on nuclear envelope breakdown and protein phosphorylation in mouse oocytes and one-cell embryos. Dev Biol 1991, 145, 119–127. [Google Scholar]
- Gavin, AC; Vassalli, JD; Cavadore, JC; Schorderet-Slatkine, S. Okadaic acid and p13suc1 modulate the reinitiation of meiosis in mouse oocytes. Mol Reprod Dev 1992, 33, 287–296. [Google Scholar]
- Levesque, JT; Sirard, MA. Effects of different kinases and phosphatases on nuclear and cytoplasmic maturation of bovine oocytes. Mol Reprod Dev 1995, 42, 114–121. [Google Scholar]
- Sasseville, M; Côté, N; Guillemette, C; Richard, FJ. New insight into the role of phosphodiesterase 3A in porcine oocyte maturation. BMC Dev Biol 2006, 6, 47. [Google Scholar]
- Swain, JE; Ding, J; Brautigan, DL; Villa-Moruzzi, E; Smith, GD. Proper chromatin condensation and maintenance of histone H3 phosphorylation during mouse oocyte meiosis requires protein phosphatase activity. Biol Reprod 2007, 76, 628–638. [Google Scholar]
- Kimm-Brinson, KL; Ramsdell, JS. The red tide toxin, brevetoxin, induces embryo toxicity and developmental abnormalities. Environ Health Perspect 2001, 109, 377–381. [Google Scholar]
- Moon, RT; Morrill, JB. The effects of Gymnodinium breve lysate on the larval development of the sea urchin Lytechinus variegatus. J Environ Sci Health 1976, 11, 673–683. [Google Scholar]
- Steidinger, KA; Burklew, MA; Ingle, RM. The effects of Gymnodinium breve toxin on estuarine animals. In Marine Pharmacognosy; Martin, DF, Padilla, GM, Eds.; Academic Press: New York, NY, USA, 1973; pp. 179–202. [Google Scholar]
- Miller, MA. Maternal transfer of lipophilic contaminants in Salmonines to their eggs. Can J Fish Aquat Sci 1993, 49, 1405–1413. [Google Scholar]
- Walker, ST. Fish mortality in the Gulf of Mexico. Proc US Natl Mus 1884, 6, 105–109. [Google Scholar]
- Taylor, HF. Mortality of fishes on the West Coast of Florida Report of the US Commissioner of Fisheries Bureau of Fisheries Document 848; Government Printing Office: Washington, USA, 1917. [Google Scholar]
- Davis, CC. Gymnodinium breve: A cause of discolored water and animal mortality in the Gulf of Mexico. Bot Gaz 1948, 109, 358–360. [Google Scholar]
- Gunter, G; Williams, RH; Davis, CC; Smith, FGW. Catastrophic mass mortality of marine animals and coincident phytoplankton bloom on the west coast of Florida, November 1946 to August 1947. Ecol Monogr 1948, 8, 310–324. [Google Scholar]
- Quick, JA; Henderson, GE. Evidences of new ichthyointoxicative phenomena in Gymnodinium breve red tides. In Proceedings of the First International Conference on Toxic Dinoflagellate Blooms; LoCicero, BR, Ed.; Massachusetts Science and Technology Foundation: Wakefield, UK, 1975; pp. 413–422. [Google Scholar]
- Riley, CM; Holt, SA; Holt, J; Buskey, EJ; Arnold, CR. Mortality of larval red drum (Sciaenops ocellatus) associated with a Ptychodiscus brevis red tide. Contrib Mar Sci 1989, 31, 137–146. [Google Scholar]
- Forrester, DJ; Gaskin, JM; White, FH; Thompson, NP; Quick, JA; Henderson, GE; Woodard, JC; Robertson, WD. An epizootic of waterfowl associated with a red tide episode in Florida. J Wildl Dis 1977, 13, 160–167. [Google Scholar]
- Bossart, GD; Baden, DG; Ewing, RY; Roberts, B; Wright, SD. Brevitoxicosis in manatees (Trichechus manatus latirostris) from the 1996 epizootic: Gross, histologic and mmunohistochemical features. Toxicol Pathol 1998, 26, 276–282. [Google Scholar]
