Climate Change and West Nile Virus in a Highly Endemic Region of North America


Abstract
:1. Introduction

2. Materials and Methods
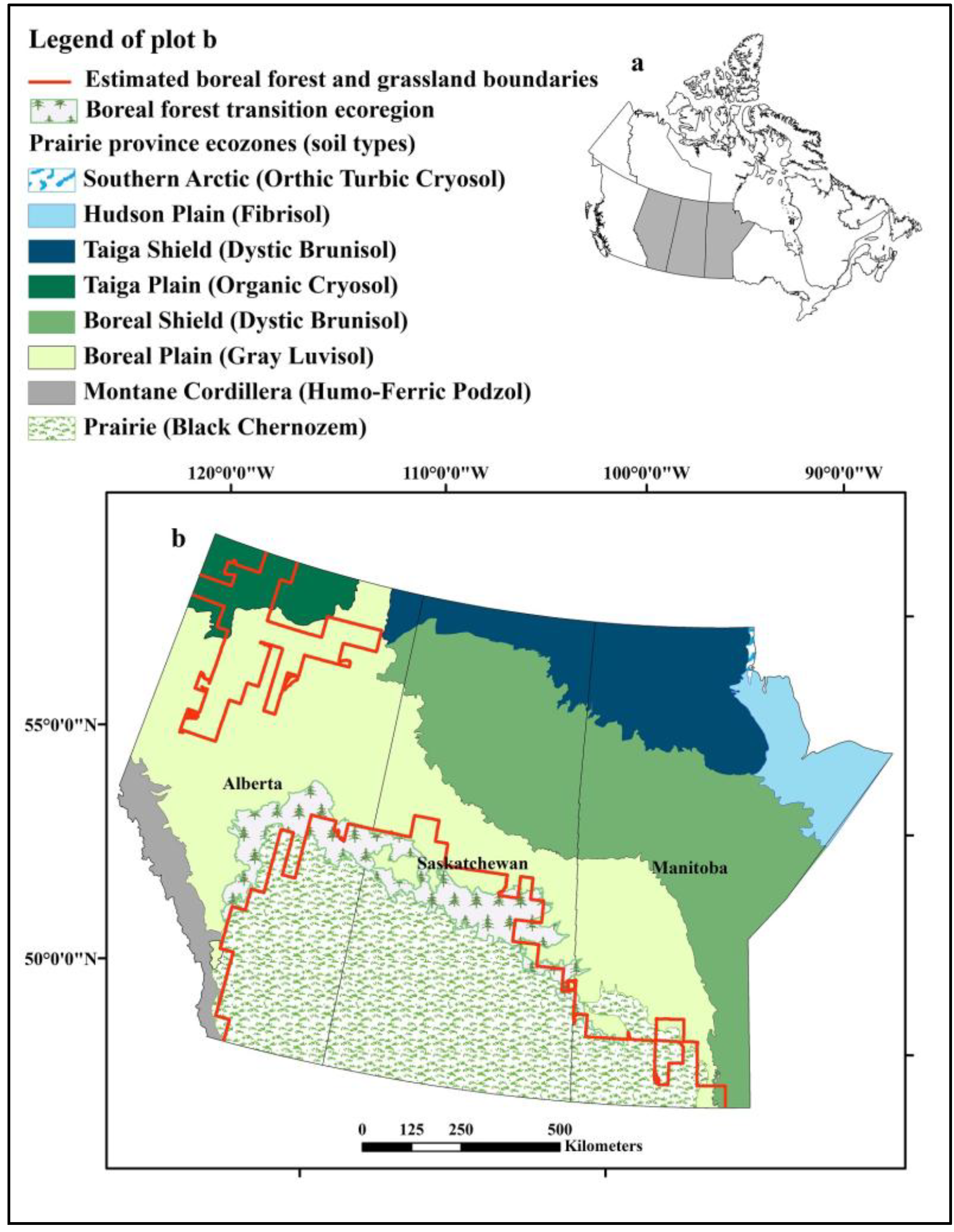
2.1. Selection of Three Climate Change Outcome Scenarios

{kind=link}
{kind=link}
{kind=link}
| General circulation models | SRA2 | SRA1B | SRB1 | Resolution Latitude (°) | Resolution Longitude (°) |
|---|---|---|---|---|---|
| BCCR-BCM2.0 | 1 | 1 | 1 | 1.9 | 1.9 |
| CGCM3.1_T47 | 5 | 5 | 5 | 2.8 | 2.8 |
| CGCM3.1_T63 | 1 | 1 | 1 | 1.9 | 1.9 |
| CNRM-CM3 | 1 | 1 | 1 | 1.9 | 1.9 |
| CSIROMk3.0 | 1 | 1 | 1 | 1.9 | 1.9 |
| CSIROMk3.5 | 1 | 1 | 1 | 1.9 | 1.9 |
| ECHAM5 | 3 | 4 | 3 | 1.9 | 1.9 |
| ECHO-G | 3 | 3 | 3 | 3.9 | 3.9 |
| FGOALS | 3 | 3 | 2.8 | 2.8 | |
| GFDL-CM2.0 | 1 | 1 | 1 | 2.0 | 2.5 |
| GFDL-CM2.1 | 1 | 1 | 1 | 2.0 | 2.5 |
| GISS-AOM | 2 | 2 | 3.0 | 4.0 | |
| GISS-EH | 3 | 4.0 | 5.0 | ||
| GISS-ER (run number) 1 | 1 (1) | 2 (2, 4) | 1 (1) | 4.0 | 5.0 |
| INGV-SXG | 1 | 1 | 1.1 | 1.1 | |
| INM-CM3.0 | 1 | 1 | 1 | 4.0 | 5.0 |
| IPSL-CM4 | 1 | 1 | 1 | 2.5 | 3.75 |
| MIROC3.2-hires | 1 | 1 | 1.1 | 1.1 | |
| MIROC3.2-medres | 3 | 3 | 3 (run 2) | 2.8 | 2.8 |
| CGCM2.3.2 | 5 | 5 | 5 | 2.8 | 2.8 |
| NCAR-CCSM (run numbers) | 4 (1–4) | 7 (1–3, 5–7, 9) | 8 (1–7, 9) | 1.4 | 1.4 |
| NCAR-PCM | 4 | 4 | 3 (run 2) | 2.8 | 2.8 |
| UKMO-HadCM3 | 1 | 1 | 1 | 2.5 | 3.75 |
| UKMO-HadGEM1 | 1 | 1 | 1.3 | 1.9 | |
| Total experiments | 40 | 54 | 48 |
