Anthracenedione Derivatives as Anticancer Agents Isolated from Secondary Metabolites of the Mangrove Endophytic Fungi



Abstract
:1. Introduction
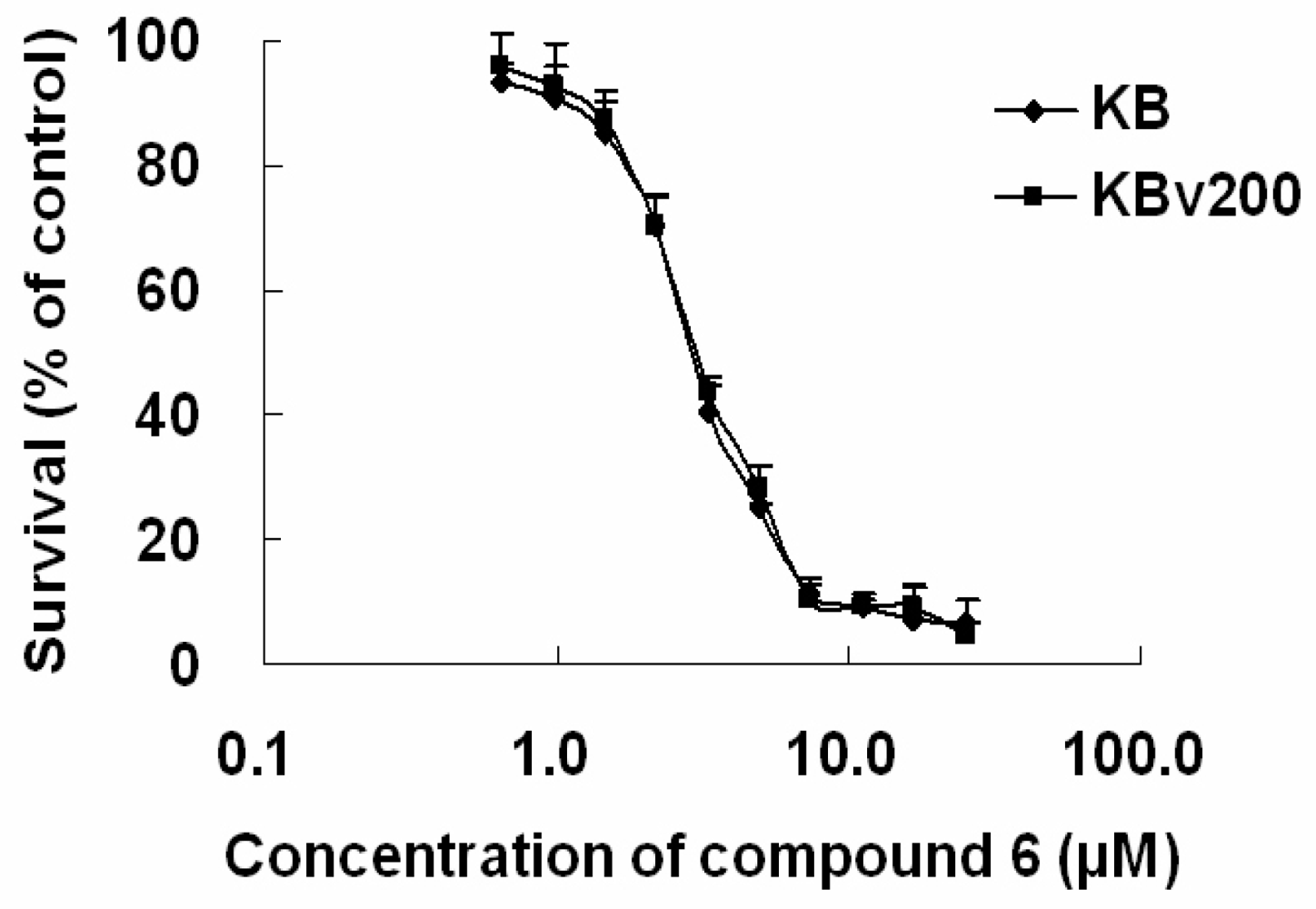
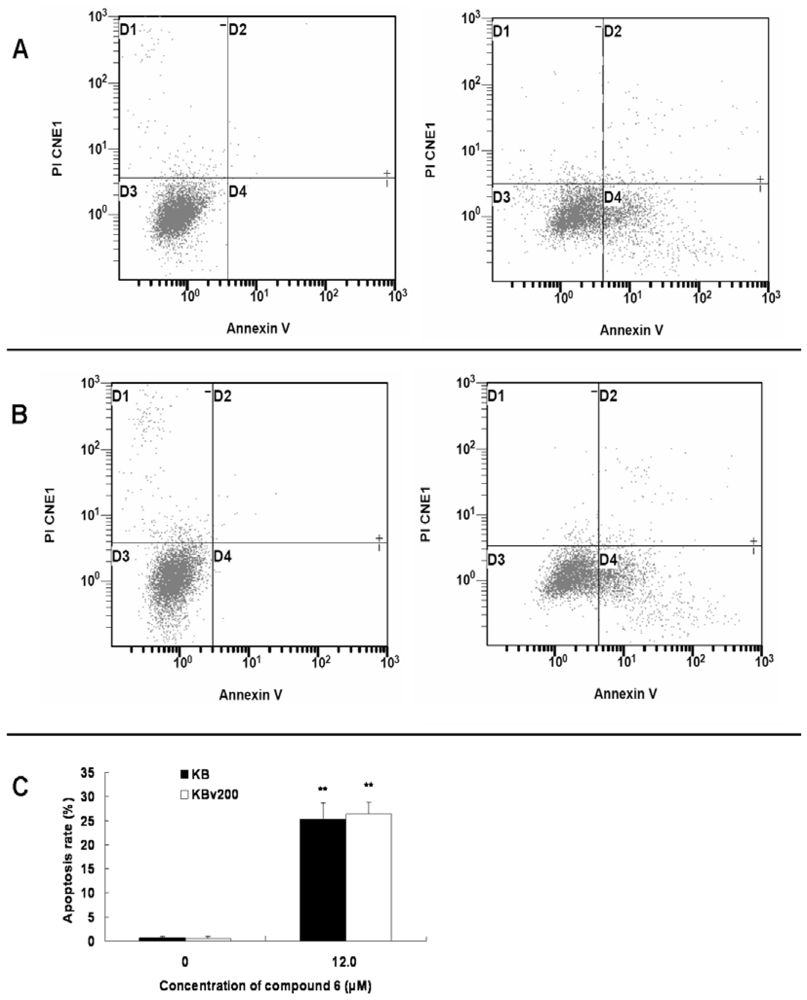
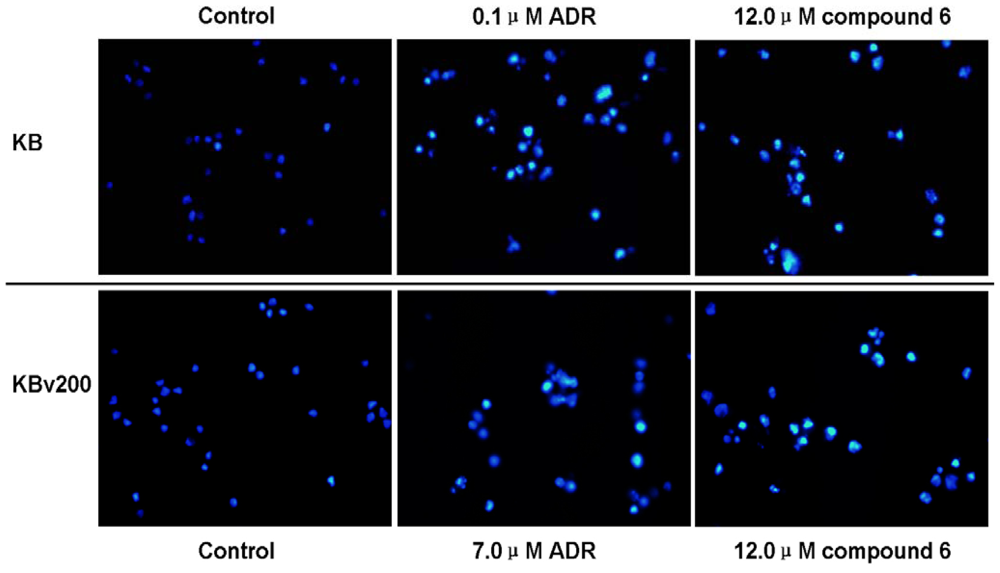
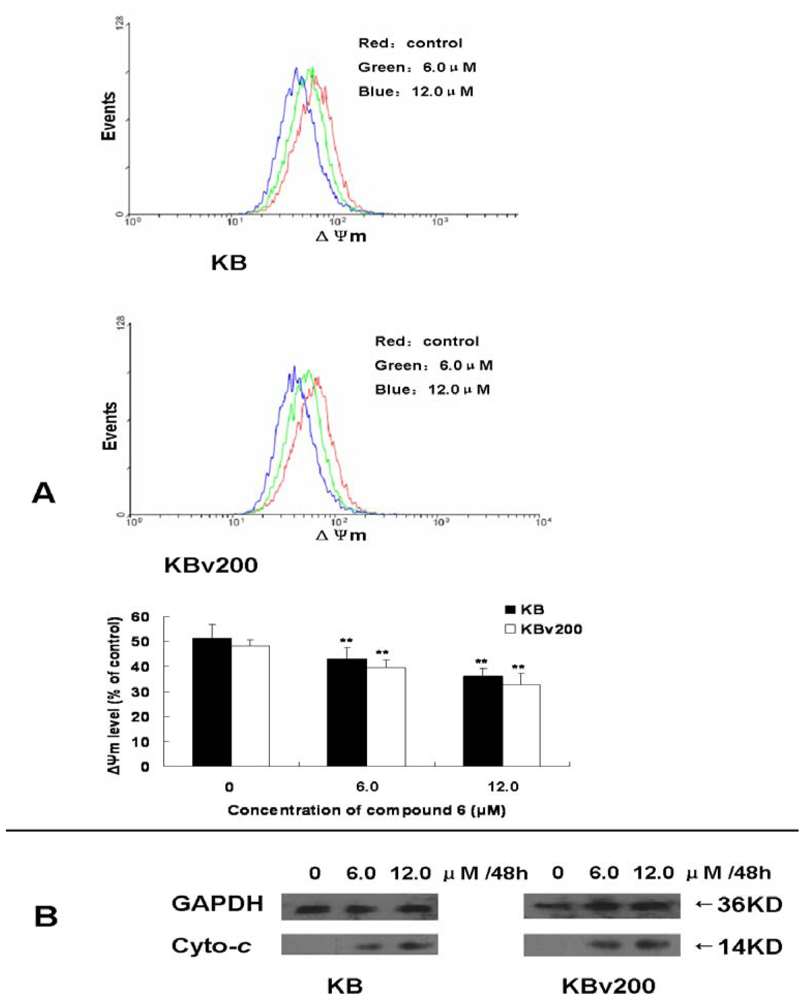
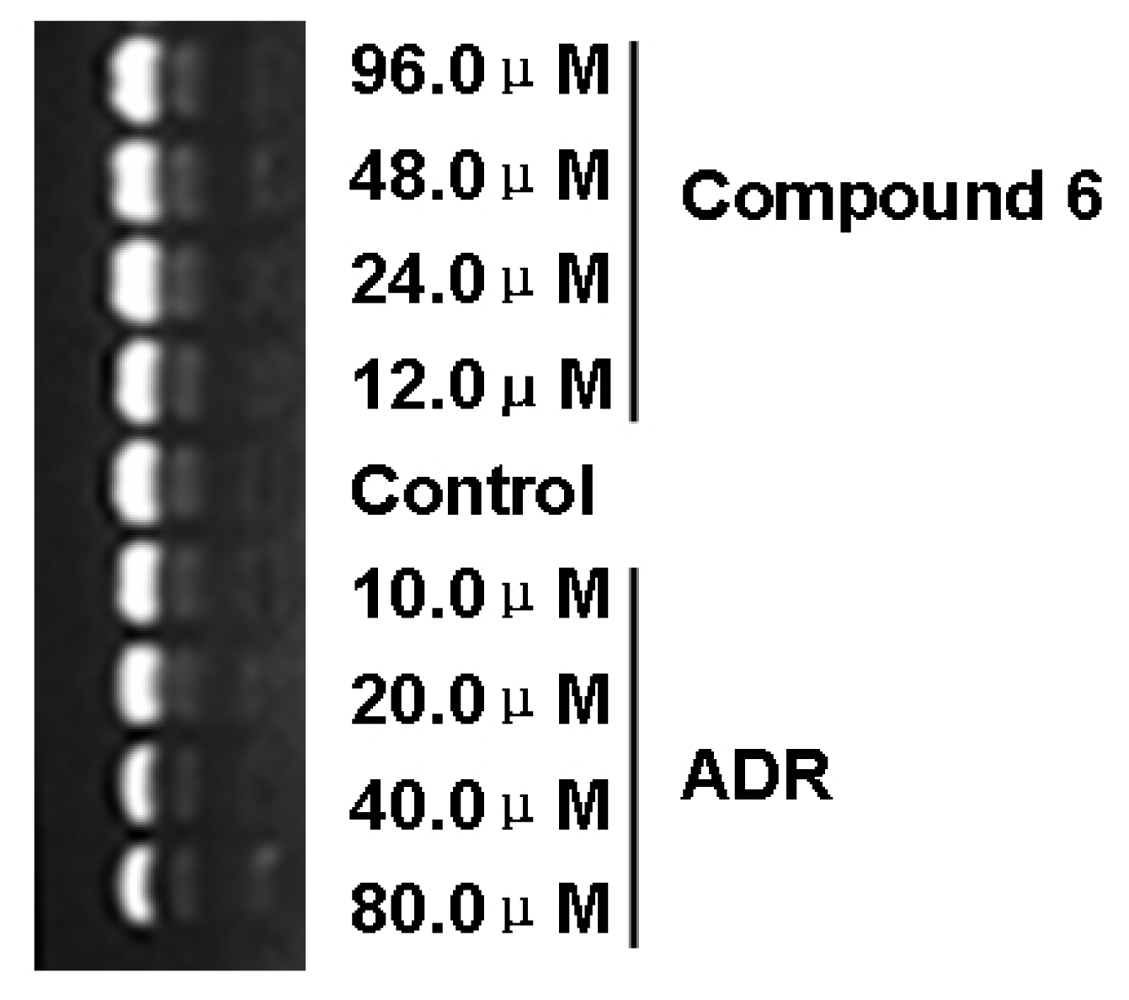
2. Results and Discussion
3. Experimental Section
3.1. Chemicals and reagents
3.2. Fermentation of fungi, extraction, isolation and identification of compounds
3.3. Cell viability assay
3.4. Determination of mitochondrial potential (ΔΨm)
3.5. Annexin V-FITC/PI assay
3.6. Hoechst 33258 staining
3.7. Subcellular fractionation for western blot analysis of cytosolic cytochrome c
3.8. DNA binding assay
4. Conclusions
Acknowledgements
- Samples Availability: Available from the authors.
References and Notes
- Smyth, MJ; Krasovskis, E; Sutton, VR; Johnstone, RW. The drug efflux protein, P-glycoprotein, additionally protects drug-resistant tumor cells from multiple forms of caspase-dependent apoptosis. Proc Natl Acad Sci USA 1998, 95, 7024–7029. [Google Scholar]
- Ruefli, AA; Tainton, KM; Darcy, PK; Smyth, MJ; Johnstone, RW. P-glycoprotein inhibits caspase-8 activation but not formation of the death inducing signal complex (disc) following Fas ligation. Cell Death Differ 2002, 9, 1266–1272. [Google Scholar]