- Flynn, KJ; Irigoien, X. Aldehyde-induced insidious effects cannot be considered as a diatom defence mechanism against copepods. Mar Ecol Prog Series 2009, 377, 79–89. [Google Scholar]
- Miralto, A; Barone, G; Romano, G; Poulet, SA; Ianora, A; Russo, GL; Buttino, I; Mazzarella, G; Laabir, M; Cabrini, M; Giacobbe, MG. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar]
- Ianora, A; Miralto, A; Poulet, SA; Carotenuto, Y; Buttino, I; Romano, G; Casotti, R; Pohnert, G; Wichard, T; Colucci-D’Amato, L; Terrazzano, G; Smetacek, V. Aldehyde suppression of copepod recruitment in blooms of a ubiquitous planktonic diatom. Nature 2004, 429, 403–407. [Google Scholar]
- Buttino, I; De Rosa, G; Carotenuto, Y; Mazzella, M; Ianora, A; Esposito, F; Vitiello, V; Quaglia, F; La Rotonda, MI; Miralto, A. Aldehyde-encapsulating liposomes impair marine grazer survivorship. J Exp Biol 2008, 211, 1426–1433. [Google Scholar]
- Caldwell, GS; Olive, PJW; Bentley, MG. Inhibition of embryonic development and fertilization in broadcast spawning marine invertebrates by water-soluble diatom extracts and the diatom toxin 2-trans,4-trans-decadienal. Aquatic Toxicol 2002, 60, 123–137. [Google Scholar]
- Caldwell, GS; Lewis, C; Olive, PJ; Bentley, MG. Exposure to 2,4-decadienal negatively impacts upon marine invertebrate larval fitness. Mar Environ Res 2005, 59, 405–417. [Google Scholar]
- Tosti, E; Romano, G; Buttino, I; Cuomo, A; Ianora, A; Miralto, A. Bioactive aldehydes from diatoms block the fertilization current in ascidian oocytes. Mol Reprod Dev 2003, 66, 72–80. [Google Scholar]
- Ramsdell, JS; Zabka, TS. In utero domoic acid toxicity: A fetal basis to adult disease in the California sea lion (Zalophus californianus). Mar Drugs 2008, 6, 262–290. [Google Scholar]
- Scholin, CA; Gulland, F; Doucette, GJ; Benson, S; Busman, M; Chavez, FP; Cordaro, J; DeLong, R; de Vogelaere, A; Harvey, J; Haulena, M; Lefebvre, K; Lipscomb, T; Loscutoff, S; Lowenstine, LJ; Marin, R, 3rd; Miller, PE; McLellan, WA; Moeller, PD; Powell, CL; Rowles, T; Silvagni, P; Silver, M; Spraker, T; Trainer, V; van Dolah, FM. Mortality of sea lions along the central California coast linked to a toxic diatom bloom. Nature 2000, 403, 80–84. [Google Scholar]
- Brodie, EC; Gulland, FMD; Greig, DJ; Hunter, M; Jaakola, J; Leger, JS; Leighfield, TA; van Dolah, FM. Domoic acid causes reproductive failure in California sea lions (Zalophus californianus). Mar Mam Sci 2006, 22, 700–707. [Google Scholar]
- Goldstein, T; Zabka, TS; Delong, RL; Wheeler, EA; Ylitalo, G; Bargu, S; Silver, M; Leighfield, T; van Dolah, F; Langlois, G; Sidor, I; Dunn, JL; Gulland, FM. The role of domoic acid in abortion and premature parturition of California sea lions (Zalophus californianus) on San Miguel Island, California. J Wildl Dis 2009, 45, 91–108. [Google Scholar]
- Lefebvre, KA; Powell, CL; Busman, M; Doucette, GJ; Moeller, PDR; Silver, JB; Miller, PE; Hughes, MP; Singaram, S; Silver, MW; Tjeerdema, RS. Detection of domoic acid in northern anchovies and California sea lions associated with an unusual mortality event. Nat Tox 1999, 7, 85–92. [Google Scholar]
- Lefebvre, KA; Dovel, SL; Silver, MW. Tissue distribution and neurotoxic effects of domoic acid in a prominent vector species, the northern anchovy Engraulis mordax. Mar Biol 2001, 138, 693–700. [Google Scholar]
- Kvitek, R; Goldberg, JD; Smith, GJ; Doucette, GJ; Silver, MW. Domoic acid contamination within eight representative species from the benthic food web of Monterey Bay, California, USA. Mar Ecol Prog Series 2008, 367, 35–47. [Google Scholar]
