Isolation, Phylogenetic Analysis and Anti-infective Activity Screening of Marine Sponge-Associated Actinomycetes

Abstract
:1. Introduction
2. Results and Discussion
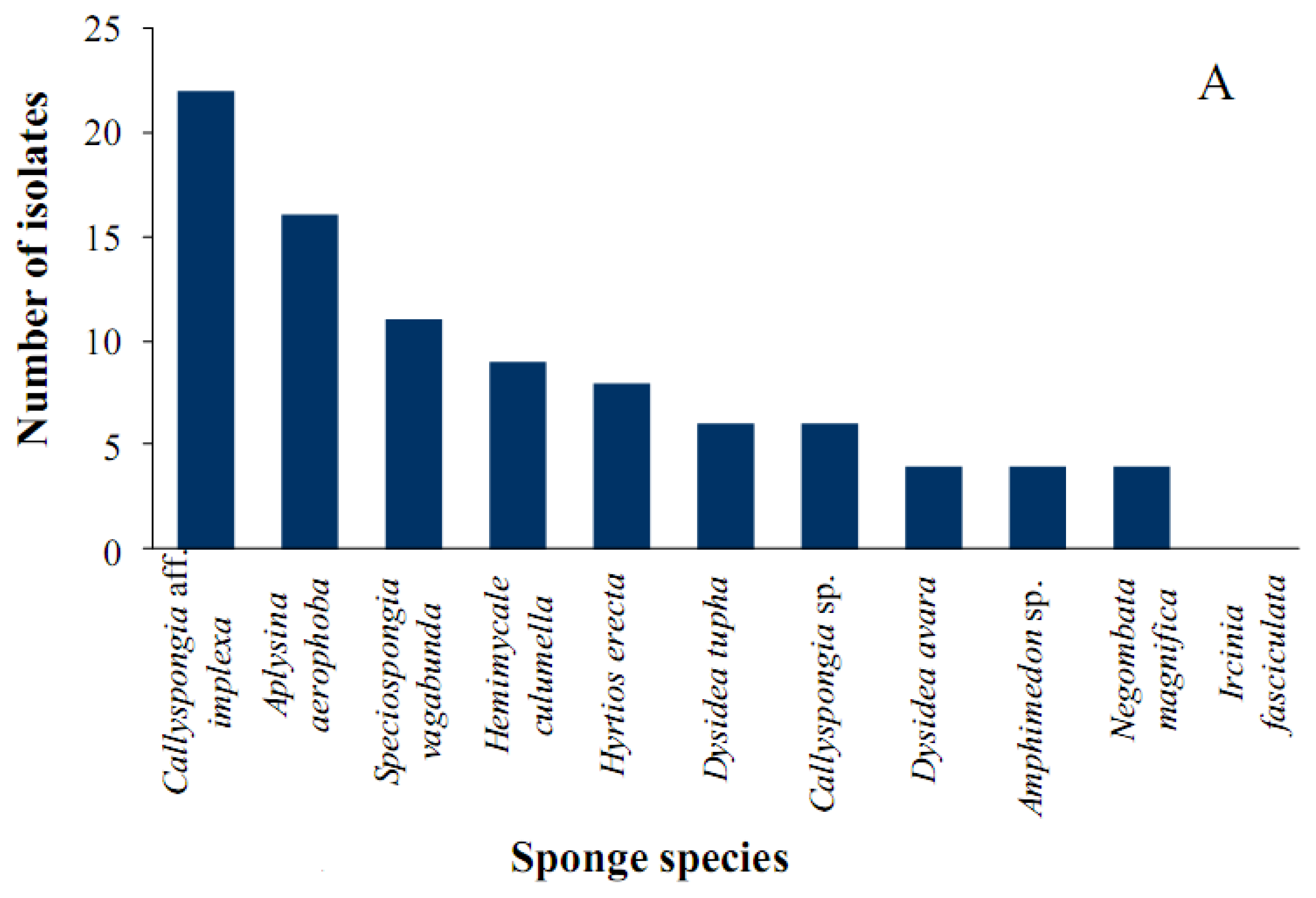
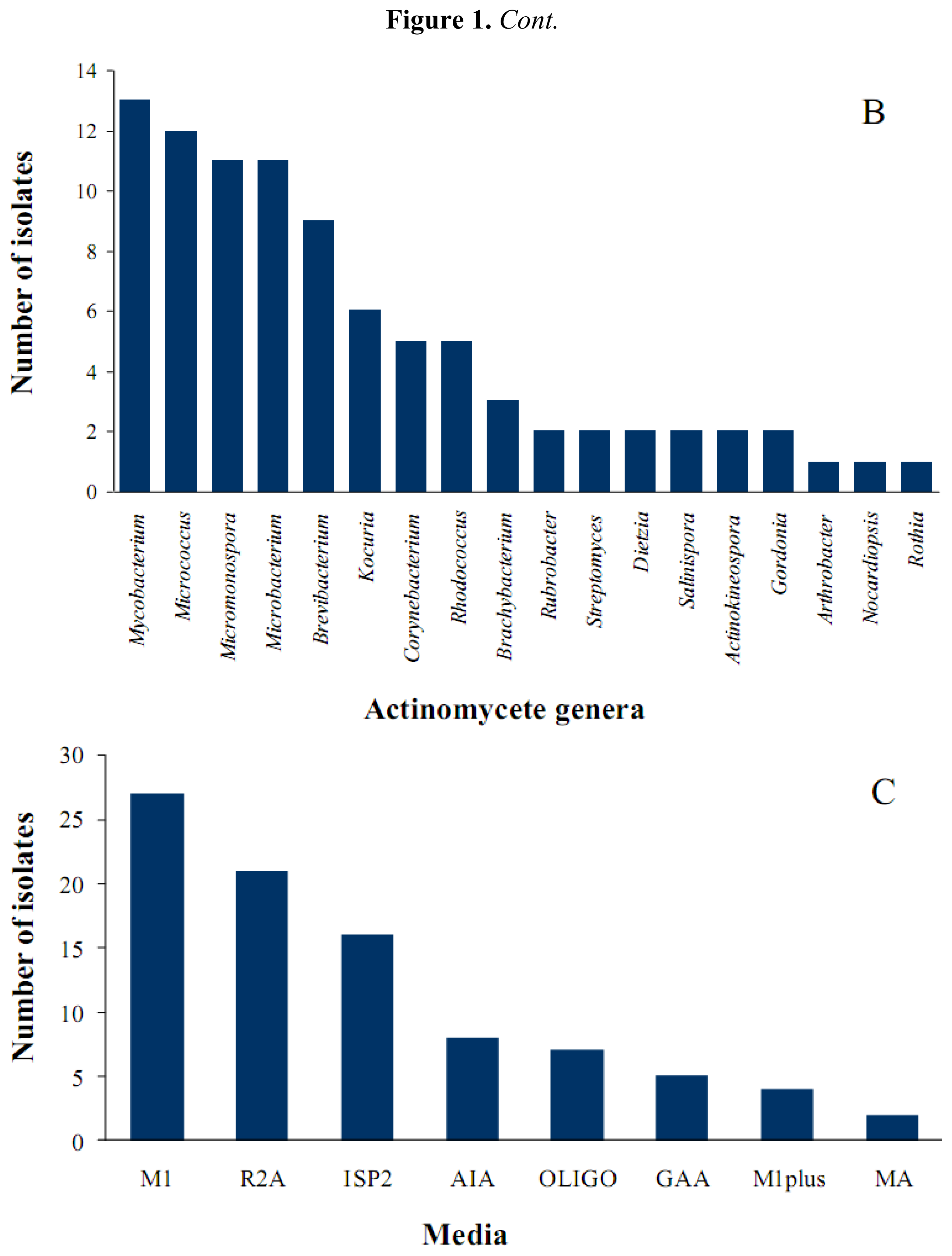
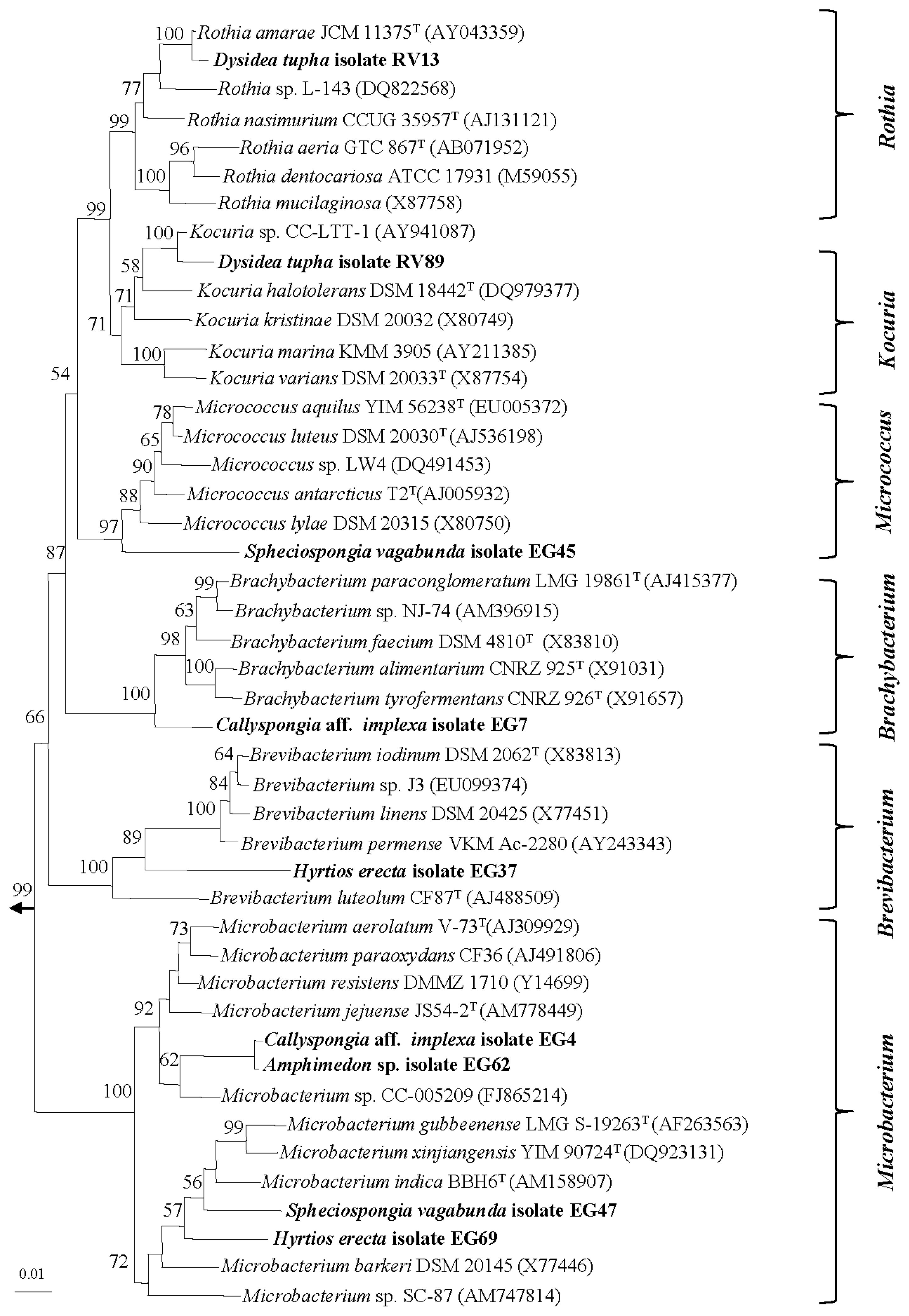
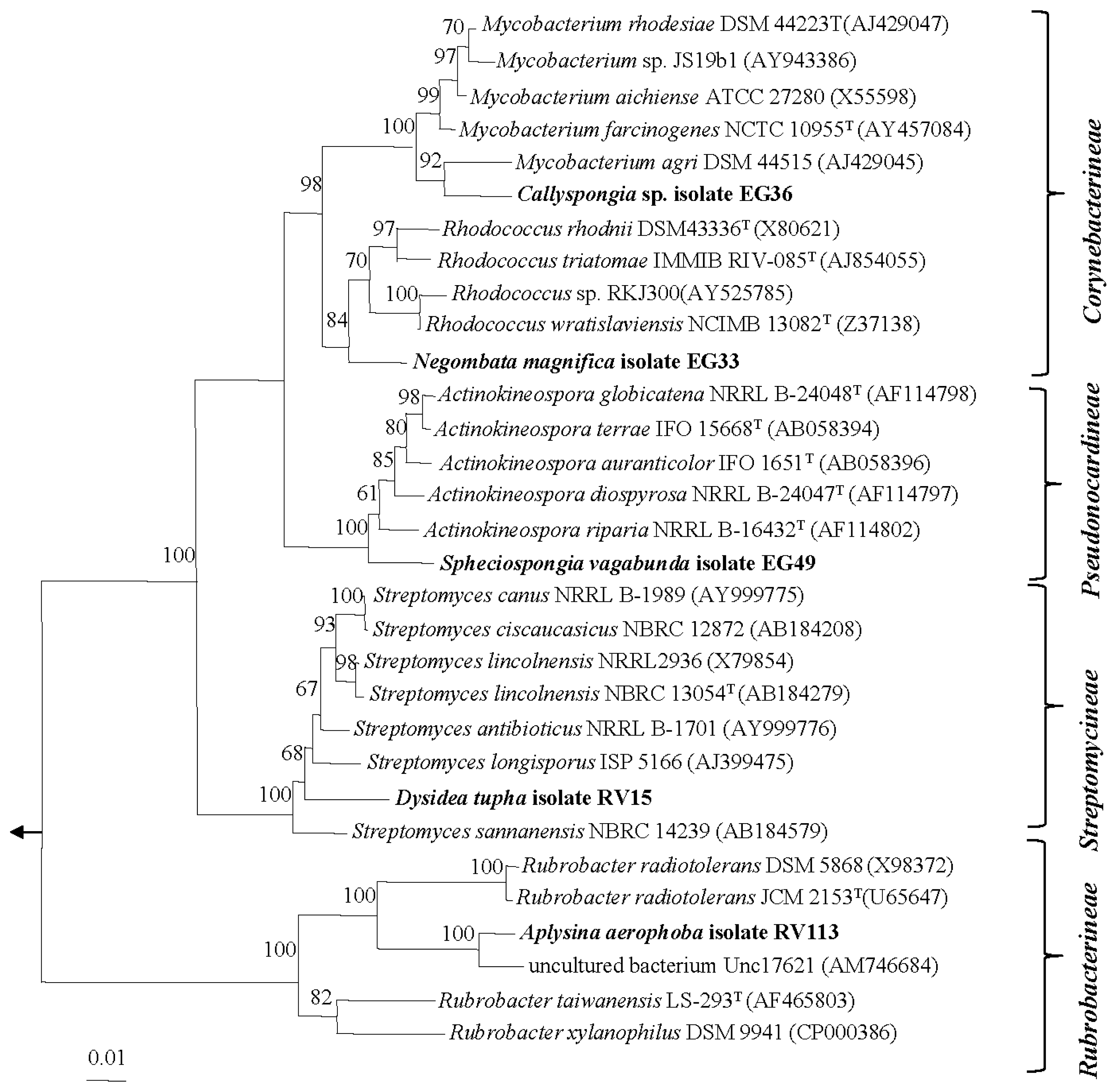
2.1. Diversity of Sponge-Associated Actinomycetes
2.2. Bioactivity Testing
3. Experimental
3.1. Sponge Collection
3.2. Actinomycete Isolation
3.3. Molecular Identification and Phylogenetic Analysis
3.4. Extract Preparation and Anti-infective Activity Screening
Acknowledgments
- Samples Availability: Available from the authors.
References
- Pinner, R; Teutsch, S; Simonsen, L; Klug, L; Graber, J; Clarke, M; Berkelman, R. Trends in infectious diseases mortality in the United States. J Am Med Assoc 1996, 275, 189–193. [Google Scholar]
- Woolhouse, MEJ. Epidemiology: Emerging diseases go global. Nature 2008, 451, 898–899. [Google Scholar]
- Morens, DM; Folkers, GK; Fauci, AS. The challenge of emerging and re-emerging infectious diseases. Nature 2004, 430, 242–249. [Google Scholar]
- Daszak, P; Cunningham, AA; Hyatt, AD. Emerging infectious diseases of wildlife-threats to biodiversity and human health. Science 2000, 287, 443–449. [Google Scholar]
- Jones, KE; Patel, NG; Levy, MA; Storeygard, A; Balk, D; Gittleman, JL; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar]
- Blunt, JW; Copp, BR; Hu, VP; Munro, MH; Northcote, PT; Prinsep, MR. Marine natural products. Nat Prod Rep 2007, 24, 31–86. [Google Scholar]
- Hentschel, U; Usher, KM; Taylor, MW. Marine sponges as microbial fermenters. FEMS Microbiol Ecol 2006, 55, 167–177. [Google Scholar]
