Recovery of Proteolytic and Collagenolytic Activities from Viscera By-products of Rayfish (Raja clavata)



Abstract
:1. Introduction
2. Results and Discussion
2.1. Study of proteolytic activities from different gastrointestinal sections
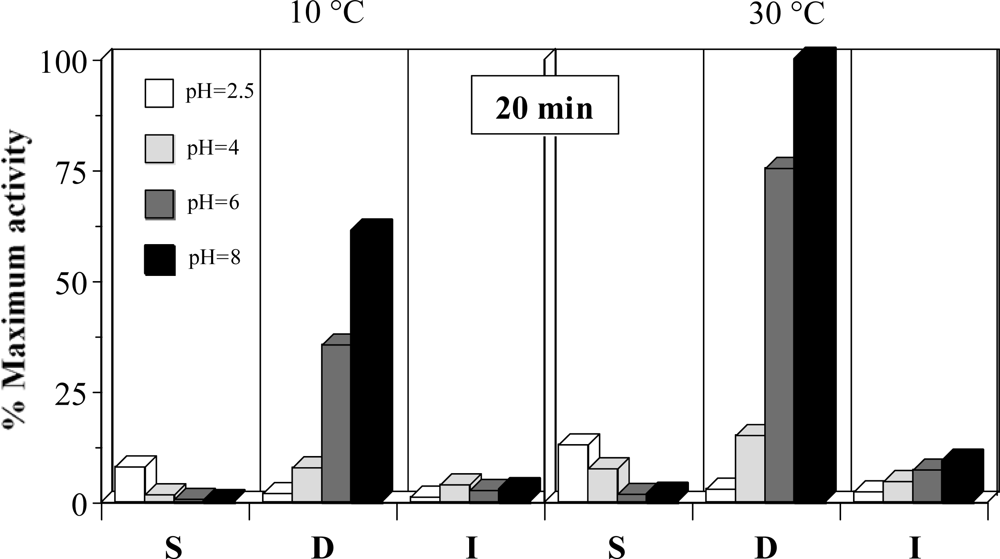
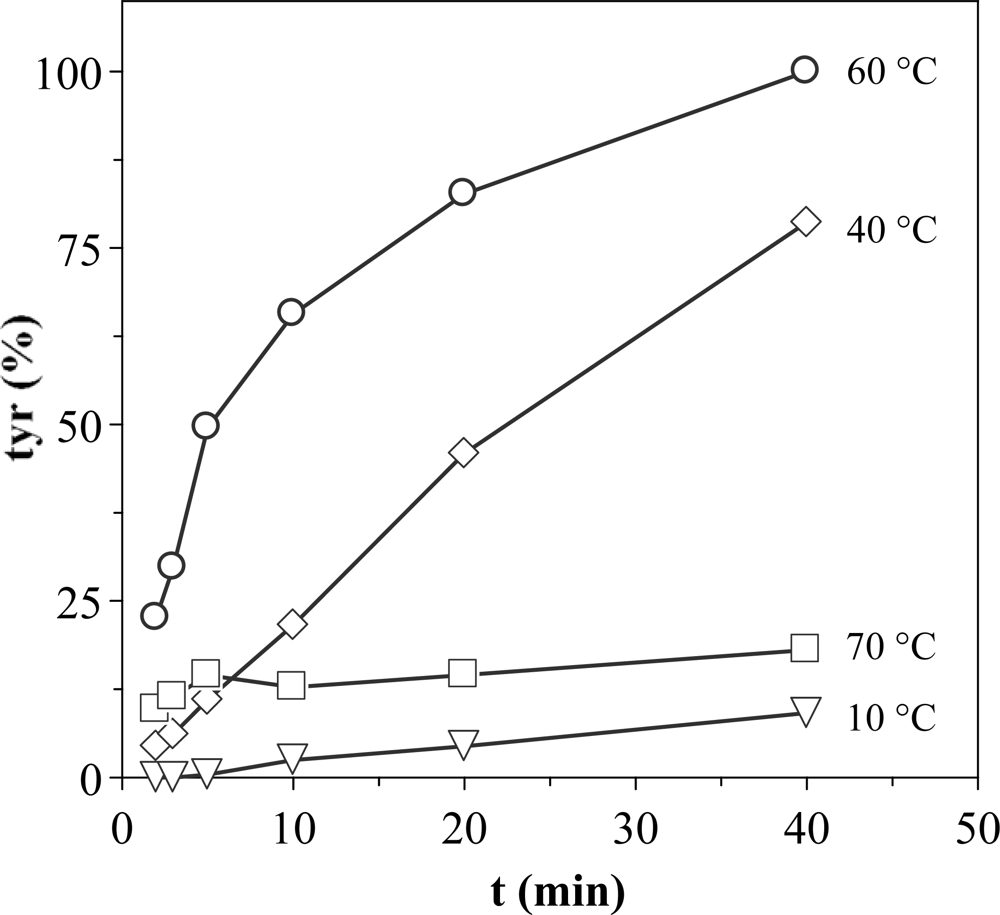
2.2. Optimization of the experimental conditions for proteolytic activity quantification
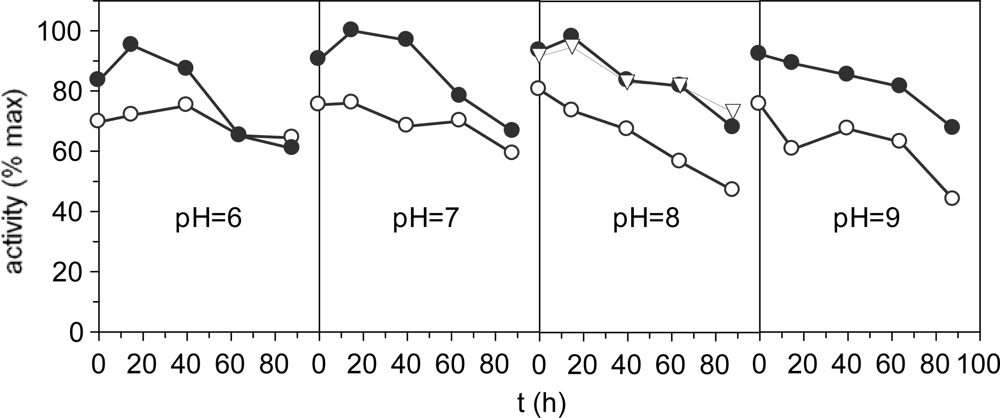
2.3. Effect of the pH on extraction conditions and stability
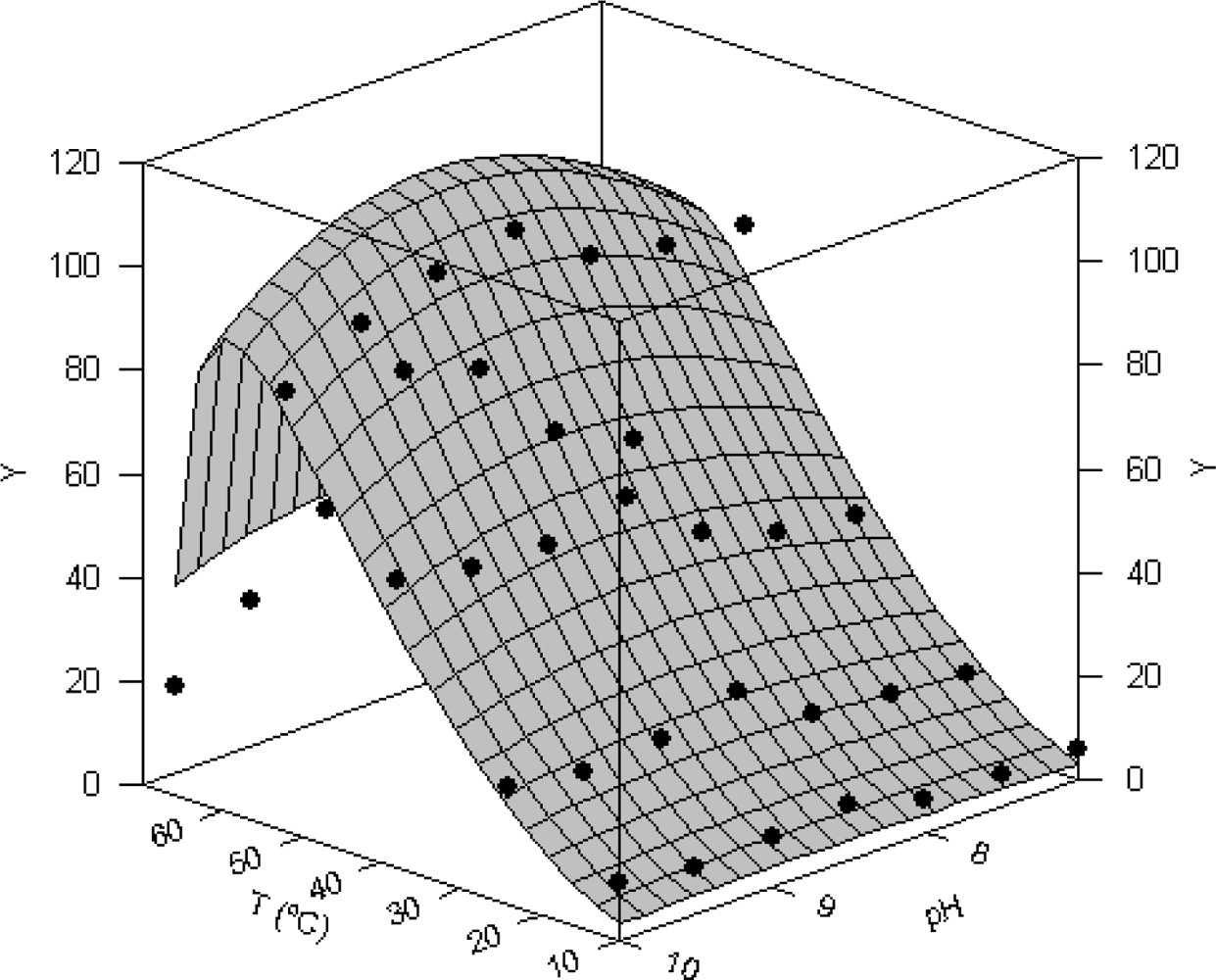
2.4. Joint effect of pH and temperature on proteolytic activity