- Ramsdell, JS. The molecular and integrative basis to domoic acid toxicity. In Phycotoxins: Chemisty and Biochemistry; Botana, L, Ed.; Blackwell Publishing: Ames, IO, USA, 2007; pp. 223–250. [Google Scholar]
- Maucher, JM; Ramsdell, JS. Maternal-fetal transfer of domoic acid in rats at two gestational time points. Environ Health Perspect 2007, 115, 1743–1746. [Google Scholar]
- Liu, H; Kelly, MS; Campbell, DA; Dong, SL; Zhu, JX; Wang, SF. Exposure to domoic acid affects larval development of king scallop Pecten maximus (Linnaeus, 1758). Aquat. Toxicol 2007, 81, 152–158. [Google Scholar]
- Pesando, D; Huitorel, P; Dolcini, V; Amade, P; Girard, JP. Caulerpenyne interferes with microtubule-dependent events during the first mitotic cycle of sea urchin eggs. Eur J Cell Biol 1998, 77, 19–26. [Google Scholar]
- O’Brien, ET; Asai, DJ; Jacobs, RS; Wilson, L. Selective inhibition of cytokinesis in sea urchin embryos by low concentrations of stypoldione, a marine natural product that reacts with sulfhydryl groups. Mol Pharmacol 1989, 35, 635–642. [Google Scholar]
- Premakumara, GA; Ratnasooriya, WD; Tillekeratne, LM; Amarasekare, AS; Atta-Ur-Rahman. Human sperm motility stimulating activity of a sulfono glycolipid isolated from Sri Lankan marine red alga Gelidiella acerosa. Asian J Androl 2001, 3, 27–31. [Google Scholar]
- Ratnasooriya, WD; Premakumara, GA; Tillekeratne, LM. Post-coital contraceptive activity of crude extracts of Sri Lankan marine red algae. Contraception 1994. [Google Scholar]
- Williamson, JE; Carson, DG; de Nys, R; Steinberg, PD. Demographic consequences of an ontogenetic shift by a sea urchin in response to host plant chemistry. Ecology 2004, 85, 1355–1371. [Google Scholar]
- Ohta, E; Okada, H; Ohta, S; Kobayashi, M; Kitagawa, I; Horiike, S; Takahashi, T; Hosoya, H; Yamamoto, K; Ikegami, S. Malformation of immature starfish oocytes by theonellapeptolide Ie, a tridecapeptide lactone from a marine sponge Petrosia species, through disturbance of cortical F-actin distribution. Biosci. Biotechnol. Biochem 2003, 67, 1908–1915. [Google Scholar]
- Liu, H; Namikoshi, M; Meguro, S; Nagai, H; Kobayashi, H; Yao, X. Isolation and characterization of polybrominated diphenyl ethers as inhibitors of microtubule assembly from the marine sponge Phyllospongia dendyi collected at Palau. J Nat Prod 2004, 67, 472–474. [Google Scholar]
- Liu, H; Namikoshi, M; Akano, K; Kobayashi, H; Nagai, H; Yao, X. Seven new meroditerpenoids, from the marine sponge Strongylophora strongylata, that inhibited the maturation of starfish oocytes. J Asian Nat Prod Res 2005, 7, 661–670. [Google Scholar]
- Bubb, MR; Senderowicz, AM; Sausville, EA; Duncan, KL; Korn, ED. Jasplakinolide, a cytotoxic natural product, induces actin polymerization and competitively inhibits the binding of phalloidin to F-actin. J Biol Chem 1994, 269, 14869–14871. [Google Scholar]
- Terada, Y; Simerly, C; Schatten, G. Microfilament stabilization by jasplakinolide arrests oocyte maturation, cortical granule exocytosis, sperm incorporation cone resorption, and cell-cycle progression, but not DNA replication, during fertilization in mice. Mol Reprod Dev 2000, 56, 89–98. [Google Scholar]
- Schatten, G; Schatten, H; Spector, I; Cline, C; Paweletz, N; Simerly, C; Petzelt, C. Latrunculin inhibits the microfilament-mediated processes during fertilization, cleavage and early development in sea urchins and mice. Exp Cell Res 1986, 166, 191–208. [Google Scholar]