- Taylor, MW; Radax, R; Steger, D; Wagner, M. Sponge-associated microorganisms: Evolution, ecology, and biotechnological potential. Microbiol Mol Biol Rev 2007, 71, 295–347. [Google Scholar]
- Webster, NS. Sponge disease: A global threat. Environ Microbiol 2007, 9, 1363–1375. [Google Scholar]
- Hentschel, U; Hopke, J; Horn, M; Friedrich, AB; Wagner, M; Hacker, J; Moore, BS. Molecular evidence for a uniform microbial community in sponges from different oceans. Appl Environ Microbiol 2002, 68, 4431–4440. [Google Scholar]
- Jiang, S; Li, X; Zhang, L; Sun, W; Dai, S; Xie, L; Liu, Y; Lee, K. Culturable actinobacteria isolated from marine sponge Iotrochota sp. Mar Biol 2008, 153, 945–952. [Google Scholar]
- Kim, TK; Garson, MJ; Fuerst, JA. Marine actinomycetes related to the ‘Salinospora’ group from the great barrier reef sponge Pseudoceratina clavata. Environ Microbiol 2005, 7, 509–518. [Google Scholar]
- Montalvo, NF; Mohamed, NM; Enticknap, JJ; Hill, RT. Novel actinobacteria from marine sponges. Anton Leeuwenhoek 2005, 87, 29–36. [Google Scholar]
- Webster, NS; Wilson, KJ; Blackall, LL; Hill, RT. Phylogenetic diversity of bacteria associated with the marine sponge Rhopaloeides odorabile. Appl Environ Microbiol 2001, 67, 434–444. [Google Scholar]
- Zhang, HT; Lee, YK; Zhang, W; Lee, HK. Culturable actinobacteria from the marine sponge Hymeniacidon perleve: Isolation and phylogenetic diversity by 16S rRNA Gene-RFLP analysis. Anton Leeuwenhoek 2006, 90, 159–169. [Google Scholar]
- Bredholdt, H; Galatenko, OA; Engelhardt, K; Fjærvik, E; Terekhova, LP; Zotchev, SB. Rare actinomycete bacteria from the shallow water sediments of the trodheim fjord, norway: Isolation, diversity and biological activity. Environ Microbiol 2007, 9, 2756–2764. [Google Scholar]