2.2. Models for Cx. tarsalis Abundance and WNV Infection Rate
| Variables (unit) | Cx. tarsalis abundance | WNV infection rate | ||
|---|---|---|---|---|
| Coefficient | 95% CI | Coefficient | 95% CI | |
| Intercept | −3.48 | −4.05 to −2.91 | −2.26 | −4.47 to −0.05 |
| Cx. tarsalis abundance (log(y+1)) | 0.55 | 0.31 to 0.79 | ||
| Climate variables | ||||
| Monthly mean temperature (1 °C) | 0.22 | 0.2 to 0.25 | ||
| 1 month lagged temperature (1 °C) | 0.07 | 0.05 to 0.09 | 0.32 | 0.22 to 0.41 |
| 3 months total of monthly mean degree days (dd) | −0.10 | −0.2 to −0.01 | ||
| Monthly total precipitation (1 mm) | 0.0033 | 0.002 to 0.005 | ||
| 1 month lagged precipitation (1 mm) | 0.0042 | 0.003 to 0.005 | −0.27 | −0.36 to −0.18 |
| 2 month lagged precipitation (1 mm) | 0.0033 | 0.002 to 0.004 | ||
| 3 months total precipitation (1 mm) | −0.05 | −0.08 to −0.02 | ||
2.3. Modeling Grassland Distribution
- For Tmean > 10 °C: PET = 93 D exp(A/9300)
- For 10 °C ≥ Tmean > –5 °C: PET = (6.2Tmean + 31) D exp(A/9300)
- For Tmean ≤ –5 °C: PET = 0
- Tmean = mean monthly temperature (unit: °C)
- D = vapour pressure deficit (unit: KPa)
- A = altitude (unit: meter)
- D = 0.5 (eTmax + eTmin) – eTdew
- eTmax = saturated vapor pressure at the maximum monthly mean temperature
- eTmin = saturated vapor pressure at the minimum monthly mean temperature
- eTdew = saturated vapor pressure at the dew point temperature.