- Spector, I; Shochet, NR; Blasberger, D; Kashman, Y. Latrunculins--novel marine macrolides that disrupt microfilament organization and affect cell growth: I. Comparison with cytochalasin D. Cell Motil Cytoskeleton 1989, 13, 127–144. [Google Scholar]
- Ohta, E; Uy, MM; Ohta, S; Yanai, M; Hirata, T; Ikegami, S. Anti-fertilization activity of a spirocyclic sesquiterpene isocyanide isolated from the marine sponge Geodia exigua and related compounds. Biosci Biotechnol Biochem 2008, 72, 1764–1771. [Google Scholar]
- Kobayashi, M; Kanzaki, K; Katayama, S; Ohashi, K; Okada, H; Ikegami, S; Kitagawa, I. Marine natural products. XXXIII. Theonellapeptolide IId, a new tridecapeptide lactone from the Okinawan marine sponge Theonella swinhoei. Chem Pharm Bull (Tokyo) 1994, 42, 1410–1415. [Google Scholar]
- Ikegami, S; Kobayashi, H; Myotoishi, Y; Ohta, S; Kato, KH. Selective inhibition of exoplasmic membrane fusion in echinoderm gametes with jaspisin, a novel antihatching substance isolated from a marine sponge. J Biol Chem 1994, 269, 23262–23267. [Google Scholar]
- Ohta, S; Kobayashi, H; Ikegami, S. Jaspisin: A novel styryl sulfate from the marine sponge, Jaspis species. Biosci Biotechnol Biochem 1994, 58, 1752–1753. [Google Scholar]
- Kato, KH; Takemoto, K; Kato, E; Miyazaki, K; Kobayashi, H; Ikegami, S. Inhibition of sea urchin fertilization by jaspisin, a specific inhibitor of matrix metalloendoproteinase. Dev Growth Differ 1998, 40, 221–230. [Google Scholar]
- Uno, M; Ohta, S; Ohta, E; Ikegami, S. Callyspongins A and B: Novel polyacetylene sulfates from the marine sponge Callyspongia truncata that inhibit fertilization of starfish gametes. J Nat Prod 1996, 59, 1146–1148. [Google Scholar]
- Kobayashi, M; Shimizu, N; Kitagawa, I; Kyogoku, Y; Harada, N; Uda, H. Absolute stereostructures of halenaquinol and halenaquinol sulfate, pentacyclic hydroquinones from the Okinawan marine sponge Xestospongia sapra, as determined by theoretical calculation of CD spectra. Tetrahedron Lett 1985, 26, 3833–3836. [Google Scholar]
- Ikegami, S; Kajiyama, N; Ozaki, Y; Myotoishi, Y; Miyashiro, S; Takayama, S; Kobayashi, M; Kitagawa, I. Selective inhibition of membrane fusion events in echinoderm gametes and embryos by halenaquinol sulfate. FEBS Lett 1992, 302, 284–286. [Google Scholar]
- Ishihara, H. Calyculin A and okadaic acid: Inhibitors of protein phosphatase activity. Biochem Biophys Res Commun 1989, 159, 871–877. [Google Scholar]
- Smith, GD; Sadhu, A; Wolf, DP. Transient exposure of rhesus macaque oocytes to calyculin-A and okadaic acid stimulates germinal vesicle breakdown permitting subsequent development and fertilization. Biol Reprod 1998, 58, 880–886. [Google Scholar]
- Agius, L; Jaccarini, V; Ballantine, JA; Ferrito, V; Pelter, A; Psaila, AF; Zammit, VA. Photodynamic action of bonellin, an integumentary chlorin of Bonellia viridis, Rolando (Echiura, Bonelliidae). Comp Biochem Physiol B 1979, 63, 109–117. [Google Scholar]
- Pelter, A; Ballantine, JA; Ferrito, V; Jaccarino, V; Psaila, AF; Schembri, PJ. Bonellin, a most unusual chlorin. J Chem Soc Chem Comm 1976, 23, 999–1000. [Google Scholar]
- Cariello, L; De Nicola Giudici, M; Tosti, E; Zanetti, L. On the mechanism of action of bonellin on the sea urchin egg. Gamete Res 1982, 5, 161–166. [Google Scholar]