- Shi, Z; Peng, XX; Kim, IW; Shukla, S; Si, QS; Robey, RW; Bates, SE; Shen, T; Ashby, CR, Jr; Fu, LW; Ambudkar, SV; Chen, ZS. Erlotinib (Tarceva, OSI-774) antagonizes ATP-binding cassette subfamily B member 1 and ATP-binding cassette subfamily G member 2-mediated drug resistance. Cancer Res 2007, 67, 11012–11020. [Google Scholar]
- Cooper, EL. Drug Discovery, CAM and Natural Products. Evid Based Complement Alternat Med 2004, 1, 215–217. [Google Scholar]
- D’Incalci, M; Simone, M; Tavecchio, M; Damia, G; Garbi, A; Erba, E. New drugs from the sea. J Chemother 2004, 16(Suppl 4), 86–89. [Google Scholar]
- Simmons, TL; Andrianasolo, E; McPhail, K; Flatt, P; Gerwick, WH. Marine natural products as anticancer drugs. Mol Cancer Ther 2005, 4, 333–342. [Google Scholar]
- Molinski, TF; Dalisay, DS; Lievens, SL; Saludes, JP. Drug development from marine natural products. Nat Rev Drug Discov 2009, 8, 69–85. [Google Scholar]
- Gordaliza, M. Natural products as leads to anticancer drugs. Clin Transl Oncol 2007, 9, 767–776. [Google Scholar]
- Sufrin, JR; Finckbeiner, S; Oliver, CM. Marine-derived metabolites of S-adenosylmethionine as templates for new anti-infectives. Mar Drugs 2009, 7, 401–434. [Google Scholar]
- Baerga-Ortiz, A. Biotechnology and biochemistry of marine natural products. P R Health Sci J 2009, 28, 251–257. [Google Scholar]
- Olano, C; Mendez, C; Salas, JA. Antitumor compounds from marine actinomycetes. Mar Drugs 2009, 7, 210–248. [Google Scholar]
- Li, DL; Li, XM; Wang, BG. Natural anthraquinone derivatives from a marine mangrove plant-derived endophytic fungus Eurotium rubrum: structural elucidation and DPPH radical scavenging activity. J Microbiol Biotechnol 2009, 19, 675–680. [Google Scholar]
