Are Known Cyanotoxins Involved in the Toxicity of Picoplanktonic and Filamentous North Atlantic Marine Cyanobacteria?




Abstract
:1. Introduction
2. Materials and Methods
2.1. Cyanobacteria strains
2.2. Identification of cyanobacteria isolates
Molecular identification
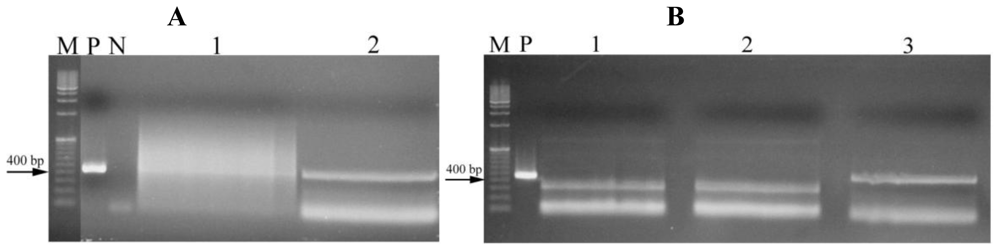
2.3. Toxin genes detection
2.4. Peptides screening by MALDI-TOF MS
2.5. Cyanobacteria extracts for Artemia salina bioassay
2.6. Artemia salina bioassay
3. Results
3.1. Cyanobacteria strains
3.2. Detection of genes involved in toxin production
3.3. MALDI-TOF MS
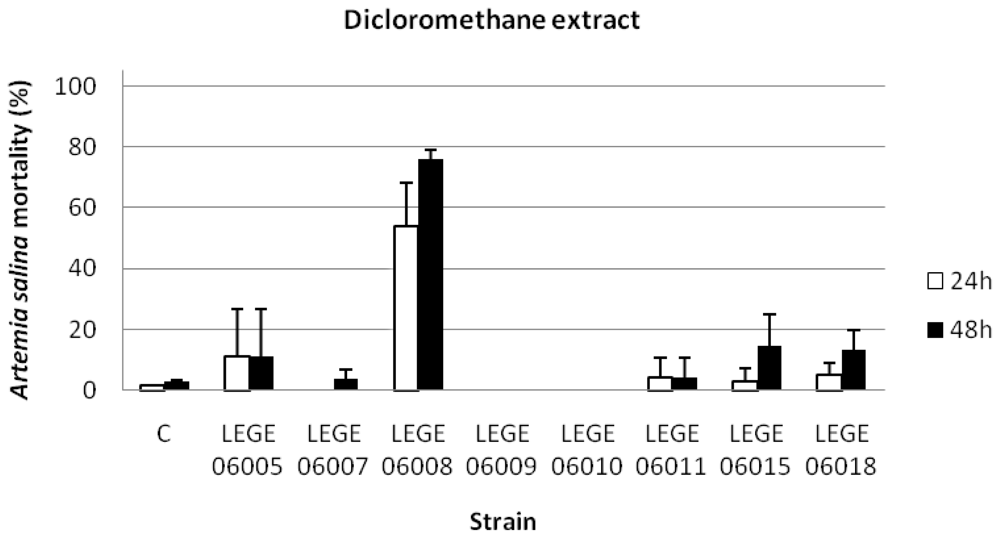
3.4. Artemia salina assay
4. Discussion
Acknowledgements
- Samples Availability: Available from the authors.