- De Nicola Giudici, M. Inhibition of motility by bonellin. II. Spermatozoa and embryos of sea urchin. Acta Embryol Morphol Exp 1982, 3, 97–106. [Google Scholar]
- Lakshmi, V; Saxena, A; Mishra, SK; Raghubir, R; Srivastava, MN; Jain, RK; Maikhuri, JP; Gupta, G. Spermicidal activity of bivittoside D from Bohadschia vitiensis. Arch Med Res 2008, 39, 631–638. [Google Scholar]
- Simon-Levert, A; Arrault, A; Bontemps-Subielos, N; Canal, C; Banaigs, B. Meroterpenes from the ascidian Aplidium aff densum. J Nat Prod 2005, 68, 1412–1415. [Google Scholar]
- Simon-Levert, A; Aze, A; Bontemps-Subielos, N; Banaigs, B; Genevière, AM. Antimitotic activity of methoxyconidiol, a meroterpene isolated from an ascidian. Chem Biol Interact 2007, 168, 106–116. [Google Scholar]
- Sipkema, D; Franssen, MC; Osinga, R; Tramper, J; Wijffels, RH. Marine sponges as pharmacy. Mar Biotechnol 2005, 7, 142–162. [Google Scholar]
- MarinLit. A marine literature database maintained by the Marine Chemistry Group. University of Canterbury: Christchurch, New Zealand, 1999. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat Prod Rep 2000, 17, 7–55. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat Prod Rep 2001, 18, 1–49. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat Prod Rep 2002, 19, 1–48. [Google Scholar]
- Wijffels, RH. Potential of sponges and microalgae for marine biotechnology. Trends Biotechnol 2007, 26, 26–31. [Google Scholar]
- Mayer, AMS; Gustafson, KR. Marine Pharmacology in 2005–6: Antitumor and Cytotoxic Compounds. Eur J Cancer 2008. [Google Scholar]
- Glaser, KB; Mayer, AMS. A Renaissance in Marine Pharmacology: From Preclinical Curiosity to Clinical Reality. Biochem Pharmacol 2009, 78, 440–448. [Google Scholar]
- Mayer, AMS; Rodriguez, AD; Berlinck, R; Hamann, MT. Marine pharmacology in 2005–6: Marine Compounds with Antibacterial, Anticoagulant, Antifungal, Anthelmitic, Anti-inflammatory, Antiprotozoal, and Antiviral Activities; affecting the Cardiovascular, Endocrine, Immune and Nervous Systems and other Miscellaneous Mechanisms of Action. Biochem Biophys Acta 2009, 1790, 283–308. [Google Scholar]
- Kitagawa, I; Kobayashi, M; Kitanaka, K; Kido, M; Kyogoku, Y. Marine natural products, XII: On the chemical constituents of the Okinawan marine sponge. Hymeniacidon aldis Chem Pharm Bull 1983, 31, 2321–2328. [Google Scholar]
- Fedoreev, SA; Prokof’eva, NG; Denisenko, VA; Rebachuk, NM. Cytotoxic activity of aaptamines from suberitid marine sponges. Pharm Chem J 1989, 22, 615–618. [Google Scholar]
- Willis, RH; De Vries, DJ. BRS1, a C30 bis-amino, bis-hydroxy polyunsaturated lipid from an Australian calcareous sponge that inhibits protein kinase C. Toxicon 1997, 35, 1125–1129. [Google Scholar]
- Jares-Erijman, EA; Sakai, R; Rinehart, KL. Crambescidins: New antiviral and cytotoxic compounds from the sponge Crambe crambe. J Ore Chem 1991, 56, 5712–5715. [Google Scholar]
- Berlinck, RGS; Braekman, JC; Daloze, D; Bruno, I; Riccio, R; Ferri, S; Spampinato, S; Speroni, E. Polycyclic guanidine alkaloids from the marine sponge Crambe crambe and Ca++ channel blocker activity of crambescidin 816. J Nat Prod 1993, 56, 1007–1015. [Google Scholar]
- Wang, DZ. Neurotoxins from marine dinoflagellates: A brief review. Mar Drugs 2008, 6, 349–71. [Google Scholar]
- Kem, WR. Sea anemone toxins: Structure and action. In The Biology of Nematocysts; Hessinger, DA, Lenhoff, HM, Eds.; Academic Press: New York, NY, USA, 1988; pp. 375–405. [Google Scholar]