2.5. Partial purification of Procolax
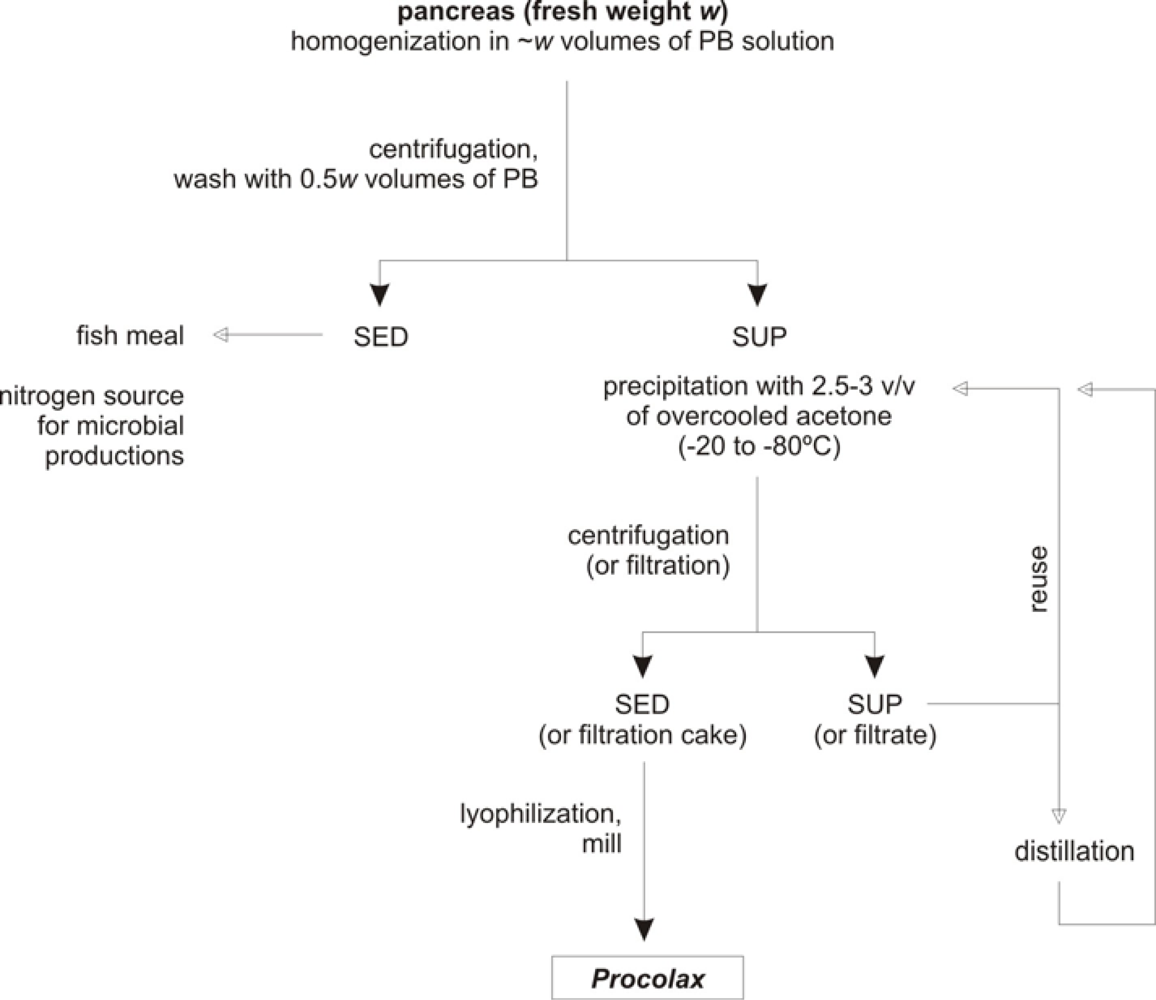
2.6. Protocol for Procolax recovery
- Initially, a –w– weight (in kg) of fresh material (pancreas and duodenum) is homogenized in 1 × w volumes (in litres) of phosphate buffer (0.05 M, pH 7.5) with Triton X-100 (0.2%) and KCl (0.08M). Subsequently, the homogenizate is maintained for 1 h with a continuous and soft agitation in order to avoid enzyme deactivation.
- The homogenate is centrifuged (4,000 g for 15 min) to obtain a supernatant with the enzymatic activity and sediment. An additional wash of this sediment with 0.5 × w volume of phosphate buffer and subsequently centrifugation in the same conditions produces an increment of 20% in the enzyme recovered. The whole of sediments could be used as source of nitrogen for bio-silage and hyaluronic acid production [32,36] or substrate for fish meal.
- All the supernatants obtained in the previous step are precipitated by means of overcooled acetone addition. Among 2.5 and 3 volumes of acetone are required and slow addition and continuous and soft agitation for 30 min are also necessary.
- The floccules formed when acetone is adding are filtrated via Whatman Nº1 filter. This process of separation is more efficient if floccules are spontaneously sedimented for 30 min and the liquid suspension is drained with a pump.
- The acetone from the filtration cake can be evaporated applying vacuum. Subsequently, the cake is lyophilized and finally milled. The powder obtained is approximately equivalent to the 10% of wet weight from the initial material.
- The hydro-acetone solution from the filtration step could be used again for a new precipitation in combination with pure acetone until 4–6 reuses. From this moment acetone should be rectified by distillation.
