Diversity and Antibacterial Activities of Fungi Derived from the Gorgonian Echinogorgia rebekka from the South China Sea



Abstract
:1. Introduction
2. Results and Discussions
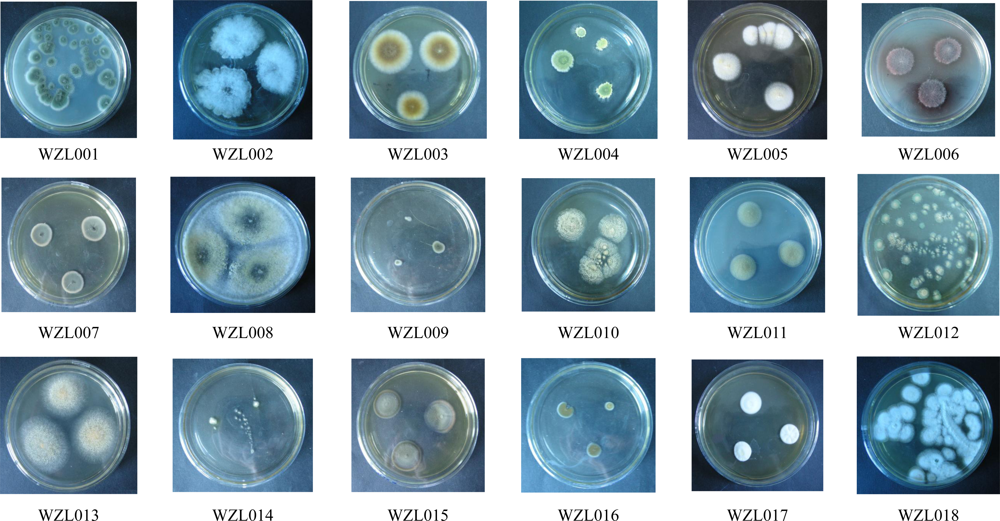
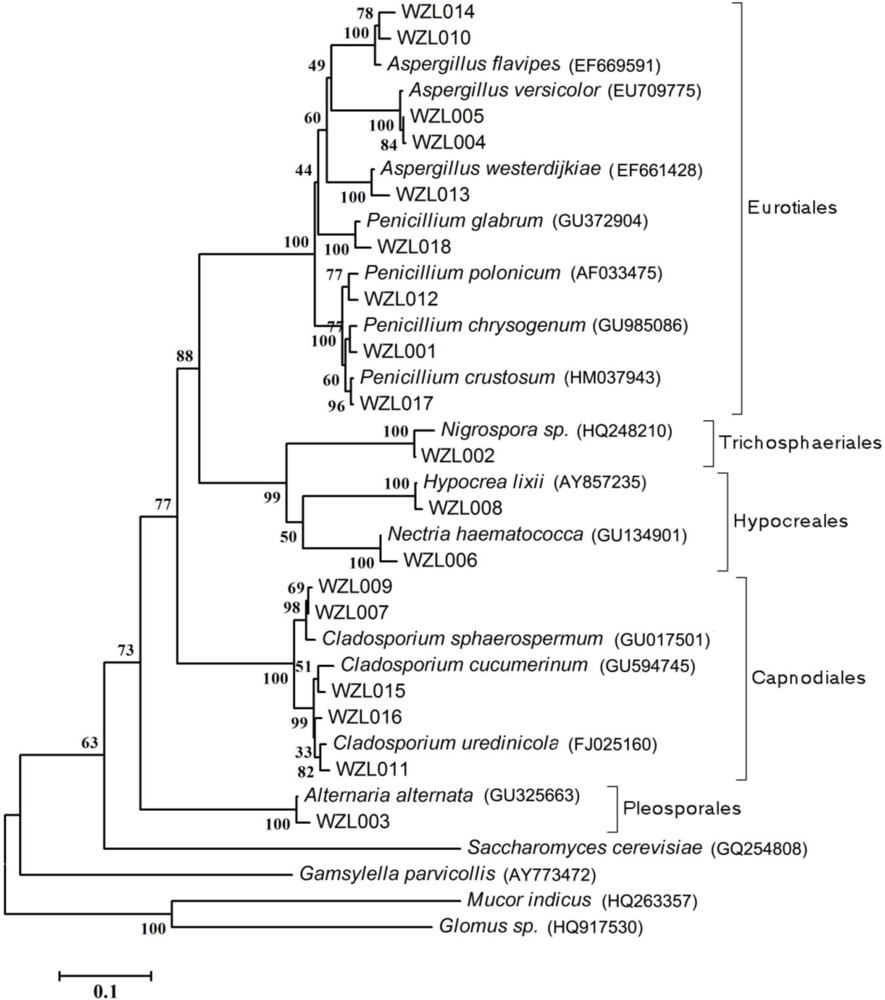
2.1. Diversity of Culturable Fungi Derived from the Gorgonian Echinogorgia rebekka
2.2. Antibacterial Activities of the Extracts from Fungal Broth and Mycelia
3. Experimental Section
3.1. Coral Materials
3.2. Fungi Isolation
3.3. Extraction of Genome DNA from Cultured Fungi
3.4. Amplification of Fungal ITS-rDNA Fragments
3.5. Sequence and Phylogenetic Analysis
3.6. Assay of Antibacterial Activity
4. Conclusions
Acknowledgments
- Samples Availability: Available from the authors.