- Fenical, W; Jensen, PR. Developing a new resource for drug discovery: Marine actinomycete bacteria. Nat Chem Biol 2006, 2, 666–673. [Google Scholar]
- Grein, A; Meyers, SP. Growth characteristics and antibiotic production of actinomycetes isolated from littoral sediments and materials suspended in sea water. J Bacteriol 1958, 76, 457–463. [Google Scholar]
- Mincer, TJ; Jensen, PR; Kauffman, CA; Fenical, W. Widespread and persistent populations of a major new marine actinobacteria taxon in ocean sediments. Appl Environ Microbiol 2002, 68, 5005–5011. [Google Scholar]
- Panthom-Aree, W; Stach, JEM; Ward, AC; Horikoshi, K; Bull, AT; Goodfellow, M. Diversity of actinomycetes isolated from challenger deep sediment (10,898 m) from the Mariana Trench. Extremophiles 2006, 10, 181–189. [Google Scholar]
- Maldonado, LA; Fenical, W; Jensen, PR; Kauffman, CA; Mincer, TJ; Ward, AC; Bull, AT; Goodfellow, M. Salinispora arenicola gen. nov., sp. nov. and Salinispora tropica sp. nov., obligate marine actinomycetes belonging to the family Micromonosporaceae. Int J Syst Evol Microbiol 2005, 55, 1759–1766. [Google Scholar]
- Lam, KS. Discovery of novel metabolites from marine actinomycetes. Curr Opin Microbiol 2006, 9, 245–251. [Google Scholar]
- Maldonado, LA; Stach, JEM; Pathom-aree, W; Ward, AC; Bull, AT; Goodfellow, M. Diversity of cultivable actinobacteria in geographically widespread marine sediments. Anton Leeuwenhoek 2005, 87, 11–18. [Google Scholar]
- Kim, TK; Hewavitharana, AK; Shaw, PN; Fuerst, JA. Discovery of a new source of rifamycin antibiotics in marine sponge actinobacteria by phylogenetic prediction. Appl Environ Microbiol 2006, 72, 2118–2125. [Google Scholar]
- Zhang, HT; Zhang, W; Jin, Y; Jin, M; Yu, X. A comparative study on the phylogenetic diversity of culturable actinobacteria isolated from five marine sponge species. Anton Leeuwenhoek 2008, 93, 241–248. [Google Scholar]