- Kem, WR; Pennington, MW; Dunn, BM. Sea anemone polypeptide toxins affecting sodium channels. Initial structure-activity investigations. In Marine Toxins Origin, Structure and Molecular Pharmacology; Hall, S, Stricharz, G, Eds.; American Chemical Society: Washington, USA, 1990; pp. 279–289. [Google Scholar]
- Norton, TR. Cardiotonic polypeptides from Anthopleura xanthogrammica (Brandt) and A. elegantissima (Brandt). Fed Proc 1981, 40, 21–25. [Google Scholar]
- Honma, T; Shiomi, K. Peptide toxins in sea anemones: Structural and functional aspects. Mar Biotechnol 2006, 8, 1–10. [Google Scholar]
- Llewellyn, LE. Sodium channel inhibiting marine toxins. Prog Mol Subcell Biol 2009, 46, 67–97. [Google Scholar]
- Castañeda, O; Sotolongo, V; Amor, AM; Stöklin, R; Anderson, AJ; Harvey, AL; Engström, A; Wernstedt, C; Karlsson, E. Characterization of a potassium channel toxin from the Caribbean sea anemone. Stichodactyla helianthus Toxicon 1995, 33, 603–613. [Google Scholar]
- Schweitz, H; Bruhn, T; Guillemare, E; Moinier, D; Lancelin, J-M; Béress, L; Lazdunski, M. Kalicludines and kaliseptine. Two different classes of sea anemone toxins for voltage-sensitive K+ channels. J Biol Chem 1995, 270, 25121–25126. [Google Scholar]
- Cotton, J; Crest, M; Bouet, F; Alessandri, N; Gola, M; Forest, E; Karlsson, E; Castañeda, O; Harvey, AL; Vita, C; Ménez, A. A potassium-channel toxin from the sea anemone Bunodosoma granulifera, an inhibitor for Kv1 channels. Revision of the amino acid sequence, disulfide-bridge assignment, chemical synthesis, and biological activity. Eur J Biochem 1997, 244, 192–202. [Google Scholar]
- Gendeh, GS; Young, LC; de Medeiros, LC; Jeyaseelan, K; Harvey, AL; Chung, MCM. A new potassium channel toxin from the sea anemone Heteractis magnifica: Isolation, cDNA cloning, and functional expression. Biochemistry 1997, 36, 11461–11471. [Google Scholar]
- Minagawa, S; Ishida, M; Nagashima, Y; Shiomi, K. Primary structure of a potassium channel toxin from the sea anemone Actinia equina. FEBS Lett 1998, 427, 149–151. [Google Scholar]
- De Smet, P; Parys, JB; Callewaert, G; Weidema, AF; Hill, E; De Smedt, H; Erneux, C; Sorrentino, V; Missiaen, L. Xestospongin C is an equally potent inhibitor of the inositol 1,4,5-triphosphate receptor and the endoplasmic-reticulum Ca2+ pumps. Cell Calcium 1999, 26, 9–13. [Google Scholar]
- Hirata, Y; Uemura, D. Halichondrins - antitumor polyether macrolides from a marine sponge. Pure Appl Chem 1986, 58, 701–710. [Google Scholar]
- Bai, RL; Paull, KD; Herald, CL; Malspeis, L; Pettit, GR; Hamel, E. Halichondrin B and homohalichondrin B, marine natural products binding in the vinca domain of tubulin: Discovery of tubulin-based mechanism of action by analysis of differential cytotoxicity data. J Biol Chem 1991, 266, 15882–15889. [Google Scholar]
- Bai, R; Cichacz, ZA; Herald, CL; Pettit, GR; Hamel, E. Spongistatin 1, a highly cytotoxic, sponge-derived, marine natural product that inhibits mitosis, microtubule assembly, and the binding of vinblastine to tubulin. Mol Pharmacol 1993, 44, 757–766. [Google Scholar]
- Koiso, Y; Morita, K; Kobayashi, M; Wang, W; Ohyabu, N; Iwasaki, S. Effects of arenastatin A and its synthetic analogs on microtubule assembly. Chemico-Biol Interact 1996, 102, 183–191. [Google Scholar]
- Ter Haar, E; Kowalski, RJ; Hamel, E; Lin, CM; Longley, RE; Gunasekera, SP; Rosenkranz, HS; Day, BW. Discodermolide, a cytotoxic marine agent that stabilizes microtubules more potently than taxol. Biochemistry 1996, 35, 243–250. [Google Scholar]