References
- Balser, TC; McMahon, KD; Bart, D; Bronson, D; Coyle, DR; Craig, N. Bridging the gap between micro- and macro-scale perspectives on the role of microbial communities in global change ecology. Plant Soil 2006, 289, 59–70. [Google Scholar]
- Gutknecht, JLM; Goodman, RM; Balser, TC. Linking soil process and microbial ecology in fresh water wetland ecosystems. Plant Soil 2006, 289, 17–34. [Google Scholar]
- Schimel, J; Balser, T; Wallenstein, M. Microbial stress-response physiology and its implications for ecosystem function. Ecology 2007, 88, 1386–1394. [Google Scholar]
- Ainsworth, TD; Thurber, RV; Gates, RD. The future of coral reefs: A microbial perspective. Trends Ecol. Evol 2009, 25, 233–240. [Google Scholar]
- Blunt, JW; Copp, BR; Hu, WP; Munro, MHG; Northcote, PT; Prinsep, MR. Marine natural products. Nat. Prod. Rep 2008, 25, 35–94. [Google Scholar]
- König, GM; Kehraus, S; Seiber, SF; Abdel-Lateff, A; Muller, D. Natural products from marine organisms and their associated microbes. Chem. Biochem 2006, 7, 229–238. [Google Scholar]
- Jensen, PR; Fenical, W. Secondary Metabolites from Marine Fungi. In Fungi in Marine Environments; Hyde, KD, Ed.; Fungal Diversity Press: Hong Kong, China, 2002; pp. 293–315. [Google Scholar]
- Li, Z. Advances in marine microbial symbionts in the China Sea and related pharmaceutical metabolites. Mar. Drugs 2009, 7, 113–129. [Google Scholar]
- Proksch, P; Edrada-Ebel, R; Ebel, R. Drugs from the sea—opportunities obstacles. Mar. Drugs 2003, 1, 5–17. [Google Scholar]
- Bhatnagar, I; Kim, S-K. Immense essence of excellence: Marine microbial bioactive compounds. Mar. Drugs 2010, 8, 2673–2701. [Google Scholar]
- Radjasa, OK. Marine invertebrate-associated bacteria in coral reef ecosystems as a new source of bioactive compounds. J. Coast. Dev 2004, 7, 65–70. [Google Scholar]
- Trisuwan, K; Rukachaisirikul, V; Sukpondma, Y; Preedanon, S; Phongpaichit, S; Rungjindamai, N; Sakayaroj, J. Epoxydons and a pyrone from the marine-derived fungus Nigrospora sp. PSU-F5. J. Nat. Prod 2008, 71, 1323–1326. [Google Scholar]
- Arvanitidou, M; Kanellou, K; Constantinidis, T; Katsouyannopoulos, V. Higher prevalence of Alternaria spp. in marine and river waters than in potable samples. Microbiol. Res 2000, 155, 49–51. [Google Scholar]
- Koh, LL; Tan, TK; Chou, LM; Goh, NKC. Fungi Associated with Gorgonians in Singapore. Proceedings of the 9th International Coral Reef Symposium, Bali, Indonesia, 23–27 October 2000.