3. Experimental Section
3.1. Waste material from rayfish and reagents
3.2. Determination of enzymatic activities
3.2.1. Proteases
3.2.2. Collagenases
3.2.3. Proteolytic activities from different intestinal sections
3.3. Effect of the pH on extraction conditions and stability
3.4. Joint effect of pH and temperature on protease activity
3.5. Partial purification of Procolax
3.6. Analytical methods
3.7. Mathematical equation and numerical methods
4. Conclusions
Acknowledgments
References and Notes
- Godfrey, T; West, S. Industrial Enzymology, 2nd ed; Macmillan: New York, NY, USA, 1996; p. 3. [Google Scholar]
- Gupta, R; Beg, QK; Khan, S; Chauhan, B. An overview on fermentation, downstream processing and properties of microbial alkaline proteases. Appl Microbiol Biotechnol 2002, 60, 381–395. [Google Scholar]
- Kumar, CG; Takagi, H. Microbial alkaline proteases from a bioindustrial viewpoint. Biotecnol Adv 1999, 17, 561–594. [Google Scholar]
- Kristinsson, HG; Rasco, BA. Fish protein hydrolysates: Production, biochemical, and functional properties. Crit Rev Food Sci Nutr 2000, 40, 43–81. [Google Scholar]
- Gildberg, A. Recovery of proteinases and protein hydrolysates from fish viscera. Bior Technol 1992, 39, 271–276. [Google Scholar]
- Gildberg, A. Utilisation of male Arctic capelin and Atlantic cod intestines for fish sauce production - Evaluation of fermentation conditions. Bior Technol 2001, 76, 119–123. [Google Scholar]
- Vázquez, JA; Murado, MA. Enzymatic hydrolysates from food wastewaters as a source of peptones for lactic acid bacteria productions. Enzyme Microb Technol 2008, 43, 66–72. [Google Scholar]
- Tao, H; Zhao, MM; Wang, JS; Yang, B; Cui, C; Zhao, QZ; Wu, H. Activity distribution of digestive proteases from Nemipterus virgatus and their responses to pH value and temperature. J Food Proc Eng 2008, 31, 21–33. [Google Scholar]
- Choudhury, GS. Food products made from protease enzyme containing fish, methods of making same, and methods to inactivate protease enzyme in fish. US Patent Nº 5,676,986 1997. [Google Scholar]
- Sovik, SL; Rustad, T. Proteolytic activity in byproducts from cod species caught at three different fishing grounds. J Agric Food Chem 2005, 53, 452–458. [Google Scholar]
- Tran, LH; Nagano, H. Isolation and characteristics of Bacillus subtilis CN2 and its collagenase production. J Food Sci 2002, 67, 1184–1187. [Google Scholar]
- Banerjee, UC; Agnihotri, R; Bhattacharyya, BC. Purification of alkaline protease of Rhizopus oryzae by foam fractionation. Bioproc Eng 1993, 9, 245–248. [Google Scholar]
- Johnvesly, B; Naik, GR. Studies on production of thermostable alkaline protease from thermophilic and alkaliphilic Bacillus sp. JB-99 in a chemically defined medium. Proc Biochem 2001, 37, 139–144. [Google Scholar]
- De Azevedo, LAI; Freire, DMG; Soares, RMA; Leite, SGF; Coelho, RRR. Production and partial characterization of thermophilic proteases from Streptomyces sp. isolated from Brazilian cerrado soil. Enzyme Microb Technol 2004, 34, 354–358. [Google Scholar]
- Ustáriz, FJ; Laca, A; García, LA; Díaz, M. Fermentation of individual proteins for protease production by Serratia marcescens. Biochem Eng J 2004, 19, 147–153. [Google Scholar]
- Dutta, JR; Dutta, PK; Banerjee, R. Modeling and optimization of protease production by a newly isolated Pseudomonas sp. using a genetic algorithm. Proc Biochem 2005, 40, 879–884. [Google Scholar]
- Tremacoldi, C; Carmona, EC. Production of extracellular alkaline proteases by Aspergillus clavatus. World J Microbiol Biotechnol 2005, 21, 169–172. [Google Scholar]