- Anderson, HJ; Coleman, JE; Andersen, RJ; Roberge, M. Cytotoxic peptides hemiasterlin, hemiasterlin A and hemiasterlin B induce mitotic arrest and abnormal spindle formation. Cancer Chemother Pharmacol 1997, 39, 223–226. [Google Scholar]
- Mooberry, SL; Tien, G; Hernandez, AH; Plubrukarn, A; Davidson, BS. Laulimalide and isolaulimalide, new paclitaxel-like microtubule-stabilizing agents. Cancer Res 1999, 59, 653–660. [Google Scholar]
- Hood, KA; West, LM; Rouwé, B; Northocote, PT; Berridge, MV; Wakefield, SJ; Miller, JH. Peloruside A, a novel antimitotic agent with paclitaxel-like microtabule-stabilizing activity. Cancer Res 2002, 62, 3356–3360. [Google Scholar]
- Isbrucker, RA; Cummins, J; Pomponi, SA; Longley, RE; Wright, AE. Tubulin polymerizing activity of dictyostatin 1, a polyketide of marine sponge origin. Biochem Pharmacol 2003, 66, 75–82. [Google Scholar]
- Kashman, Y; Groweiss, A; Shmueli, U. Latruncutin, a new 2-thiazolidinone macrolide from the marine sponge Latrunculia magnifica. Tetrahedron Lett 1980, 21, 3629–3632. [Google Scholar]
- Coue, M; Brenner, SL; Spector, I; Korn, ED. Inhibition of actin polymerization by latrunculin A. FEBS Lett 1987, 213, 316–318. [Google Scholar]
- Fusetani, N; Yasumuro, K; Matsunaga, S; Hashimoto, K. Mycalolides A–C, hybrid macrolides of ulapualides and halichondramide, from a sponge of the genus Mycale. Tetrahedron Lett 1989, 30, 2809–2812. [Google Scholar]
- Saito, S; Watabe, S; Ozaki, H; Fusetani, N; Karaki, H. Mycalolide, a novel actin depolymerizing agent. J Biol Chem 1994, 269, 29710–29714. [Google Scholar]
- Bubb, MR; Spector, I; Bershadsky, AD; Korn, ED. Swinholide A is a microfilament disrupting marine toxin that stabilizes actin dimers and severs actin filaments. J Biol Chem 1995, 270, 3463–3466. [Google Scholar]
- Anderluh, G; Macek, P. Cytolytic peptide and protein toxins from sea anemones (Anthozoa: Actiniaria). Toxicon 2002, 40, 111–124. [Google Scholar]
- Crnigoj Kristan, K; Viero, G; Dalla Serra, M; Maček, P; Anderluh, G. Molecular mechanism of pore formation by actinoporins. Toxicon 2009. [Google Scholar]
- Amador, ML; Jimeno, J; Paz-Ares, L; Cortes-Funes, H; Hidalgo, M. Progress in the development and acquisition of anticancer agents from marine sources. Ann Oncol 2003, 14, 1607–1615. [Google Scholar]
- D’Incalci, M; Simone, M; Tavecchio, M; Damia, G; Garbi, A; Erba, E. New drugs from the sea. J Chemother 2004, 16, 86–89. [Google Scholar]
- Russo, GL; Ciarcia, G; Presidente, E; Siciliano, RA; Tosti, E. Cytotoxic and apoptogenic activity of a methanolic extract from the marine invertebrate Ciona intestinalis on malignant cell lines. Med Chem 2008, 4, 106–109. [Google Scholar]



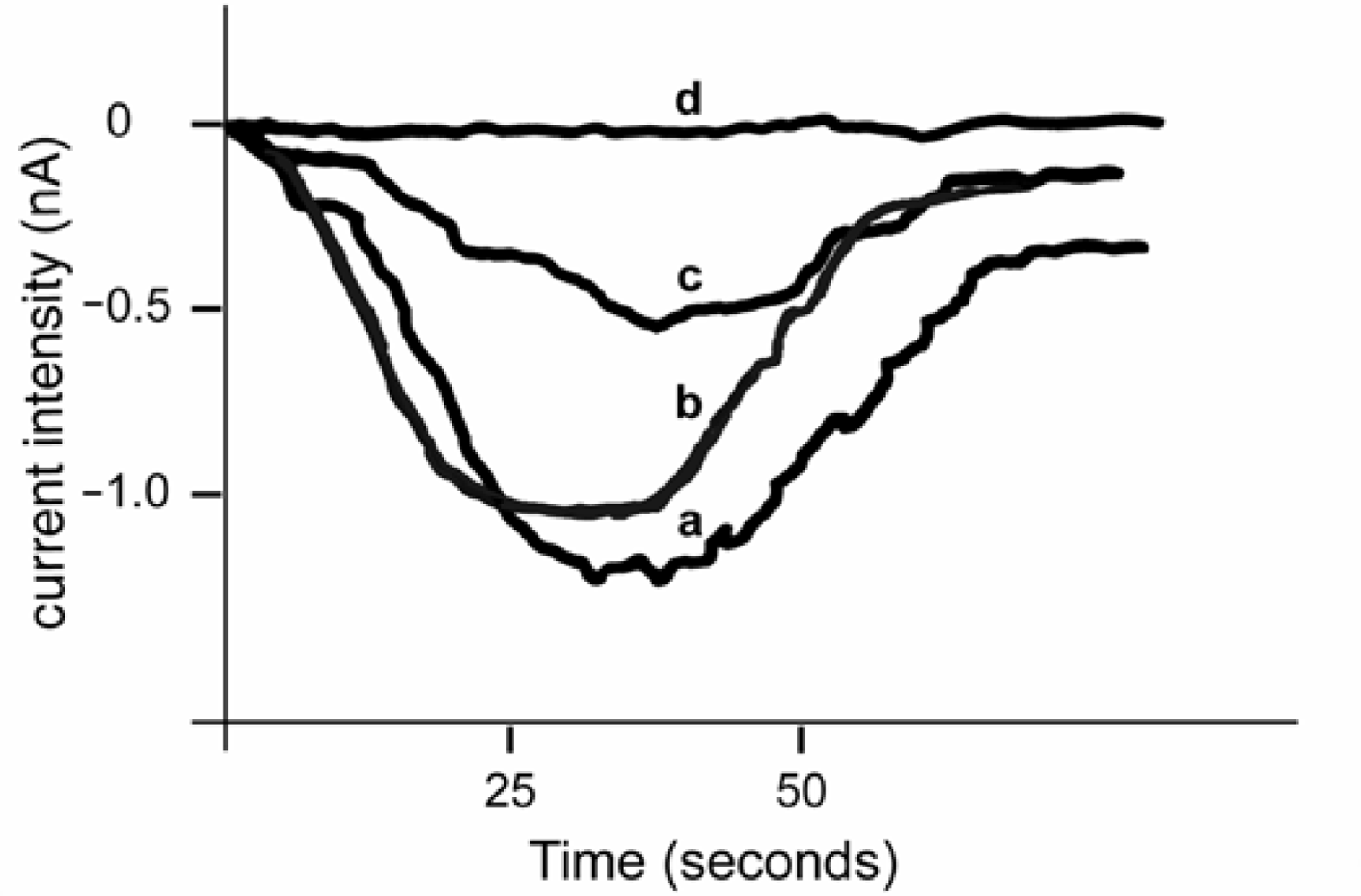{kind=link}
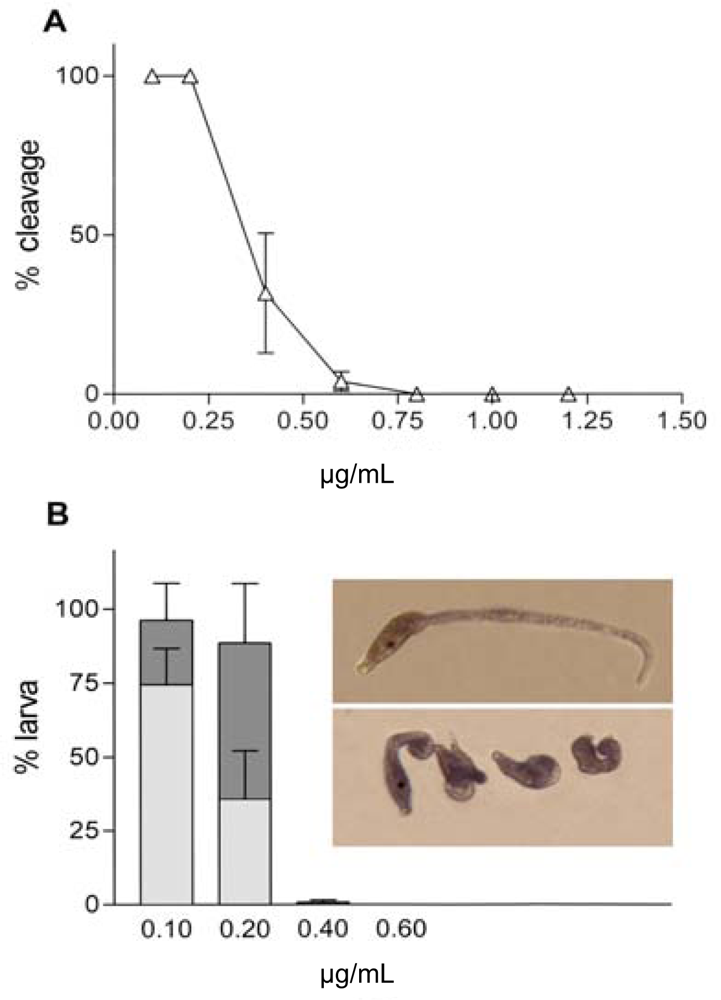{kind=link}
| Drug | Source | Target - Impact | References |
|---|---|---|---|
| Tetrodotoxin | Bacteria including Vibrio sp. | - Voltage-gated sodium channels; - Inhibition of fertilisation current in C. intestinalis; teratogenic effect | [14,96,200] |
| Maitotoxin | Dinoflagellate Gambierdiscus toxicus | - Cationic channel - Mouse acrosome reaction; inhibition of sea urchin egg fertilisation | [101,103] |
| Okadaic acid | Dinoflagellates Prorocentrum lima and Dinophysis spp. | - Protein phosphatases; - Inhibition of phosphorylation based processes of cellular signaling | [104,123] |