- Baker, PW; Kennedy, J; Dobson, ADW; Marchesi, JR. Phylogenetic diversity and antimicrobial activities of fungi associated with Haliclona simulans isolated from Irish coastal waters. Mar. Biotechnol 2009, 11, 540–547. [Google Scholar]
- Da Silva, M; Passarini, MRZ; Bonugli, RC; Sette, LD. Cnidarian-derived filamentous fungi from Brazil: Isolation, characterization and RBBR decolourization screening. Environ. Technol 2008, 29, 1331–1339. [Google Scholar]
- Holler, U; Wright, AD; Matthee, GF; Konig, GM; Draeger, D; Aust, HJ. Fungi from marine sponges: Diversity, biological activity and secondary metabolites. Mycol. Res 2000, 104, 1354–1365. [Google Scholar]
- Bugni, TS; Ireland, CM. Marine-derived fungi: A chemically and biologically diverse group of microorganisms. Nat. Prod. Rep 2004, 21, 143–163. [Google Scholar]
- Konings, WN; Poolman, B; Driessen, AM. Can the excretion of metabolites by bacteria be manipulated? FEMS Microbiol. Lett 1992, 2, 93–108. [Google Scholar]
- Li, Q; Wang, G. Diversity of fungal isolates from three Hawaiian marine sponges. Microbiol. Res 2009, 164, 233–241. [Google Scholar]
- Wei, JC. The Identification Manual of Fungi; Shanghai Science and Technology Press: Shanghai, China, 1979. [Google Scholar]
- Fredricks, DN; Smith, C; Meier, A. Comparison of six DNA extraction methods for recovery of fungal DNA as assessed by quantitative PCR. J. Clin. Microbiol 2005, 43, 5122–5128. [Google Scholar]
- White, TJ; Bruns, T; Lee, S; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Application; Innis, MA, Gelfand, DH, Sninsky, JJ, White, TJ, Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- National Center for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov (accessed on 10 June 2011).
- Thompson, JD; Gibson, TJ; Plewniak, F; Jeanmougin, F; Higgins, DG. The ClustalX windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 1997, 25, 4876–4882. [Google Scholar]
- Hall, TA. BioEdit: A user friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser 1999, 41, 95–98. [Google Scholar]
- Tamura, K; Peterson, D; Peterson, N; Stecher, G; Nei, M; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 2011. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol 1980, 16, 111–120. [Google Scholar]
- Gottler, LM; Ramamoorthy, A. Structure, membrane orientation, mechanism, and function of pexiganan—A highly potent antimicrobial peptide designed from magainin. Biochim. Biophys Acta 2009, 1788, 1680–1686. [Google Scholar]
- Marsh, EN; Buer, BC; Ramamoorthy, A. Fluorine—A new element in the design of membrane-active peptides. Mol. BioSyst 2009, 5, 1143–1147. [Google Scholar]



{kind=link}
{kind=link}
| Strain | Morphological Identification | Seq. Length (bp) | Related Strain (BLAST) | Access. No. | Similarity (%) | Overlap (bp) |
|---|---|---|---|---|---|---|
| WZL001 | Penicillium sp. | 559 | P. chrysogenum | GU985086 | 98 | 554 |
| WZL002 | Nigrospora sp. | 528 | Nigrospora sp. | HQ248210 | 99 | 513 |
| WZL003 | Alternaria sp. | 537 | Alternaria alternata | GU325663 | 100 | 517 |
| WZL004 | Aspergillus sp. | 545 | Aspergillus versicolor | EU709775 | 99 | 537 |
| WZL005 | Aspergillus sp. | 533 | Aspergillus sp. | HQ637366 | 98 | 527 |
| WZL006 | Nectria sp. | 543 | N. haematococca | GU134901 | 98 | 521 |
| WZL007 | Cladosporium sp. | 527 | Cladosporium sp. | JF819134 | 100 | 509 |