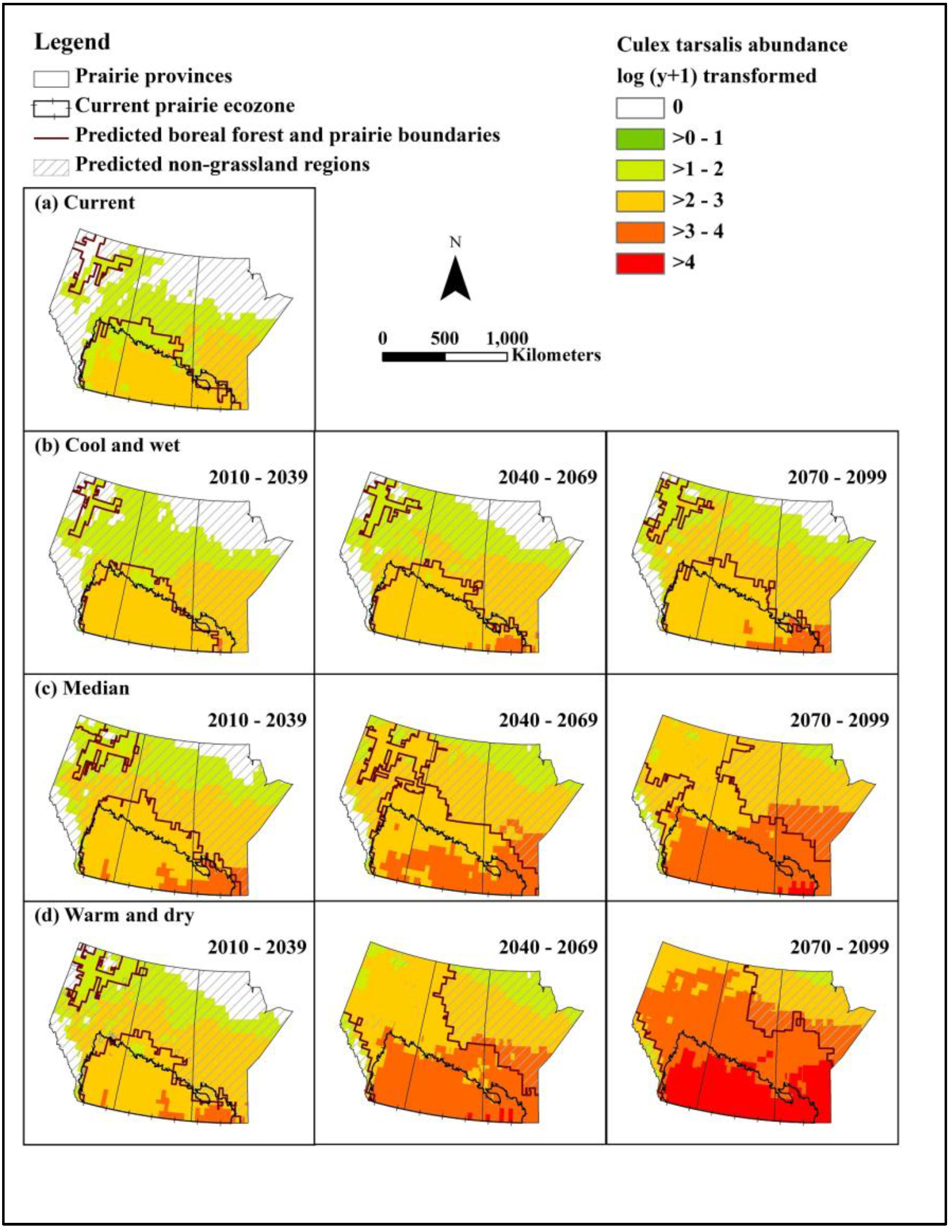
2.4. Maps of Current and Future Distribution
2.4.1. Maps of Cx. tarsalis Abundance
2.4.2. Maps of WNV Infection Rate in Cx. tarsalis
3. Results
3.1. Selection of Three Climate Change Outcome Scenarios
| Experiments and time slices | Emissions Scenarios | Outcome Scenarios | Change in annual total precipitation (SD); mm | Change in mean annual temperature (SD); °C |
|---|---|---|---|---|
| 2010–2039 (2020s) | ||||
| NCAR-PCM run 2 | B1 | Cool, wet | 22.4 (8.9) | 1.14 (0.27) |
| MIMR | B1 | Median | 25.8 (16.3) | 1.63 (0.18) |
| UKMO-HadGEM1 run 1 | A2 | Warm, dry | 44.2 (16.9) | 1.65 (0.40) |
| 2040–2069 (2050s) | ||||
| NCAR-PCM run 2 | B1 | Cool, wet | 41.1 (12.8) | 1.77 (0.28) |
| MIMR | B1 | Median | 37.9 (28.1) | 3.04 (0.11) |
| UKMO-HadGEM1 run 1 | A2 | Warm, dry | 21.3 (22.9) | 4.03 (0.39) |
| 2070–2099 (2080s) | ||||
| NCAR-PCM run 2 | B1 | Cool, wet | 52.3 (12.6) | 2.42 (0.33) |
| MIMR | B1 | Median | 46.4 (31.2) | 4.24 (0.19) |
| UKMO-HadGEM1 run 1 | A2 | Warm, dry | 29.4 (31.4) | 6.80 (0.50) |
3.2. Grassland Distribution

3.3. Culex tarsalis Abundance and Distribution
3.4. WNV Distribution and Infection Rate in Cx. tarsalis
| Outcome scenarios and time slices | Distribution area 2 | Area expansion 3 | Fold change 4 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Grassland | Cx. tarsalis | WNV | Grassland | Cx. tarsalis | WNV | Grassland | Cx. tarsalis | WNV | ||||
| Current 1 | 607,018 | 566,506 | 539,877 | |||||||||
| 2010–2039 | ||||||||||||
| Cool, wet | 543,502 | 536,042 | 516,619 | −23,004 | −30,464 | −23,258 | 0.90 | 0.95 | 0.96 | |||
| Median | 727,509 | 727,029 | 711,578 | 120,491 | 160,523 | 171,701 | 1.20 | 1.28 | 1.32 | |||
| Warm, dry | 624,783 | 617,767 | 586,466 | 17,765 | 51,261 | 46,589 | 1.03 | 1.09 | 1.09 | |||
| 2040–2069 | ||||||||||||
| Cool, wet | 599,256 | 599,256 | 582,998 | −7,762 | 32,750 | 43,121 | 0.99 | 1.06 | 1.08 | |||
| Median | 905,701 | 905,701 | 872,337 | 298,683 | 339,195 | 332,460 | 1.49 | 1.60 | 1.62 | |||
| Warm, dry | 1,198,242 | 1,198,242 | 1,151,876 | 591,224 | 631,736 | 611,999 | 1.97 | 2.12 | 2.13 | |||
| 2070–2099 | ||||||||||||