- Agrawal, D; Patidar, P; Banerjee, T; Patil, S. Alkaline protease production by a soil isolate of Beauveria felina under SSF condition: Parameter optimization and application to soy protein hydrolysis. Proc Biochem 2005, 40, 1131–1136. [Google Scholar]
- Vázquez, JA; Docasal, SF; Mirón, J; González, MP; Murado, MA. Proteases production by two Vibrio species on residuals marine media. J Ind Microbiol Biotechnol 2006, 33, 661–668. [Google Scholar]
- Kishimura, H; Hayashi, K; Miyashita, Y; Nonami, Y. Characteristics of trypsins from the viscera of true sardine (Sardinops melanostictus) and the pyloric ceca of arabesque greenling (Pleuroprammus azonus). Food Chem 2006, 97, 65–70. [Google Scholar]
- Khaled, HB; Bougatef, A; Balti, R; Triki-Ellouz, Y; Souissi, N; Nasri, M. Isolation and characterisation of trypsin from sardinelle (Sardinella aurita) viscera. J Sci Food Agric 2008, 88, 2654–2662. [Google Scholar]
- Espósito, TS; Amaral, IPG; Buarque, DS; Oliveira, GB; Carvalho, LB, Jr; Bezerra, RS. Fish processing waste as a source of alkaline proteases for laundry detergent. Food Chem 2009, 112, 125–130. [Google Scholar]
- Castillo-Yañez, FJ; Pachecho-Aguilar, R; Lugo-Sánchez, ME; García-Sánchez, G; Quintero-Reyes, IE. Biochemical characterization of an isoform of chymotrypsin from the viscera of Monterey sardine (Sardinops sagax caerulea), and comparison with bovine chymotrypsin. Food Chem 2009, 112, 634–639. [Google Scholar]
- Klomklao, S; Kishimura, H; Benjakul, S; Simpson, BK. Autolysis and biochemical properties of endogenous proteinases in Japanese sandfish (Arctoscopus japonicus). Int J Food Sci Technol 2009, 44, 1344–1350. [Google Scholar]
- Simpson, BK; Haard, NF. Cold-adapted enzymes from fish. In Food Biotechnology; Knorr, D, Ed.; Marcel Dekker: New York, NY, USA, 1987; pp. 495–528. [Google Scholar]
- Asgeirsson, B; Fox, JW; Bjarnason, JB. Purification and characterization of trypsin from the poikilotherm Gadus morhua. Eur J Biochem 1989, 180, 85–94. [Google Scholar]
- Kristjansson, MM. Purification and characterization of trypsin from the pyloric caeca of rainbow trout (Oncorhynchus mykiss). J Agric Food Chem 1991, 39, 1738–1742. [Google Scholar]
- Simpson, BK; Haard, NF. Marine Enzymes. In Encyclopedia of Food Science and Technology, 2nd ed; Francis, FJ, Ed.; Wiley: New York, NY, USA, 1999; Volume 3, pp. 495–528. [Google Scholar]
- Blanco, M; Sotelo, CG; Chapela, MJ; Pérez-Martín, RI. Towards sustainable and efficient use of fishery resources: Present and future trends. Trends Food Sci Technol 2007, 18, 29–36. [Google Scholar]
- Vázquez, JA; González, MP; Murado, MA. A new marine medium. Use of different fish peptones and comparative study of selected species of marine bacteria. Enzyme Microb Technol 2004, 35, 385–392. [Google Scholar]
- Vázquez, JA; González, MP; Murado, MA. Preliminary tests on nisin and pediocin production using waste protein sources. Factorial and kinetic studies. Bior Technol 2006, 97, 605–613. [Google Scholar]
- Vázquez, JA; Docasal, SF; Prieto, MA; González, MP; Murado, MA. Growth and metabolic features of lactic acid bacteria in media with hydrolysed fish viscera. An approach to bio-silage of fishing by-products. Bior Technol 2008, 99, 6246–6257. [Google Scholar]
- Castillo-Yánez, FJ; Pacheco-Aguilar, R; García-Carreño, FL; Navarrete-Del Toro, MA. Characterization of acidic proteolytic enzymes from Monterey sardine (Sardinops sagax caerulea) viscera. Food Chem 2004, 85, 343–350. [Google Scholar]