- Xia, XK; Huang, HR; She, ZG; Shao, CL; Liu, F; Cai, XL; Vrijmoed, LLP; Lin, YC. (1)H and (13)C NMR assignments for five anthraquinones from the mangrove endophytic fungus Halorosellinia sp. (No. 1403). Magn Reson Chem 2007, 45, 1006–1009. [Google Scholar]
- She, ZG; Huang, HR; Lin, YC; Vrijmoed, LLP; Jones, EBG. 1,4-dihydroxy-2-methoxy-7-methylanthracene-9,10-dione. Acta Crystallogr Sect E 2006, 62, O3737–3738. [Google Scholar]
- Jiang, GC; Lin, YC; Zhou, SN; Vrijmoed, LLP; Jones, EBG. Studies on the Secondary Metabolites of Mangrove Fungus No. 1403 from the South China Sea. Acta Scientiarum Naturalium Universitatis Sun yat-sen 2000, 39, 68–72. [Google Scholar]
- Tatyana, K; Francesca, E; Luca, Z; Giovanni, F; Cheng, YC; Ginger, ED; Enzo, T. Inhibition of HIV-1 ribonuclease H activity by novel frangula-emodine derivatives. Med Chem 2009, 5, 398–410. [Google Scholar]
- Park, MY; Kwon, HJ; Sung, MK. Evaluation of aloin and aloe-emodin as anti-inflammatory agents in aloe by using murine macrophages. Biosci Biotechnol Biochem 2009, 73, 828–832. [Google Scholar]
- Yan, YY; Su, XD; Liang, YJ; Zhang, JY; Shi, CJ; Lu, Y; Gu, LQ; Fu, LW. Emodin azide methyl anthraquinone derivative triggers mitochondrial-dependent cell apoptosis involving in caspase-8-mediated Bid cleavage. Mol Cancer Ther 2008, 7, 1688–1697. [Google Scholar]
- Huang, H; Jia, Q; Ma, J; Qin, G; Chen, Y; Xi, Y; Lin, L; Zhu, W; Ding, J; Jiang, H; Liu, H. Discovering novel quercetin-3-O-amino acid-esters as a new class of Src tyrosine kinase inhibitors. Eur J Med Chem 2009, 44, 1982–1988. [Google Scholar]