References
- Tan, LT. Bioactive natural products from marine cyanobacteria for drug discovery. Phytochemistry 2007, 68, 954–979. [Google Scholar]
- Dittmann, E; Neilan, BA; Borner, T. Molecular biology of peptide and polyketide biosynthesis in cyanobacteria. Appl. Microbiol. Biotechnol 2001, 57, 467–473. [Google Scholar]
- Volk, RB. Screening of microalgae for species excreting norharmane, a manifold biologically active indole alkaloid. Microbiol. Res 2008, 163, 307–313. [Google Scholar]
- Codd, GA; Morrison, LF; Metcalf, JS. Cyanobacterial toxins: risk management for health protection. Toxicol. Appl. Pharmacol 2005, 203, 264–272. [Google Scholar]
- Wiegand, C; Pflugmacher, S. Ecotoxicological effects of selected cyanobacterial secondary metabolites: a short review. Toxicol. Appl. Pharmacol 2005, 203, 201–218. [Google Scholar]
- Albert, S; O’Neil, JM; Udy, JW; Ahern, KS; O’Sullivan, CM; Dennison, WC. Blooms of the cyanobacterium Lyngbya majuscula in coastal Queensland, Australia: disparate sites, common factors. Mar. Pollut. Bull 2005, 51, 428–437. [Google Scholar]
- Pittman, SJ; Pittman, KM. Short-term consequences of a benthic cyanobacterial bloom (Lyngbya majuscula Gomont) for fish and penaeid prawns in Moreton Bay (Queensland, Australia). Estuar. Coast. Shelf Sci 2005, 63, 619–632. [Google Scholar]
- Camargo, JA; Alonso, A. Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: A global assessment. Environ. Int 2006, 32, 831–849. [Google Scholar]
- Ahern, KS; Ahern, CR; Udy, JW. In situ field experiment shows Lyngbya majuscula (cyanobacterium) growth stimulated by added iron, phosphorus and nitrogen. Harmful Algae 2007, 7, 389–404. [Google Scholar]
- Hakanson, L; Bryhn, AC; Hytteborn, JK. On the issue of limiting nutrient and predictions of cyanobacteria in aquatic systems. Sci. Total Environ 2007, 379, 89–108. [Google Scholar]
- Pliński, M; Mazur-Marzec, H; Jóźwiak, T; Kobos, J. The potential causes of cyanobacterial blooms in Baltic Sea estuaries. Oceanol. Hydrobiol. Stud 2007, 36, 125–137. [Google Scholar]
- Martins, R; Pereira, P; Welker, M; Fastner, J; Vasconcelos, VM. Toxicity of culturable cyanobacteria strains isolated from the Portuguese coast. Toxicon 2005, 46, 454–464. [Google Scholar]
- Martins, R; Fernandez, N; Beiras, R; Vasconcelos, V. Toxicity assessment of crude and partially purified extracts of marine Synechocystis and Synechococcus cyanobacterial strains in marine invertebrates. Toxicon 2007, 50, 791–799. [Google Scholar]
- Selheim, F; Herfindal, L; Martins, R; Vasconcelos, V; Doskeland, SO. Neuro-apoptogenic and blood platelet targeting toxins in benthic marine cyanobacteria from the Portuguese coast. Aquat. Toxicol 2005, 74, 294–306. [Google Scholar]
- Martins, RF; Ramos, MF; Herfindal, L; Sousa, JA; Skaerven, K; Vasconcelos, VM. Antimicrobial and cytotoxic assessment of marine cyanobacteria - Synechocystis and Synechococcus. Mar. Drugs 2008, 6, 1–11. [Google Scholar]
- Kotai, J. Instructions for Preparation of Modified Nutrient Solution Z8 for Algae; Norwegian Institute for Water Research B-11769: Blindern, Oslo, Norway, 1972; p. 5. [Google Scholar]
- Baker, P. Identification of Common Noxious Cyanobacteria, Part II Chroococcales, Oscillatoriales; Urban Water Research Association of Australia: Melbourne, Australia, 1992. [Google Scholar]
- Rippka, R. Recognition and Identification of Cyanobacteria, Methods in Enzymology; Academic press Inc: New York, NY, USA, 1998; pp. 28–67. [Google Scholar]
- Komárek, J; Anagnostidis, K. Cyanoprokariota. 1. Teil: Chroococcales; Gustav Fischer: Berlin, Germany, 1999. [Google Scholar]
- Castenholz, RW; Boone, DR. Garrity, GM, Ed.; The Archae and the Deeply Branching and Phototrophic Bacteria. In Bergey’s Manual of Systematic Bacteriology; Springer: New York, NY, USA, 2001. [Google Scholar]
- Cronberg, G; Annadotter, H. Manual on Aquatic Cyanobacteria-A Photo Guide and a Synopsis of Their Toxicology; Internatinal Society for the Study of Harmful Algae, United Nations Educational, Scientific and Cultural Organization: Denmark, 2006. [Google Scholar]