| Cool, wet | 664,150 | 664,150 | 657,321 | 57,132 | 97,644 | 117,444 | 1.09 | 1.17 | 1.22 | |||
| Median | 1,082,641 | 1,082,641 | 1,036,084 | 475,623 | 516,135 | 496,207 | 1.78 | 1.91 | 1.92 | |||
| Warm, dry | 1,449,128 | 1,449,128 | 1,263,070 | 842110 | 882,622 | 723,193 | 2.39 | 2.56 | 2.34 | |||
| Outcome scenarios | May | June | July | August | September | |||
|---|---|---|---|---|---|---|---|---|
| Abun (SD) | Abun (SD) | Fold change 1 | Abun (SD) | Fold change | Abun (SD) | Fold change | Abun (SD) | |
| Current | None | 1.22 (0.33) | 2.26 (0.32) | 2.20 (0.32) | None | |||
| 2010–2039 | ||||||||
| Cool, wet | None | 1.48 (0.32) | 1.21 | 2.56 (0.29) | 1.13 | 2.43 (0.31) | 1.10 | None |
| Median | None | 1.57 (0.31) | 1.29 | 2.68 (0.32) | 1.19 | 2.75 (0.30) | 1.25 | 0.28 (0.60) |
| Warm, dry | None | 1.71 (0.31) | 1.40 | 2.62 (0.28) | 1.16 | 2.65 (0.30) | 1.20 | 0.08 (0.33) |
| 2040–2069 | ||||||||
| Cool, wet | None | 1.57 (0.29) | 1.29 | 2.66 (0.26) | 1.18 | 2.54 (0.31) | 1.15 | 0.05 (0.28) |
| Median | 0.05 (0.16) | 1.97 (0.26) | 1.61 | 3.03 (0.29) | 1.34 | 3.02 (0.27) | 1.37 | 1.11 (0.76) |
| Warm, dry | 0.37 (0.39) | 2.30 (0.32) | 1.89 | 3.34 (0.34) | 1.48 | 3.49 (0.32) | 1.59 | 1.90 (0.50) |
| 2070–2099 | ||||||||
| Cool, wet | 0.02 (0.09) | 1.72 (0.33) | 1.41 | 2.75 (0.28) | 1.22 | 2.69 (0.30) | 1.22 | 0.28 (0.63) |
| Median | 0.49 (0.36) | 2.26 (0.27) | 1.85 | 3.32 (0.32) | 1.47 | 3.39 (0.33) | 1.54 | 1.83 (0.36) |
| Warm, dry | 1.18 (0.27) | 3.04 (0.31) | 2.49 | 4.27 (0.32) | 1.89 | 4.33 (0.31) | 1.97 | 2.87 (0.26) |
| Outcome scenarios | May | June | July | August | September | |||
|---|---|---|---|---|---|---|---|---|
| IR(SD) | IR (SD) | Fold change 1 | IR (SD) | Fold change | IR (SD) | Fold change | IR (SD) | |
| Current | None | 0.54 (0.25) | 1.33 (0.45) | 2.23 (0.94) | None | |||
| 2010–2039 | ||||||||
| Cool, wet | None | 0.51 (0.16) | 0.94 | 1.14 (0.42) | 0.86 | 2.88 (1.27) | 1.29 | None |
| Median | None | 0.86 (0.25) | 1.59 | 2.02 (0.96) | 1.52 | 5.17 (2.64) | 2.32 | 0.44 (1.32) |
| Warm, dry | None | 0.67 (0.22) | 1.24 | 1.67 (0.71) | 1.26 | 2.84 (1.42) | 1.27 | 0.40 (0.99) |
| 2040–2069 | ||||||||