- Webster, NS; Hill, RT. The culturable microbial community of the great barrier reef sponge Rhopaloeides odorabile. Appl Environ Microbiol 2001, 138, 843–851. [Google Scholar]
- Stackebrandt, E; Ebers, J. Taxonomic parameters revisited: Tarnished gold standards. Microbiol Today 2006, 33, 152–155. [Google Scholar]
- Radwan, M; Hanora, A; Zan, J; Mohamed, NM; Abo-Elmatty, DM; Abou-El-Ela, SH; Hill, RT. Bacterial community analyses of two Red Sea sponges. Mar Biotechnol 2009. [Google Scholar] [CrossRef]
- Bultel-Ponce, VC; Debitus, JP; Berge, GP; Guyot, M. Metabolites from the marine sponge-associated bacterium Micrococcus luteus. J Mar Biotech 1998, 6, 233–236. [Google Scholar]
- Lee, HK; Lee, DS; Lim, J; Kim, JS; Im, KS; Jung, JH. Topoisomerase I inhibitors from the Streptomyces sp. Strain KM86–9B isolated from a marine sponge. Arch Pharm Res 1998, 2, 729–733. [Google Scholar]
- Pimentel-Elardo, SM. Novel Anti-infective secondary metabolites and biosynthetic gene clusters from actinomycetes associated with marine sponges. Dissertation thesis, University of Würzburg, Würzburg, Germany, 2008. [Google Scholar]
- Oishi, H; Noto, T; Sasaki, H; Suzuki, K; Hayashi, T; Okazaki, H; Ando, K; Sawada, M. Thiolactomycin, a new antibiotic. 1. taxonomy of the prodrug organism, fermentation and biological properties. J Antibiot 1982, 35, 391–395. [Google Scholar]
- Loiseau, PM; Imbertie, L; Bories, C; Betbeder, D; De Miguel, V. Design and antileishmanial activity of amphotericin b-loaded stable ionic amphiphile biovector formulations. Antimicrob Agents Chemother 2002, 46(5), 1597–1601. [Google Scholar]