- Ben Khaled, H; Bougatef, A; Balti, R; Triki-Ellouz, Y; Souissi, N; Nasri, M. Isolation and characterisation of trypsin from sardinelle (Sardinella aurita) viscera. J Sci Food Agric 2008, 88, 2654–2662. [Google Scholar]
- Michail, M; Vasiliadou, M; Zotos, A. Partial purification and comparison of precipitation techniques of proteolytic enzymes from trout (Salmo gairdnerii) heads. Food Chem 2006, 97, 50–55. [Google Scholar]
- Vázquez, JA; Montemayor, MI; Fraguas, J; Murado, MA. High production of hyaluronic and lactic acids by Streptococcus zooepidemicus in fed-batch culture using commercial and marine peptones from fishing by-products. Biochem Eng J 2009, 44, 125–130. [Google Scholar]
- Kunitz, M. Crystalline soybean trypsin inhibitor 2: General properties. J Gen Physiol 1947, 30, 291–310. [Google Scholar]
- Barker, TW; Worgan, JT. The utilisation of palm oil processing effluents as substrates for microbial protein production by the fungus. Aspergillus oryzae Eur J Appl Microbiol Biotechnol 1981, 11, 234–240. [Google Scholar]
- van Wart, HE; Steinbrink, DR. A continuous spectrophotometric assay for Clostridium histolyticum collagenase. Anal Biochem 1981, 113, 356–365. [Google Scholar]
- Lowry, OH; Rosebrough, NJ; Farr, AL; Randall, RJ. Protein measurement with the folin phenol reagent. J Biol Chem 1951, 270, 27299–27304. [Google Scholar]
- Rosso, L; Lobry, JR; Flandrois, JP. An unexpected correlation between cardinal temperatures of microbial growth highlighted by a new model. J Theor Biol 1993, 162, 447–463. [Google Scholar]
- Rosso, L; Lobry, JR; Bajard, S; Flandrois, JP. Convenient model to describe the combined effect of temperature and pH on microbial growth. Appl Env Microbiol 1995, 61, 610–616. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (NH4)2SO4 saturation (%) | A: Total Protein (g/L) | B: Total Activity (EU/mL) | C: Specific activity (EU/g protein) | D: Recovery (%) | E: Purification factor | F: C × D |
|---|---|---|---|---|---|---|
| 20 | 2.78 | 0.218 | 78.3 | 21.0 | 1.34 | 1647 |
| 40 | 3.59 | 0.623 | 173.6 | 72.4 | 2.97 | 12570 |
| 60 | 4.84 | 0.804 | 166.0 | 86.1 | 2.84 | 14299 |
| 80 | 4.49 | 0.621 | 138.2 | 87.0 | 2.36 | 12021 |
| % from the total extract | Ratio: first extract/wash | |||||
|---|---|---|---|---|---|---|
| Protein | Proteases | Collagenases | Protein | Proteases | Collagenases | |
| Extract | 78.1 | 79.6 | 83.8 | 3.6 | 3.9 | 5.2 |
| Wash | 21.9 | 20.4 | 16.2 | 1 | 1 | 1 |
| Proteases | Collagenases | |
|---|---|---|
| Pancreas | 62.4 ± 5.2 | 2590 ± 198 |
| Duodenum without pancreas | 6.5 ± 0.5 | 660 ± 72 |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Murado, M.A.; González, M.d.P.; Vázquez, J.A. Recovery of Proteolytic and Collagenolytic Activities from Viscera By-products of Rayfish (Raja clavata). Mar. Drugs 2009, 7, 803-815. https://doi.org/10.3390/md7040803
Murado MA, González MdP, Vázquez JA. Recovery of Proteolytic and Collagenolytic Activities from Viscera By-products of Rayfish (Raja clavata). Marine Drugs. 2009; 7(4):803-815. https://doi.org/10.3390/md7040803
Chicago/Turabian StyleMurado, Miguel Anxo, María del Pilar González, and José Antonio Vázquez. 2009. "Recovery of Proteolytic and Collagenolytic Activities from Viscera By-products of Rayfish (Raja clavata)" Marine Drugs 7, no. 4: 803-815. https://doi.org/10.3390/md7040803
APA StyleMurado, M. A., González, M. d. P., & Vázquez, J. A. (2009). Recovery of Proteolytic and Collagenolytic Activities from Viscera By-products of Rayfish (Raja clavata). Marine Drugs, 7(4), 803-815. https://doi.org/10.3390/md7040803