| Cool, wet | None | 0.72 (0.23) | 1.33 | 1.62 (0.66) | 1.22 | 3.20 (1.47) | 1.43 | 0.06 (0.41) |
| Median | 0.02 (0.09) | 1.91 (0.71) | 3.54 | 4.87 (1.91) | 3.66 | 17.91 (10.0) | 8.03 | 4.32 (4.63) |
| Warm, dry | 0.17 (0.21) | 1.37 (0.46) | 2.54 | 5.91 (3.08) | 4.44 | 18.08 (9.98) | 8.11 | 10.18 (6.93) |
| 2070–2099 | ||||||||
| Cool, wet | 0.004 (0.03) | 1.01 (0.31) | 1.87 | 1.91 (0.87) | 1.44 | 3.74 (1.76) | 1.68 | 0.28 (0.87) |
| Median | 0.001 (0.001) | 1.79 (0.64) | 3.31 | 5.53 (2.57) | 4.16 | 19.95 (11.81) | 8.95 | 9.44 (7.06) |
| Warm, dry | 0.61 (0.22) | 2.70 (0.93) | 5.00 | 17.55 (6.70) | 13.20 | 61.21 (27.78) | 27.45 | 30.89 (11.66) |

4. Discussion
5. Conclusions
Acknowledgements
Conflict of Interest
References
- Gubler, D.J.; Reiter, P.; Ebi, K.L.; Yap, W.; Nasci, R.; Patz, J.A. Climate variability and change in the United States: Potential impacts on vector- and rodent-borne diseases. Environ. Health Perspect. 2001, 109, 223–233. [Google Scholar] [CrossRef]
- Hunter, P.R. Climate change and waterborne and vector-borne disease. J. Appl. Microbiol. 2003, 94, 37–46. [Google Scholar] [CrossRef]
- Kovats, R.S.; Campbell-Lendrum, D.H.; McMichel, A.J.; Woodward, A.; Cox, J.S.H. Early effects of climate change: Do they include changes in vector-borne disease? Philos. Trans. R. Soc. Lond. B: Biol. Sci. 2001, 356, 1057–1068. [Google Scholar] [CrossRef]
- Greer, A.; Ng, V.; Fisman, D. Climate change and infectious diseases in North America: The road ahead. Can. Med. Assoc. J. 2008, 178, 715–722. [Google Scholar]
- Chan, N.Y.; Ebi, K.L.; Smith, F.; Wilson, T.F.; Smith, A.E. An integrated assessment framework for climate change and infectious diseases. Environ. Health Perspect. 1999, 107, 329–337. [Google Scholar]
- Nash, D.; Mostashari, F.; Fine, A.; Miller, J.; O’Leary, D.; Murray, K.; Huang, A.; Rosenberg, A.; Greenberg, A.; Sherman, M.; et al. The outbreak of West Nile virus infection in the New York city area in 1999. N. Engl. J. Med. 2001, 344, 1807–1814. [Google Scholar] [CrossRef]
- Public Health Agency of Canada. West Nile MONITOR–2007 Human Surveillance. Available online: http://www.phac-aspc.gc.ca/WNV-VNW/mon-hmnsurv—2007.eng.php (accessed on 23 February 2013).