- Neilan, BA; Jacobs, D; Del Dot, T; Blackall, LL; Hawkins, PR; Cox, PT; Goodman, AE. rRNA sequences and evolutionary relationships among toxic and nontoxic cyanobacteria of the genus Microcystis. Int. J. Syst. Bacteriol 1997, 47, 693–697. [Google Scholar]
- Jungblut, AD; Hawes, I; Mountfort, D; Hitzfeld, B; Dietrich, DR; Burns, BP; Neilan, BA. Diversity within cyanobacterial mat communities in variable salinity meltwater ponds of McMurdo Ice Shelf, Antarctica. Environ. Microbiol 2005, 7, 519–529. [Google Scholar]
- Pearson, LA; Neilan, BA. The molecular genetics of cyanobacterial toxicity as a basis for monitoring water quality and public health risk. Curr. Opin. Biotechnol 2008, 19, 281–288. [Google Scholar]
- Moffitt, MC; Neilan, BA. Characterization of the nodularin synthetase gene cluster and proposed theory of the evolution of cyanobacterial hepatotoxins. Appl. Env. Microbiol 2004, 70, 6353–6362. [Google Scholar]
- Jungblut, AD; Neilan, BA. Molecular identification and evolution of the cyclic peptide hepatotoxins, microcystin and nodularin, synthetase genes in three orders of cyanobacteria. Arch. Microbiol 2006, 185, 107–114. [Google Scholar]
- Fergusson, KM; Saint, CP. Multiplex PCR assay for Cylindrospermopsis raciborskii and cylindrospermopsin-producing cyanobacteria. Environ. Toxicol 2003, 18, 120–125. [Google Scholar]
- Hisbergues, M; Christiansen, G; Rouhiainen, L; Sivonen, K; Borner, T. PCR-based identification of microcystin-producing genotypes of different cyanobacterial genera. Arch. Microbiol 2003, 180, 402–410. [Google Scholar]
- Schembri, MA; Neilan, BA; Saint, CP. Identification of genes implicated in toxin production in the cyanobacterium Cylindrospermopsis raciborskii. Environ. Toxicol 2001, 16, 413–421. [Google Scholar]
- Welker, M; Marsalek, B; Sejnohova, L; von Dohren, H. Detection and identification of oligopeptides in Microcystis (cyanobacteria) colonies: toward an understanding of metabolic diversity. Peptides 2006, 27, 2090–2103. [Google Scholar]
- Carballo, JL; Hernández-Inda, ZL; Pérez, P; García-Grávalos, MD. A comparison between two brine shrimp assays to detect in vitro cytotoxicity in marine natural products. BMC Biotechnology 2002, 2, 17. [Google Scholar]
- Christiansen, G; Fastner, J; Erhard, M; Borner, T; Dittmann, E. Microcystin biosynthesis in Planktothrix: genes, evolution, and manipulation. J. Bacteriol 2003, 185, 564–572. [Google Scholar]
- Rantala, A; Fewer, DP; Hisbergues, M; Rouhiainen, L; Vaitomaa, J; Borner, T; Sivonen, K. Phylogenetic evidence for the early evolution of microcystin synthesis. Proc. Natl. Acad. Sci. USA 2004, 101, 568–573. [Google Scholar]
- Tsuge, K; Akiyama, T; Shoda, M. Cloning, sequencing, and characterization of the iturin A operon. J. Bacteriol 2001, 183, 6265–6273. [Google Scholar]
- Mikalsen, B; Boison, G; Skulberg, OM; Fastner, J; Davies, W; Gabrielsen, TM; Rudi, K; Jakobsen, KS. Natural variation in the synthase operon mcyABC and impact on microcystin production in Microcystis strains. J. Bacteriol 2003, 185, 2774–2785. [Google Scholar]
- Moffitt, MC; Neilan, BA. On the presence of peptide synthetase and polyketide synthase genes in the cyanobacterial genus Nodularia. FEMS Microbiol. Lett 2001, 196, 207–214. [Google Scholar]
- Kaebernick, M; Rohrlack, T; Christoffersen, K; Neilan, BA. A spontaneous mutant of microcystin biosynthesis: genetic characterization and effect on Daphnia. Environ. Microbiol 2001, 3, 669–679. [Google Scholar]
- Prati, M; Molteni, M; Pomati, F; Rossetti, C; Bernardini, G. Biological effect of the Planktothrix sp. FP1 cyanobacterial extract. Toxicon 2002, 40, 267–272. [Google Scholar]
- Rantala, A; Rajaniemi-Wacklin, P; Lyra, C; Lepistö, L; Rintala, J; Mankiewicz-Boczek, J; Sivonen, K. Detection of microcystin-producing cyanobacteria in Finnish lakes with genus-specific microcystin synthetase gene E (mcyE) PCR and associations with environmental factors. Appl. Environ. Microbiol 2006, 72, 6101–6110. [Google Scholar]