- Chiu, TH; Lai, WW; Hsia, TC; Yang, JS; Lai, TY; Wu, PP; Ma, CY; Yeh, CC; Ho, CC; Lu, HF; Wood, WG; Chung, JG. Aloe-emodin induces cell death through S-phase arrest and caspase-dependent pathways in human tongue squamous cancer SCC-4 cells. Anticancer Res 2009, 29, 4503–4511. [Google Scholar]
- Nadanaciva, S; Will, Y. The role of mitochondrial dysfunction and drug safety. Idrugs 2009, 12, 706–710. [Google Scholar]
- Johannsen, DL; Ravussin, E. The role of mitochondria in health and disease. Curr Opin Pharmacol 2009, 9, 780–786. [Google Scholar]
- George, NM; Targy, N; Evans, JJ; Zhang, L; Luo, X. Bax contains two functional mitochondrial targeting sequences and translocates to mitochondria in a conformational change- and homo-oligomerization-driven process. J Biol Chem 2010, 285, 1384–1392. [Google Scholar]
- Rabbani, A; Finn, RM; Ausio, J. The anthracycline antibiotics: antitumor drugs that alter chromatin structure. Bioessays 2005, 27, 50–56. [Google Scholar]
- Moreno-Aspitia, A; Perez, EA. Anthracycline- and/or taxane-resistant breast cancer: results of a literature review to determine the clinical challenges and current treatment trends. Clin Ther 2009, 57, 1619–1640. [Google Scholar]
- Rohr, J. Comparison of multicyclic polyketides by folding analysis: a novel approach to recognize biosynthetic and/or evolutionary interrelationships of the natural products or intermediates and its exemplification on hepta-, octa-, and decaketides. J Org Chem 1992, 57, 5217–5223. [Google Scholar]