- Curry, P. Saskatchewan mosquitoes and West Nile virus. Blue Jay 2004, 62, 104–111. [Google Scholar]
- Yiannakoulias, N.W.; Schopflocher, D.P.; Svenson, L.W. Modelling geographic variations in West Nile virus. Can. J. Public Health. 2006, 97, 374–378. [Google Scholar]
- Turell, M.J.; Dohm, D.J.; Sardelis, M.R.; O’Guinn, M.L.; Andreadis, T.G.; Blow, J.A. An update on the potential of North American mosquitoes (Diptera: Culicidae) to transmit West Nile virus. J. Med. Entomol. 2005, 42, 57–62. [Google Scholar] [CrossRef]
- Waeckerlin, R.; Swann, J.; Elkin, B.; Zuliani, A.; Cork, S. Flavi and Bunyavirus Mosquito Vector Distribution in North Western Canada. In Proceedings of 15th International Congress on Infectious Diseases, Bangkok, Thailand, 13–16 June 2012; International Society for Infectious Diseases: Brookline, MA, USA, 2012. [Google Scholar]
- Jenkins, D.W. Bionomics of Culex tarsalis in relation to Western equine encephalomyelitis. Am. J. Trop. Med. Hyg. 1950, 30 (Suppl.), 909–916. [Google Scholar]
- Epp, T.Y.; Waldner, C.L.; Berke, O. Predicting geographical human risk of West Nile virus—Saskatchewan, 2003 and 2007. Can. J. Public Health. 2009, 100, 344–348. [Google Scholar]
- Chuang, T.W.; Hildreth, M.B.; Vanroekel, D.L.; Wimberly, M.C. Weather and land cover influences on mosquito populations in Sioux Falls, South Dakota. J. Med. Entomol. 2011, 48, 669–679. [Google Scholar] [CrossRef]
- Beehler, J.W.; Mulla, M.S. Effects of organic enrichment on temporal distribution and abundance of culicine egg rafts. J. Am. Mosq. Control Assoc. 1995, 11, 167–171. [Google Scholar]
- Zou, L.; Miller, S.N.; Schmidtmann, E.T. Mosquito larval habitat mapping using remote sensing and GIS: Implications of coalbed methane development and West Nile virus. J. Med. Entomol. 2006, 43, 1034–1041. [Google Scholar] [CrossRef]
- Wimberly, M.C.; Hildreth, M.B.; Boyte, S.P.; Lindquist, E.; Kightlinger, L. Ecological niche of the 2003 West Nile virus epidemic in the Northern Great Plains of the United States. PLoS ONE 2008, 3. [Google Scholar] [CrossRef]
- Chuang, T.-W.; Hockett, C.W.; Kightlinger, L.; Wimberly, M.C. Landscape-level spatial patterns of West Nile virus risk in the Northern Great Plains. Am. J. Trop. Med. Hyg. 2012, 86, 724–731. [Google Scholar] [CrossRef]
- Goddard, L.B.; Roth, A.E.; Reisen, W.K.; Scott, T.W. Vertical transmission of West Nile virus by three California Culex (Diptera: Culicidae) species. J. Med. Entomol. 2003, 40, 743–746. [Google Scholar] [CrossRef]
- Kent, R.; Juliusson, L.; Weissmann, M.; Evans, S.; Komar, N. Seasonal blood-feeding behavior of Culex tarsalis (Diptera: Culicidae) in Weld county, Colorado, 2007. J. Med. Entomol. 2009, 46, 380–390. [Google Scholar] [CrossRef]
- Thiemann, T.; Lemenager, D.; Kluh, S.; Carroll, B.; Lothrop, H.; Reisen, W. Spatial variation in host feeding patterns of Culex tarsalis and the Culex pipiens Complex (Diptera: Culicidae) in California. J. Med. Entomol. 2012, 49, 903–916. [Google Scholar] [CrossRef]
- Lee, J.H.; Hassan, H.; Hill, G.; Cupp, E.W.; Higazi, T.B.; Mitchell, C.J.; Godsey, M.S., Jr; Unnasch, T.R. Identification of mosquito avian-derived blood meals by polymerase chain reaction-heteroduplex analysis. Am. J. Trop. Med. Hyg. 2002, 66, 599–604. [Google Scholar]
- Artsob, H. Arthropod-borne disease in Canada: A clinician’s perspective from the ‘Cold Zone’. Paediatr. Child Health 2000, 5, 206–213. [Google Scholar]