- Bull, AT; Stach, JE. Marine actinobacteria: New opportunities for natural product search and discovery. Trends Microbiol 2007, 15, 491–499. [Google Scholar]
- Deushi, T; Yamaguchi, T; Kamiya, K; Iwasaki, A; Mizoguchi, T; Nakayama, M; Watanabe, I; ITOH, H; Mri, T. A new aminoglycoside antibiotic, sannamycin C and its 4-N-glycyl derivative. J Antibiot 1980, 33, 1274–1280. [Google Scholar]
- Bell, KS; Philp, JC; Aw, DWJ; Christofi, N. The genus Rhodococcus. J Appl Microbiol 1998, 85, 195–210. [Google Scholar]
- Iwatsuki, M; Tomoda, H; Uchida, R; Gouda, H; Hirono, S; Omura, S. Lariatins, antimycobacterial peptides produced by Rhodococcus sp. K01-B0171, have a lasso structure. J Am Chem Soc 2006, 128. [Google Scholar]
- Wicke, C; Hüners, M; Wray, V; Nimtz, M; Bilitewski, U; Lang, S. Production and structure elucidation of glycoglycerolipids from a marine sponge-associated Microbacterium species. J Nat Prod 2000, 63, 621–626. [Google Scholar]
- Shirling, EB; Gottlieb, D. Methods for characterization of Streptomyces species. Int J Syst Bacteriol 1966, 16, 313–340. [Google Scholar]
- Olson, JB; Lord, CC; McCarthy, PJ. Improved recoverability of microbial colonies from sponge samples. Mar Microb Ecol 2000, 40, 139–147. [Google Scholar]
- Lechevalier, MP. Actinomycetes of sewage-treatment plants. Environ Protection Technol Ser 1975. EPA-600/2-75-031. [Google Scholar]
- Weiner, RM; Segall, AM; Colwell, RR. Characterization of a marine bacterium associated with Crassostrea virginica (the eastern oyster). Appl Environ Microbiol 1985, 49, 83–90. [Google Scholar]
- Reasoner, DJ; Geldreich, EE. A new medium for the enumeration and subculture of bacteria from potable water. Appl Environ Microbiol 1985, 49, 1–7. [Google Scholar]