- Khan, F; Sherwani, AF; Afzal, M. Chromosomal aberration and micronucleus studies of two topoisomerase (II) targeting anthracyclines. J Environ Biol 2009, 30, 409–412. [Google Scholar]
- Zhang, JY; Wu, HY; Xia, XK; Liang, YJ; Yan, YY; She, ZG; Lin, YC; Fu, LW. Anthracenedione derivative 1403P-3 induces apoptosis in KB and KBv200 cells via reactive oxygen species-independent mitochondrial pathway and death receptor pathway. Cancer Biol Ther 2007, 6, 1413–1421. [Google Scholar]
- Zagotto, G; Sissi, C; Moro, S; Dal, Ben D; Parkinson, GN; Fox, KR; Neidle, S; Palumbo, M. Amide bond direction modulates G-quadruplex recognition and telomerase inhibition by 2,6 and 2,7 bis-substituted anthracenedione derivatives. Bioorg Med Chem 2008, 16, 354–361. [Google Scholar]
- Raghava, S; Kompella, UB. AQ4, an antitumor anthracenedione, inhibits endothelial cell proliferation and vascular endothelial growth factor secretion: implications for the therapy of ocular neovascular disorders. Eur J Pharmacol 2007, 568, 68–74. [Google Scholar]
- Dai, CL; Tiwari, AK; Wu, CP; Su, XD; Wang, SR; Liu, DG; Ashby, CR, Jr; Huang, Y; Robey, RW; Liang, YJ; Chen, LM; Shi, CJ; Ambudkar, SV; Chen, ZS; Fu, LW. Lapatinib (Tykerb, GW572016) reverses multidrug resistance in cancer cells by inhibiting the activity of ATP-binding cassette subfamily B member 1 and G member 2. Cancer Res 2008, 68, 7905–7914. [Google Scholar]
