Gustatory Detection of Tetrodotoxin and Saxitoxin, and Its Competitive Inhibition by Quinine and Strychnine in Freshwater Fishes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
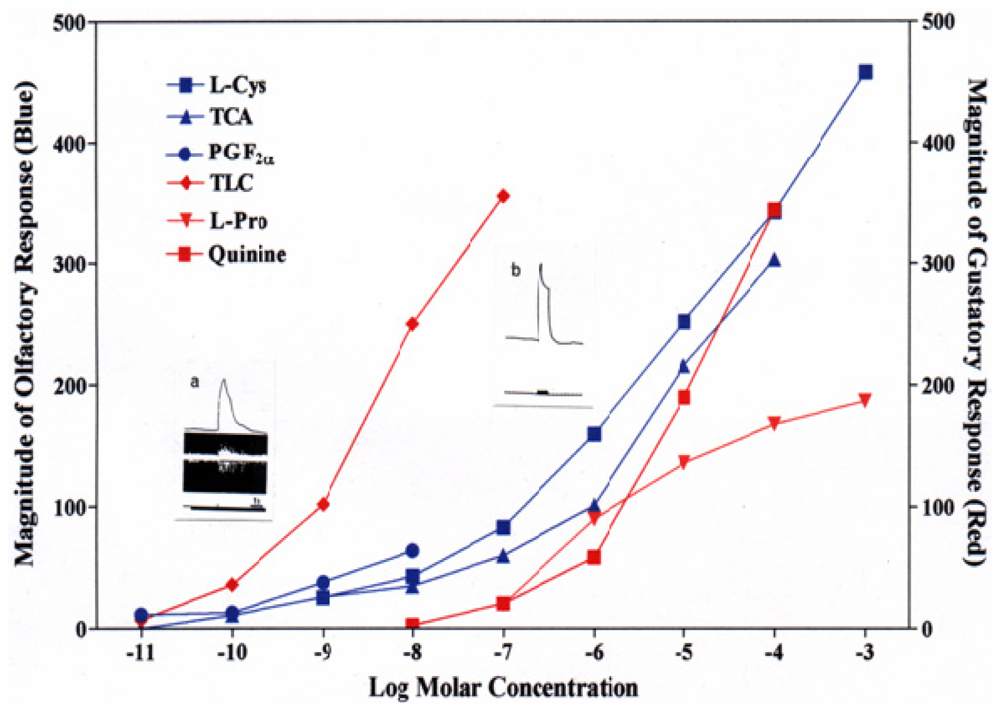
2. Characteristics of Fish Olfaction and Gustation
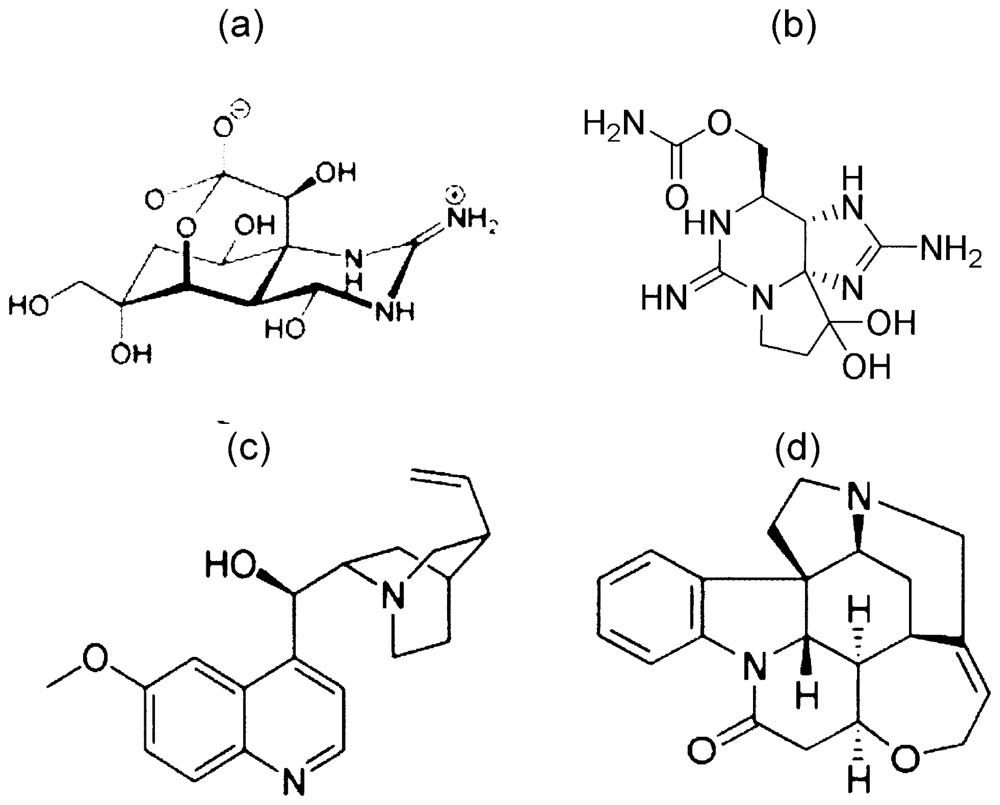
3. High Gustatory Sensitivity and Specificity to TTX and STX
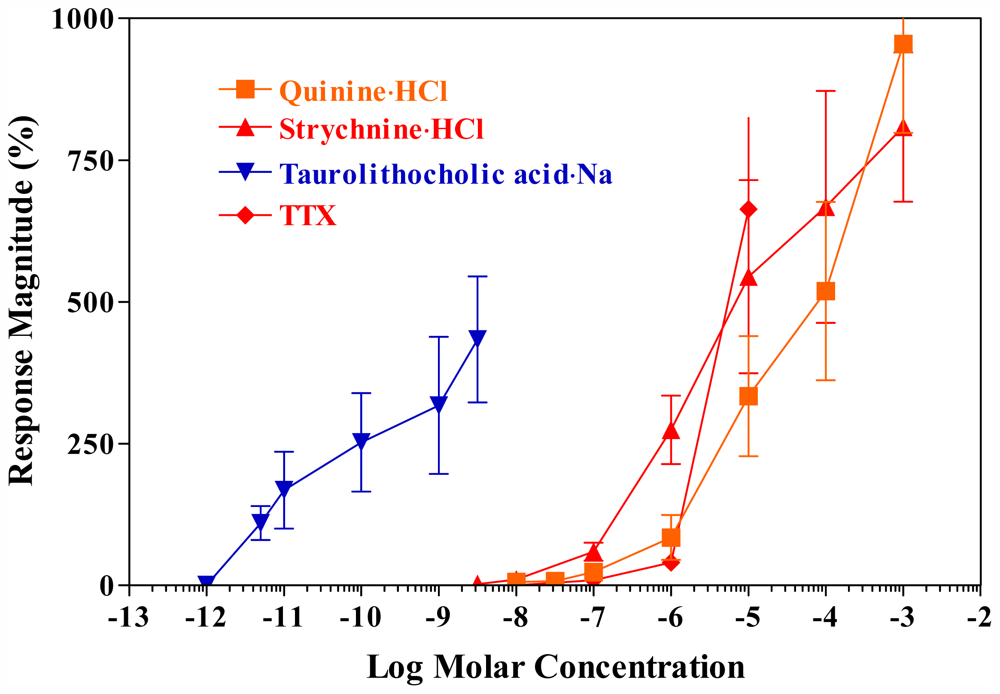
4. Competitive Inhibition of TTX Responses by Q·HCl, Strychnine and TLCA
5. Concluding Remarks
References
- Hara, T.J. Chemoreception (Smell and Taste): An Introduction. In Encyclopedia of Fish Physiology: From Genome to Environment; Farrell, A.P., Ed.; Academic Press: San Diego, CA, USA, 2011; Volume 1, pp. 183–186. [Google Scholar]
- Zielinski, B.S.; Hara, T.J. The Neurobiology of Fish Olfaction. In Sensory Biology of Jawed Fishes; Kapoor, B.G., Hara, T.J., Eds.; Science Publishers: Enfields, UK, 2001; pp. 347–366. [Google Scholar]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar]
- Ngai, J.; Dowling, M.M.; Buck, L.; Axel, R.; Chess, A. The family of genes encoding odorant receptors in the channel catfish. Cell 1993, 72, 657–666. [Google Scholar]
- Niimura, Y.; Nei, M. Evolutionary dynamics of olfactory receptor genes in fishes and tetrapods. Proc. Natl. Acad. Sci. USA 2005, 102, 6039–6044. [Google Scholar]
- Hara, T.J. Feeding behaviour in some teleosts is triggered by single amino acids primarily through olfaction. J. Fish Biol 2006, 68, 810–825. [Google Scholar]
- Hara, T.J. Gustation. In Sensory Systems Neuroscience; Hara, T.J., Zielinski, B.S., Eds.; Fish Physiology Series 25; Academic Press: San Diego, CA, USA, 2006; pp. 45–96. [Google Scholar]
- Hara, T.J. Neurophysiology of Gustation. In Encyclopedia of Fish Physiology: From Genome to Environment; Farrell, A.P., Ed.; Academic Press: San Diego, CA, USA, 2011; Volume 1, pp. 218–226. [Google Scholar]