| WZL008 | Hypocrea sp. | 594 | H. lixii | AY857235 | 99 | 577 |
| WZL009 | Cladosporium sp. | 529 | C. sphaerospermum | JF793522 | 99 | 518 |
| WZL010 | Aspergillus sp. | 553 | A. flavipes | EF669591 | 98 | 539 |
| WZL011 | Cladosporium sp. | 526 | C. uredinicola | FJ025160 | 99 | 509 |
| WZL012 | Penicillium sp. | 558 | P. polonicum | AF033475 | 98 | 545 |
| WZL013 | Aspergillus sp. | 561 | A. westerdijkiae | EF661428 | 98 | 548 |
| WZL014 | Aspergillus sp. | 565 | Aspergillus sp. | HQ731625 | 98 | 551 |
| WZL015 | Cladosporium sp. | 529 | C. cladosporioides | GU932679 | 99 | 518 |
| WZL016 | Cladosporium sp. | 518 | C. cucumerinum | GU594745 | 99 | 509 |
| WZL017 | Penicillium sp. | 563 | P. crustosum | HM037943 | 99 | 559 |
| WZL018 | Penicillium sp. | 556 | P. glabrum | GU372904 | 99 | 544 |
| Fungal Strain | B. subtilis | S. aureus | E. coli | M. tetragenus | V. anguillarum | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Class | Order | Genus | Strain | Broth | Mycelia | Broth | Mycelia | Broth | Mycelia | Broth | Mycelia | Broth | Mycelia |
| Eurotiomycetes | Eurotiales | Penicillium | WZL001 | − | − | − | − | − | − | ++ | − | − | − |
| WZL012 | − | − | − | ++ | − | − | − | − | − | − | |||
| WZL017 | − | +++ | ++ | − | − | − | ++ | − | − | − | |||
| WZL018 | − | − | − | − | − | − | − | ++++ | − | − | |||
| Aspergillus | WZL004 | − | − | ++ | ++ | − | − | − | − | − | − | ||
| WZL005 | − | − | − | ++ | − | − | − | − | − | − | |||
| WZL010 | − | − | ++ | ++ | − | − | − | − | − | − | |||
| WZL013 | − | − | − | − | − | − | ++ | − | − | − | |||
| WZL014 | − | − | ++ | − | − | − | − | − | − | − | |||
| Dothideomycetes | Pleosporales | Alternaria | WZL003 | − | − | − | + | − | − | − | − | − | − |
| Capnodiales | Cladosporium | WZL007 | − | − | ++ | − | − | − | + | − | − | − | |
| WZL009 | − | ++ | +++ | ++ | − | − | ++ | − | − | − | |||
| WZL011 | − | − | ++ | ++ | − | − | − | − | − | − | |||
| WZL015 | − | +++ | ++ | − | − | − | ++ | − | − | − | |||
| WZL016 | − | − | ++ | − | − | − | − | − | − | − | |||
| Sordariomycetes | Trichosphaeriales | Nigrospora | WZL002 | − | − | − | − | − | − | − | ++ | − | − |
| Hypocreales | Nectria | WZL006 | − | − | − | ++ | − | − | − | − | − | − | |
| Hypocrea | WZL008 | − | − | − | − | ++ | − | ++ | − | ++ | − | ||
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, Y.-N.; Shao, C.-L.; Zheng, C.-J.; Chen, Y.-Y.; Wang, C.-Y. Diversity and Antibacterial Activities of Fungi Derived from the Gorgonian Echinogorgia rebekka from the South China Sea. Mar. Drugs 2011, 9, 1379-1390. https://doi.org/10.3390/md9081379
Wang Y-N, Shao C-L, Zheng C-J, Chen Y-Y, Wang C-Y. Diversity and Antibacterial Activities of Fungi Derived from the Gorgonian Echinogorgia rebekka from the South China Sea. Marine Drugs. 2011; 9(8):1379-1390. https://doi.org/10.3390/md9081379
Chicago/Turabian StyleWang, Ya-Nan, Chang-Lun Shao, Cai-Juan Zheng, Yi-Yan Chen, and Chang-Yun Wang. 2011. "Diversity and Antibacterial Activities of Fungi Derived from the Gorgonian Echinogorgia rebekka from the South China Sea" Marine Drugs 9, no. 8: 1379-1390. https://doi.org/10.3390/md9081379
APA StyleWang, Y. -N., Shao, C. -L., Zheng, C. -J., Chen, Y. -Y., & Wang, C. -Y. (2011). Diversity and Antibacterial Activities of Fungi Derived from the Gorgonian Echinogorgia rebekka from the South China Sea. Marine Drugs, 9(8), 1379-1390. https://doi.org/10.3390/md9081379