- Zhang, JY; Tao, LY; Liang, YJ; Yan, YY; Dai, CL; Xia, XK; She, ZG; Lin, YC; Fu, LW. Secalonic acid D induced leukemia cell apoptosis and cell cycle arrest of G(1) with involvement of GSK-3beta/beta-catenin/c-Myc pathway. Cell Cycle 2009, 8, 2444–2450. [Google Scholar]
- Wang, XH; Jia, DZ; Liang, YJ; Yan, SL; Ding, Y; Chen, LM; Shi, Z; Zeng, MS; Liu, GF; Fu, LW. Lgf-YL-9 induces apoptosis in human epidermoid carcinoma KB cells and multidrug resistant KBv200 cells via reactive oxygen species-independent mitochondrial pathway. Cancer Lett 2006, 249, 256–270. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
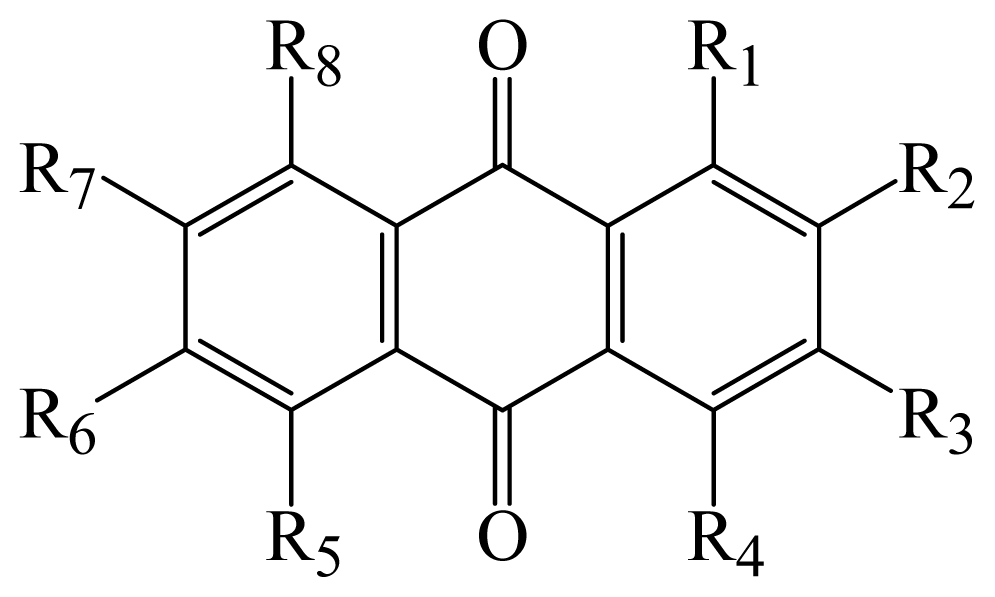
| Compound | R1 | R2 | R3 | R4 | R6 | R7 | R8 | Cytotoxicity KB | IC50 ( μM ) KBv200 |
| 1 | methoxy | — | methoxy | — | methyl | — | — | 57.32 | 90.86 |
| 2 | hydroxy | methoxy | — | hydroxy | — | methyl | — | 305.14 | * |
| 3 | hydroxy | — | methyl | — | — | — | hydroxy | 174.87 | 331.97 |
| 4 | hydroxy | methoxy | — | hydroxy | methyl | hydroxy | — | * | * |
| 5 | hydroxy | — | hydroxy | — | methyl | — | 114.09 | 86.45 | |
| 6 | hydroxy | — | methyl | — | — | — | — | 3.17 | 3.21 |
| 7 | hydroxy | — | — | — | — | — | hydroxy | 56.56 | 109.15 |
| 8 | hydroxy | — | hydroxy | — | — | — | hydroxy | * | 72.60 |
| 9 | hydroxy | — | hydroxy | — | methoxy | — | methyl | 38.05 | 34.64 |
| 10 | hydroxy | — | hydroxy | — | methyl | — | hydroxy | * | * |
| 11 | methoxy | — | methyl | — | — | — | hydroxy | * | 185.68 |
| 12 | hydroxy | — | hydroxy | — | methyl | — | methoxy | * | 190.81 |