- Gilmour, K.M.; Perry, S.F. Branchial Chemoreceptor Regulation of Cardiorespiratory Function. In Sensory Systems Neuroscience; Hara, T.J., Zielinski, S.B., Eds.; Fish Physiology Series 25; Academic Press: San Diego, CA, USA, 2006; pp. 97–151. [Google Scholar]
- Yamashita, S.; Yamada, T.; Hara, T.J. Gustatory responses to feeding- and non-feeding-stimulant chemicals, with special emphasis on amino acids, in rainbow trout. J. Fish Biol 2006, 68, 783–800. [Google Scholar]
- Hara, T.J. Chemoreception. In The Physiology of Fishes; Evans, D.H., Ed.; CRC Press: Boca Raton, FL, USA, 1993; pp. 191–218. [Google Scholar]
- Hara, T.J. The Diversity of Chemical Stimulation in Fish Olfaction and Gustation. In Advances in Fish Biology; Singh, H.R., Ed.; Hindustan Publishing Corporation: Delhi, India, 1994; pp. 167–184. [Google Scholar]
- Sorensen, P.W.; Caprio, J. Chemoreception. In The Physiology of Fishes, 2nd ed; Evans, D.H., Ed.; CRC Press: Boca Raton, FL, USA, 1998; pp. 375–405. [Google Scholar]
- Caprio, J. Olfaction and taste in the channel catfish: An electrophysiological study of the responses to amino acids and derivatives. J. Comp. Physiol. A 1978, 123, 357–371. [Google Scholar]
- Speca, D.J.; Lin, D.M.; Sorensen, P.W.; Isacoff, E.Y.; Ngai, J.; Dittman, A.H. Functional identification of a goldfish odorant receptor. Neuron 1999, 23, 487–498. [Google Scholar]
- Laberge, F.; Hara, T.J. Behavioural and electrophysiological responses to F-prostaglandins, putative spawning pheromones, in three salmonids fishes. J. Fish Biol 2003, 62, 206–221. [Google Scholar]
- Laberge, F.; Hara, T.J. Non-oscillatory discharges of an F-prostaglandin responsive neuron population in the olfactory bulb-telencephalon transition area in lake whitefish. Neuroscience 2003, 116, 1089–1095. [Google Scholar]
- Laberge, F.; Hara, T.J. Electrophysiological demonstration of independent olfactory receptor types and associated neuronal responses in the trout olfactory bulb. Comp. Biochem. Physiol 2004, 137A, 397–408. [Google Scholar]
- Narahashi, T. Tetrodotoxin: A brief history. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci 2008, 84, 147–154. [Google Scholar]