| Brevetoxin | Dinoflagellate Karenia brevis | - Voltage-gated sodium channels; - Teratogenic effect | [124] |
| 2-trans,4-trans Decadienal | Diatoms | - Fertilisation current; - Inhibition of embryonic development and fertilisation in broadcast spawning marine invertebrates | [140,142] |
| Domoic acid | Alga Chondria armata Diatom Pseudonitzschia | - Ionotropic glutamate receptors; - Neurotoxicity; reduction of larval growth and survival in P. maximus ; reproductive failure in California sea lions. | [143,152] |
| Caulerpenyne | Alga Caulerpa taxifolia | - Microtubules; - Inhibition of first cell division | [153] |
| Stypoldione | Alga Stypopodium zonale | - Sulfhydryl groups of proteins - Inhibition of cytokinesis in sea urchin embryos | [154] |
| Sulfonoglycolipid S-ACT-1 | Alga Gelidiella acerosa | - Sperm - Stimulation of sperm motility | [155] |
| Crude extract | Alga Gracilaria corticata, | - Increase of pre-implantation loss in femal rats | [156] |
| Crude extract | Alga Gelidiella acerosa | - Increase of post-implantation loss in female rats | [156] |
| Jasplakinolide | Sponge Jaspis johnstoni | - Actin; - Arrest of in vitro maturation | [161,162] |
| Theonellapeptolide Ie | Sponge Petrosia | - Cortical F-actin distribution; - Abnormal maturation | [158] |
| Strongylophorine | Sponge Strongylophora strongylata | -Actin; -Inhibition of the maturation of starfish oocytes | [160] |
| Latrunculin | Sponge Latrunculia magnifica | - Microfilament; - Inhibition of microfilament-mediated processes during fertilisation, cleavage and early development in sea urchins and mice. | [163] |
| (−)-10-epi-Axiso-nitrile-3 | Sponge Geodia Exigua | - Phosphocreatine shuttle; - Sea urchin and starfish sperm immobilization | [165] |
| Theonellapeptolide IId | Sponge Theonella swinhoei | - Inhibition of fertilisation of the sea urchin | [166] |
| Jaspisin | Sponge Jaspis sp. | - Exoplasmic membrane fusion; - Block fusion between sperm acrosomal process and egg plasma membrane; block of embryo hatching | [167–169] |
| Callyspongins A and B | Sponge Callyspongia truncata | - Inhibition of sperm-egg fusion | [170] |
| Halenaquinol sulfate | Sponge Xestospongia sapra | - Inhibition of sperm-egg fusion | [171,172] |
| Calyculin A | Sponge Discodermia calyx | - Protein phosphatases - Modulation of phosphorylation based processes of cellular signalling | [174] |
| Bonellin | Echiura Bonellia viridis | - Membrane proteins; - Formation of peroxydase; cytolysis of unfertilised eggs, inhibition of cleavage and of larval motility | [178] |
| Bivittoside D | Sea cucumber Bohadschia vitiensis | - Sperm; - Membrane permeabilization; spermicide activity in human | [179] |
| Methoxyconidiol | Ascidian Aplidium aff. densum | - Microtubules; - Inhibition of cleavage | [180,181] |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Silvestre, F.; Tosti, E. Impact of Marine Drugs on Animal Reproductive Processes. Mar. Drugs 2009, 7, 539-564. https://doi.org/10.3390/md7040539
Silvestre F, Tosti E. Impact of Marine Drugs on Animal Reproductive Processes. Marine Drugs. 2009; 7(4):539-564. https://doi.org/10.3390/md7040539
Chicago/Turabian StyleSilvestre, Francesco, and Elisabetta Tosti. 2009. "Impact of Marine Drugs on Animal Reproductive Processes" Marine Drugs 7, no. 4: 539-564. https://doi.org/10.3390/md7040539
APA StyleSilvestre, F., & Tosti, E. (2009). Impact of Marine Drugs on Animal Reproductive Processes. Marine Drugs, 7(4), 539-564. https://doi.org/10.3390/md7040539