- Hongoh, V.; Berrang-Ford, L.; Ogden, N.H.; Lindsay, R.; Scott, M.E.; Artsob, H. A review of environmental determinants and risk factors for avian-associated mosquito arboviruses in Canada. Biodiversity 2009, 10, 83–91. [Google Scholar] [CrossRef]
- Lafferty, K.D. The ecology of climate change and infectious diseases. Ecology 2009, 90, 888–900. [Google Scholar] [CrossRef]
- Hogg, E.H.; Hurdle, P. The aspen parkland in western Canada: A dry-climate analogue for the future boreal forest? Water. Air. Soil Pollut. 1995, 82, 391–400. [Google Scholar] [CrossRef]
- Thorpe, J. Vulnerability of Prairie Grasslands to Climate Change; Saskatchewan Research Council: Saskatoon, SK, Canada, 2011. [Google Scholar]
- Rizzo, B.; Wiken, E. Assessing the sensitivity of Canada’s ecosystems to climatic change. Clim. Change 1992, 21, 37–55. [Google Scholar] [CrossRef]
- Hogg, E.H. Climate and the southern limit of the western Canadian boreal forest. Can. J. For. Res. 1994, 24, 1835–1845. [Google Scholar] [CrossRef]
- General Guidelines on the Use of Scenario Data for Climate Impact and Adaptation Assessment, Version 2; IPCC-TGICA: Geneva, Switzerland, 2007.
- Randall, D.A.; Wood, R.A.; Bony, S.; Colman, R.; Fichefet, T.; Fyfe, J.; Kattsov, V.; Pitman, A.; Shukla, J.; Srinivasan, J. Climate Models and Their Evaluation. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 591–648. [Google Scholar]
- Trawinski, P.; Mackay, D. Spatial autocorrelation of West Nile virus vector mosquito abundance in a seasonally wet suburban environment. J. Geogr. Syst. 2009, 11, 67–87. [Google Scholar] [CrossRef]
- Biggerstaff, B. PooledInfRate, Version 3.0: A Microsoft Excel Add-In to Compute Prevalence Estimates from Pooled Samples; Centers for Disease Control and Prevention: Fort Collins, CO, USA, 2006. [Google Scholar]
- Chiang, C.L.; Reeves, W.C. Statistical estimation of virus infection rates in mosquito vector populations. Am. J. Epidemiol. 1962, 75, 377–391. [Google Scholar]
- Hogg, E.H. Temporal scaling of moisture and the forest-grassland boundary in western Canada. Agric. For. Meteorol. 1997, 84, 115–122. [Google Scholar] [CrossRef]
- Reisen, W.K. Effect of temperature on Culex tarsalis (Diptera: Culicidae) from the Coachella and San Joaquin valleys of California. J. Med. Entomol. 1995, 32, 636–645. [Google Scholar]
- Reisen, W.K.; Fang, Y.; Martinez, V.M. Effects of temperature on the transmission of West Nile virus by Culex tarsalis (Diptera: Culicidae). J. Med. Entomol. 2006, 43, 309–317. [Google Scholar] [CrossRef]
- Schrag, A.; Konrad, S.; Miller, S.; Walker, B.; Forrest, S. Climate-change impacts on sagebrush habitat and West Nile virus transmission risk and conservation implications for greater sage-grouse. GeoJournal 2011, 76, 561–575. [Google Scholar] [CrossRef]
- Reisen, W.; Meyer, R.; Presser, S.; Hardy, J. Effect of temperature on the transmission of western equine encephalomyelitis and St. Louis encephalitis viruses by Culex tarsalis (Diptera: Culicidae). J. Med. Entomol. 1993, 30, 151–160. [Google Scholar]
- Ostfeld, R.S. Climate change and the distribution and intensity of infectious diseases. Ecology 2009, 90, 903–905. [Google Scholar] [CrossRef]
- Brownstein, J.; Holford, T.; Fish, D. Effect of climate change on Lyme disease risk in North America. EcoHealth 2005, 2, 38–46. [Google Scholar] [CrossRef]