- Lyman, J; Fleming, R. Composition of seawater. J Mar Res 1940, 3, 134–146. [Google Scholar]
- Hentschel, U; Schmid, M; Wagner, M; Fieseler, L; Gernert, C; Hacker, J. Isolation and phylogenetic analysis of bacteria with antimicrobial activities from the mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. FEMS Microbiol Ecol 2001, 35, 305–312. [Google Scholar]
- Lane, DJ. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E, Goodfellow, M, Eds.; John Wiley: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Ashelford, KE; Chuzhanova, NA; Fry, JC; Jones, AJ; Weightman, AJ. At least 1 in 20 16S rRNA sequence records currently held in public repositories is estimated to contain substantial anomalies. Appl Environ Microbiol 2006, 71, 7724–7736. [Google Scholar]
- Wang, Q; Garrity, GM; Tiedje, JM; Cole, JR. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol 2007, 73(16), 5261–5267. [Google Scholar]
- Ludwig, W; Strunk, O; Westram, R; Richter, L; Meier, H; Yadhukumar; Buchner, A; Lai, T; Steppi, S; Jobb, G; et al. ARB: A software environment for sequence data. Nuc Acid Res 2004, 32, 1363–1371. [Google Scholar]
- Inderlied, CB; Salfinger, M. Antimicrobial agents and susceptibility tests: Mycobacteria. In Manual of Clinical Microbiology, 6th ed; Murray, PR, Baron, EJ, Pfaller, MA, Tenover, FC, Yolken, RH, Eds.; American Society for Microbiology: Washington DC, USA, 1995; pp. 1385–1404. [Google Scholar]
- Ponte-Sucre, A; Vicik, R; Schultheis, M; Schirmeister, T; Moll, H. Aziridine-2,3-dicarboxylates, peptidomimetic cysteine protease inhibitors with antileishmanial activity. Antimicrob Agents Chemother 2006, 50, 2439–2447. [Google Scholar]
- Huber, W; Koella, JC. A Comparison of three methods of estimating ec50 in studies of drug resistance of malaria parasites. Acta Trop 1993, 55, 257–261. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sponge isolate | Inhibition zone diameter (mm) | ||