- Richardson, LL; Sekar, R; Myers, JL; Gantar, M; Voss, JD; Kaczmarsky, L; Remily, ER; Boyer, GL; Zimba, PV. The presence of the cyanobacterial toxin microcystin in black band disease of corals. FEMS Microbiol. Lett 2007, 272, 182–187. [Google Scholar]
- Gantar, M; Sekar, R; Richardson, LL. Cyanotoxins from black band disease of corals and from other coral reef environments. Microb. Ecol 2009, 58, 856–864. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain code | Substrate | Genus (16S rRNA highest match NCBI) | % similarity | Accession Nº |
|---|---|---|---|---|
| LEGE 06005 | Sea water | Synechococcus sp. UH7 | 92 | HM124558 |
| LEGE 06007 | Rock | Phormidium sp. HBC9 | 99 | HM124560 |
| LEGE 06008 | Rock | Cyanobium sp. NS01 | 99 | HM124561 |
| LEGE 06009 | Rock | Leptolyngbya sp. 0BB32S02 | 96 | HM124563 |
| LEGE 06010 | Rock | Leptolyngbya sp. 0BB32S02 | 98 | HM124564 |
| LEGE 06011 | Rock | Cyanobium sp. NS01 | 98 | HM124557 |
| LEGE 06015 | Patella sp. | Cyanobium sp. NS01 | 99 | HM124559 |
| LEGE 06018 | Rock | Oscillatoria sp. CCAP 1459/26 | 100 | HM124562 |
| Primer | Sequence (5′→3′) | A | B | Size (bp) | Amplified gene | Reference |
|---|---|---|---|---|---|---|
| 27F | AGAGTTTGATCCTGGCTCAG | 52 | 60 | 780 | 16S rRNA | [22] |
| 809R | GCTTCGGCACGGCTCGGGTCGATA | 64 | [23] | |||
| 740F | GGCYRWAWCTGACACTSAGGGA | - | 50 | 754 | 16S rRNA | [22] |
| 1494R | TACGGCTACCTTGTTACGAC | 56 | ||||
| mcyA-Cd F | AAAATTAAAAGCCGTATCAAA | 51 | 59 | 297 | Microcystin synthetase | [28] |
| mcyA-Cd R | AAAAGTGTTTTATTAGCGGCTCAT | 43 | ||||
| HEPF | TTTGGGGTTAACTTTTTTGGGCATAGTC | 57 | 52 | 472 | Microcystin/nodularin synthetase | [23] |
| HEPR | AATTCTTGAGGCTGTAAATCGGGTTT | 55 | ||||
| PKS M4 | GAAGCTCTGGAATCCGGTAA | 52 | 55 | 650 | Cylindrospermopsin polyketide synthase | [29] |
| PKS M5 | AATCCTTACGGGATCCGGTGC | 56 | [29] | |||
| M13 | GGCAAATTGTGATAGCCACGAGC | 57 | 55 | 597 | Cylindrospermopsin peptide synthetase | [29] |
| M14 | GATGGAACATCGCTCACTGGTG | 57 | [29] | |||
| Strain | Genus | mcyE gene (highest match NCBI) | % similarity | Accession Nº |
|---|---|---|---|---|
| LEGE 06010 | Leptolyngbya sp. | Microcystis sp. CYN06 microcystin synthetase E (mycE) gene, complete cds | 99% | HM124567 |
| LEGE 06018 | Oscillatoria sp. | Microcystis sp. CYN06 microcystin synthetase E (mycE) gene, complete cds | 99% | HM124566 |
| Strain | LEGE | LEGE | LEGE | LEGE | LEGE | LEGE | LEGE | LEGE | |
|---|---|---|---|---|---|---|---|---|---|
| Extract | 6005 | 6007 | 6008 | 6009 | 60010 | 6011 | 6015 | 6018 | |
| 24h | D | - | - | 51.4 | - | - | - | - | - |
| M | 51 | 51.3 | 50.6 | 50.9 | - | 49.9 | 50.8 | 51.4 | |
| A | 49.5 | 51.1 | 51 | 51.3 | 51.5 | 50.7 | 51 | 49.6 | |
| 48h | D | - | - | 51.2 | - | - | - | - | - |
| M | 50.8 | 50.7 | 49.9 | 50 | - | 48.3 | 50.7 | 51.2 | |
| A | 49.2 | 50.9 | 50.8 | 51.1 | 51.5 | 50.2 | 51 | 49.6 | |
© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Frazão, B.; Martins, R.; Vasconcelos, V. Are Known Cyanotoxins Involved in the Toxicity of Picoplanktonic and Filamentous North Atlantic Marine Cyanobacteria? Mar. Drugs 2010, 8, 1908-1919. https://doi.org/10.3390/md8061908
Frazão B, Martins R, Vasconcelos V. Are Known Cyanotoxins Involved in the Toxicity of Picoplanktonic and Filamentous North Atlantic Marine Cyanobacteria? Marine Drugs. 2010; 8(6):1908-1919. https://doi.org/10.3390/md8061908
Chicago/Turabian StyleFrazão, Bárbara, Rosário Martins, and Vitor Vasconcelos. 2010. "Are Known Cyanotoxins Involved in the Toxicity of Picoplanktonic and Filamentous North Atlantic Marine Cyanobacteria?" Marine Drugs 8, no. 6: 1908-1919. https://doi.org/10.3390/md8061908
APA StyleFrazão, B., Martins, R., & Vasconcelos, V. (2010). Are Known Cyanotoxins Involved in the Toxicity of Picoplanktonic and Filamentous North Atlantic Marine Cyanobacteria? Marine Drugs, 8(6), 1908-1919. https://doi.org/10.3390/md8061908