- Henn, J.B.; Metzger, M.E.; Kwan, J.A.; Harbison, J.E.; Fritz, C.L.; Riggs-Nagy, J.; Shindelbower, M.; Kramer, V.L. Development time of Culex mosquitoes in stormwater management structures in California. J. Am. Mosq. Control Assoc. 2008, 24, 90–97. [Google Scholar] [CrossRef]
- Reeves, W.C. Ecology of mosquitoes in relation to arboviruses. Annu. Rev. Entomol. 1965, 10, 25–46. [Google Scholar] [CrossRef]
- Schwarz, A.G.; Thorpe, J.P.; Redmann, R.E. Isolated Grasslands in the Boreal Forest Region of Western Canada. In Proceedings of the 10th North American Prairie Conference, Denton, TX, USA, 22–26 June 1986; Texas Woman’s University: Denton, TX, USA, 1986; pp. 22–26. [Google Scholar]
- Schwarz, A.G.; Wein, R.W. Grassland Ecosystems and Climate Change: Hypotheses for the Northern Grassland-Boreal Forest Ecotone. In Proceedings of the 12th North American Prairie Conference, Cedar Falls, IA, USA, 6–9 August 1992; Smith, D.D., Jacobs, C.A., Eds.; University of Northern Iowa: Cedar Falls, IA, USA, 1992; pp. 1–4. [Google Scholar]
- Schwarz, A.G.; Wein, R.W. Threatened dry grasslands in the continental boreal forests of Wood Buffalo National Park. Can. J. Bot. 1997, 75, 1363–1370. [Google Scholar] [CrossRef]
- Dobson, A. Climate variability, global change, immunity, and the dynamics of infectious diseas. Ecology 2009, 90, 920–927. [Google Scholar] [CrossRef]
- Patz, J.A.; Reisen, W.K. Immunology, climate change and vector-borne diseases. Trends Immunol. 2001, 22, 171–172. [Google Scholar] [CrossRef]
- Rohr, J.R.; Dobson, A.P.; Johnson, P.T.J.; Kilpatrick, A.M.; Paull, S.H.; Raffel, T.R.; Ruiz-Moreno, D.; Thomas, M.B. Frontiers in climate change—Disease research. Trends Ecol. Evol. 2011, 26, 270–277. [Google Scholar] [CrossRef]
- Meehl, G.A.; Stocker, T.F.; Collins, W.D.; Friedlingstein, P.; Gaye, A.T.; Gregory, J.M.; Kitoh, A.; Knutti, R.; Murphy, J. M.; Noda, A.; et al. Global Climate Projections. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 749–844. [Google Scholar]
- Zou, L.; Miller, S.N.; Schmidtmann, E.T. A GIS tool to estimate West Nile virus risk based on a degree-day model. Environ. Monit. Assess. 2007, 129, 413–420. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, C.C.; Jenkins, E.; Epp, T.; Waldner, C.; Curry, P.S.; Soos, C. Climate Change and West Nile Virus in a Highly Endemic Region of North America. Int. J. Environ. Res. Public Health 2013, 10, 3052-3071. https://doi.org/10.3390/ijerph10073052
Chen CC, Jenkins E, Epp T, Waldner C, Curry PS, Soos C. Climate Change and West Nile Virus in a Highly Endemic Region of North America. International Journal of Environmental Research and Public Health. 2013; 10(7):3052-3071. https://doi.org/10.3390/ijerph10073052
Chicago/Turabian StyleChen, Chen C., Emily Jenkins, Tasha Epp, Cheryl Waldner, Philip S. Curry, and Catherine Soos. 2013. "Climate Change and West Nile Virus in a Highly Endemic Region of North America" International Journal of Environmental Research and Public Health 10, no. 7: 3052-3071. https://doi.org/10.3390/ijerph10073052
APA StyleChen, C. C., Jenkins, E., Epp, T., Waldner, C., Curry, P. S., & Soos, C. (2013). Climate Change and West Nile Virus in a Highly Endemic Region of North America. International Journal of Environmental Research and Public Health, 10(7), 3052-3071. https://doi.org/10.3390/ijerph10073052