|---|---|---|---|
| S. aureus 8325 | E. faecalis JH212 | C. albicans S314 | |
| Actinokineospora sp. EG49* | 0 | 0 | 12 |
| Dietzia sp. EG67 | 13 | 0 | 0 |
| Microbacterium sp. EG69* | 10 | 9 | 0 |
| Micromonospora sp. RV115 | 12 | 10 | 0 |
| Rhodococcus sp. EG33* | 12 | 8 | 0 |
| Rubrobacter sp. RV113** | 9 | 0 | 0 |
| Streptomyces sp. RV15* | 17 | 11 | 13 |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Abdelmohsen, U.R.; Pimentel-Elardo, S.M.; Hanora, A.; Radwan, M.; Abou-El-Ela, S.H.; Ahmed, S.; Hentschel, U. Isolation, Phylogenetic Analysis and Anti-infective Activity Screening of Marine Sponge-Associated Actinomycetes. Mar. Drugs 2010, 8, 399-412. https://doi.org/10.3390/md8030399
Abdelmohsen UR, Pimentel-Elardo SM, Hanora A, Radwan M, Abou-El-Ela SH, Ahmed S, Hentschel U. Isolation, Phylogenetic Analysis and Anti-infective Activity Screening of Marine Sponge-Associated Actinomycetes. Marine Drugs. 2010; 8(3):399-412. https://doi.org/10.3390/md8030399
Chicago/Turabian StyleAbdelmohsen, Usama Ramadan, Sheila M. Pimentel-Elardo, Amro Hanora, Mona Radwan, Soad H. Abou-El-Ela, Safwat Ahmed, and Ute Hentschel. 2010. "Isolation, Phylogenetic Analysis and Anti-infective Activity Screening of Marine Sponge-Associated Actinomycetes" Marine Drugs 8, no. 3: 399-412. https://doi.org/10.3390/md8030399
APA StyleAbdelmohsen, U. R., Pimentel-Elardo, S. M., Hanora, A., Radwan, M., Abou-El-Ela, S. H., Ahmed, S., & Hentschel, U. (2010). Isolation, Phylogenetic Analysis and Anti-infective Activity Screening of Marine Sponge-Associated Actinomycetes. Marine Drugs, 8(3), 399-412. https://doi.org/10.3390/md8030399