| 13 | hydroxy | methoxy | — | methoxy | methyl | hydroxy | — | * | 301.47 |
| 14 | hydroxy | methoxy | methoxy | methyl | 68.39 | 243.69 | |||
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, J.-y.; Tao, L.-y.; Liang, Y.-j.; Chen, L.-m.; Mi, Y.-j.; Zheng, L.-s.; Wang, F.; She, Z.-g.; Lin, Y.-c.; To, K.K.W.; et al. Anthracenedione Derivatives as Anticancer Agents Isolated from Secondary Metabolites of the Mangrove Endophytic Fungi. Mar. Drugs 2010, 8, 1469-1481. https://doi.org/10.3390/md8041469
Zhang J-y, Tao L-y, Liang Y-j, Chen L-m, Mi Y-j, Zheng L-s, Wang F, She Z-g, Lin Y-c, To KKW, et al. Anthracenedione Derivatives as Anticancer Agents Isolated from Secondary Metabolites of the Mangrove Endophytic Fungi. Marine Drugs. 2010; 8(4):1469-1481. https://doi.org/10.3390/md8041469
Chicago/Turabian StyleZhang, Jian-ye, Li-yang Tao, Yong-ju Liang, Li-ming Chen, Yan-jun Mi, Li-sheng Zheng, Fang Wang, Zhi-gang She, Yong-cheng Lin, Kenneth Kin Wah To, and et al. 2010. "Anthracenedione Derivatives as Anticancer Agents Isolated from Secondary Metabolites of the Mangrove Endophytic Fungi" Marine Drugs 8, no. 4: 1469-1481. https://doi.org/10.3390/md8041469
APA StyleZhang, J. -y., Tao, L. -y., Liang, Y. -j., Chen, L. -m., Mi, Y. -j., Zheng, L. -s., Wang, F., She, Z. -g., Lin, Y. -c., To, K. K. W., & Fu, L. -w. (2010). Anthracenedione Derivatives as Anticancer Agents Isolated from Secondary Metabolites of the Mangrove Endophytic Fungi. Marine Drugs, 8(4), 1469-1481. https://doi.org/10.3390/md8041469