- Yamamori, K.; Nakamura, M.; Matsui, T.; Hara, T.J. Gustatory responses to tetrodotoxin and saxitoxin in fish: A possible mechanism for avoiding marine toxins. Can. J. Fish. Aquat. Sci 1988, 45, 2182–2186. [Google Scholar]
- Ozeki, M.; Noma, A. The action of tetrodotoxin, procaine and acetylcholine on gustatory reception in frog and rat. Jpn. J. Physiol 1972, 22, 467–475. [Google Scholar]
- Kitada, Y.; Hara, T.J. Effects of diluted natural water and altered ionic environments on gustatory responses in rainbow trout (Oncorhynchus mykiss). J. Exp. Biol 1994, 186, 173–186. [Google Scholar]
- Yamashita, S.; Yamada, T.; Hara, T.J. Gustatory responses to feeding- and non-feeding-stimulant chemicals, with an emphasis on amino acids, in rainbow trout. J. Fish Biol 2006, 68, 783–800. [Google Scholar]
- Kinnamon, S.C.; Cummings, T.A. Chemosensory transduction mechanisms in taste. Annu. Rev. Physiol 1992, 54, 715–731. [Google Scholar]
- Avenet, P.; Lindemann, B. Patch-clamp study of isolated taste receptor cells of the frog. J. Membr. Biol 1987, 97, 223–240. [Google Scholar]




© 2011 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hara, T.J. Gustatory Detection of Tetrodotoxin and Saxitoxin, and Its Competitive Inhibition by Quinine and Strychnine in Freshwater Fishes. Mar. Drugs 2011, 9, 2283-2290. https://doi.org/10.3390/md9112283
Hara TJ. Gustatory Detection of Tetrodotoxin and Saxitoxin, and Its Competitive Inhibition by Quinine and Strychnine in Freshwater Fishes. Marine Drugs. 2011; 9(11):2283-2290. https://doi.org/10.3390/md9112283
Chicago/Turabian StyleHara, Toshiaki J. 2011. "Gustatory Detection of Tetrodotoxin and Saxitoxin, and Its Competitive Inhibition by Quinine and Strychnine in Freshwater Fishes" Marine Drugs 9, no. 11: 2283-2290. https://doi.org/10.3390/md9112283
APA StyleHara, T. J. (2011). Gustatory Detection of Tetrodotoxin and Saxitoxin, and Its Competitive Inhibition by Quinine and Strychnine in Freshwater Fishes. Marine Drugs, 9(11), 2283-2290. https://doi.org/10.3390/md9112283