Genetic Detection of Pseudomonas spp. in Commercial Amazonian Fish

Abstract
:1. Introduction

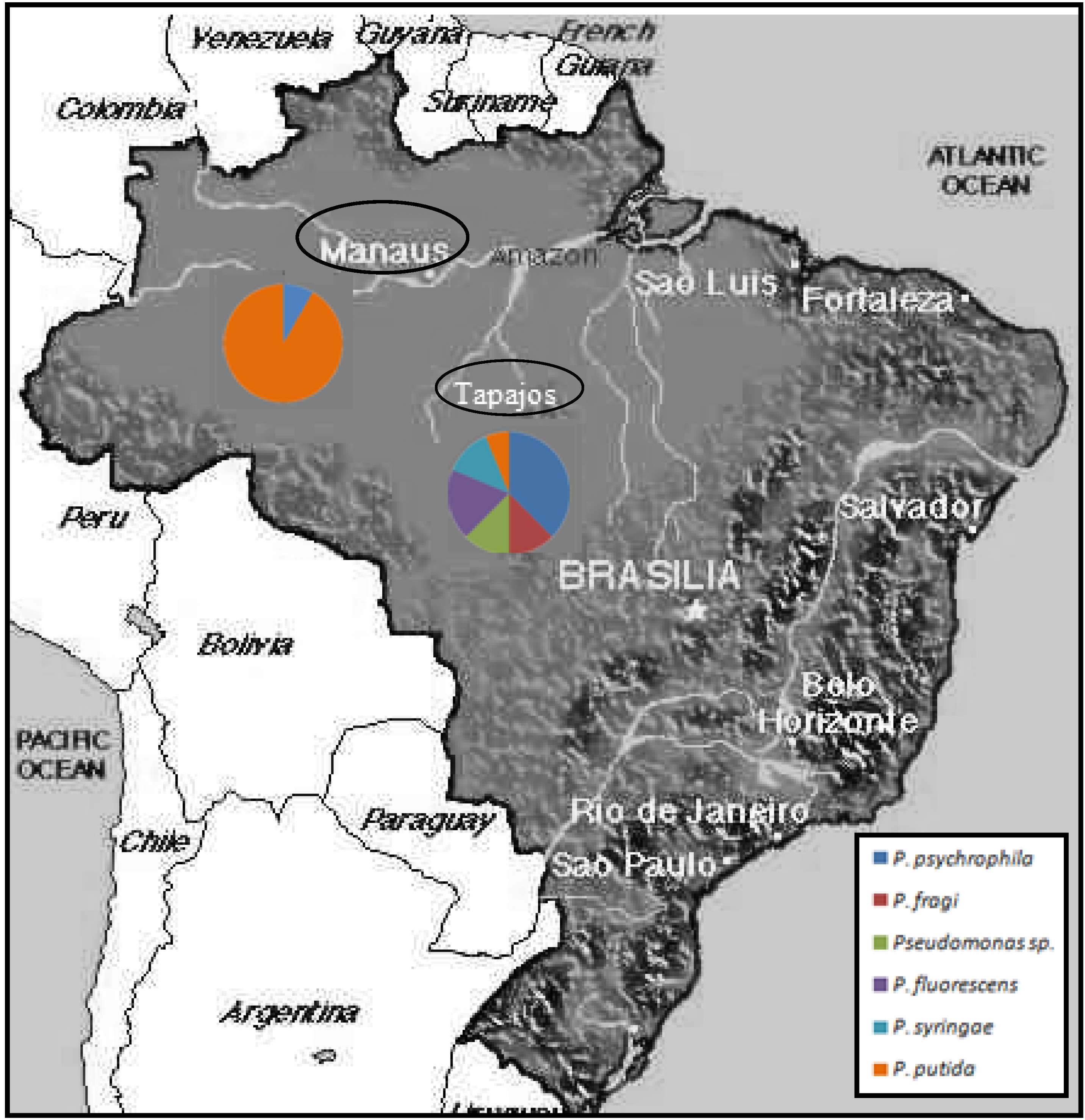{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Origin | Fish spp. | Common name | Pseudomonas species |
|---|---|---|---|---|
| T1 | Tapajós | Prochilodus nigricans | Curimatá | Pseudomonas psychrophila |
| T2 | Tapajós | Cetopsis candiru | Candiru | Pseudomonas spp. |
| T3 | Tapajós | Leporinus piau | Piau | Pseudomonas psychrophila |
| T4 | Tapajós | Leporinus piau | Piau | Pseudomonas syringae |
| T5 | Tapajós | Serrasalmus rhombeus | Piranha | Pseudomonas fragi |
| T6 | Tapajós | Leporinus piau | Piau | Pseudomonas fluorescens |
| T7 | Tapajós | Ageneiosus brevifilis | Bocudo | Pseudomonas fluorescens |
| T8 | Tapajós | Leporinus piau | Piau | Pseudomonas psychrophila |
| T9 | Tapajós | Leporinus piau | Piau | Pseudomonas syringae |
| T10 | Tapajós | Leporinus piau | Piau | Pseudomonas fluorescens |
| T11 | Tapajós | Leporinus piau | Piau | Pseudomonas spp. |
| T12 | Tapajós | Leporinus piau | Piau | Pseudomonas psychrophila |
| T13 | Tapajós | Prochilodus nigricans | Curimatá | Pseudomonas psychrophila |
| T14 | Tapajós | Leporinus piau | Piau | Pseudomonas fragi |
| T15 | Tapajós | Leporinus piau | Piau | Pseudomonas psychrophila |
| T16 | Tapajós | Leporinus piau | Piau | Pseudomonas putida |
| T17 | Tapajós | Leporinus piau | Piau | - |
| T18 | Tapajós | Leporinus piau | Piau | - |
| M1 | Negro | Chaetobranchopsis orbicularis | Acará branco | Pseudomonas putida |
| M2 | Negro | Astonotus ocellatus | Acará-açú | Pseudomonas putida |
| M3 | Negro | Astonotus ocellatus | Acará-açú | Pseudomonas putida |
| M4 | Negro | Astonotus ocellatus | Acará-açú | Pseudomonas putida |
| M5 | Negro | Osteoglossum bicirrhosum | Aruanà | Pseudomonas putida |
| M6 | Negro | Brachypatystoma rousseauxii | Dourada | Pseudomonas putida |
| M7 | Negro | Semaprochilodus insignis | Jaraquí | Pseudomonas putida |
| M8 | Negro | Plagioscion squamosissimus | Pescada | Pseudomonas putida |
| M9 | Negro | Phractocephalus hemioliopterus | Pirarara | Pseudomonas psychrophila |
| M10 | Negro | Pseudoplatystoma fasciatum | Surubim | Pseudomonas putida |
| M11 | Negro | Cichla temensis | Tucunaré | Pseudomonas putida |
| M12 | Negro | Cichla temensis | Tucunaré | Pseudomonas putida |
2. Experimental Section
2.1. Sampling

2.2. Genetic Analyses
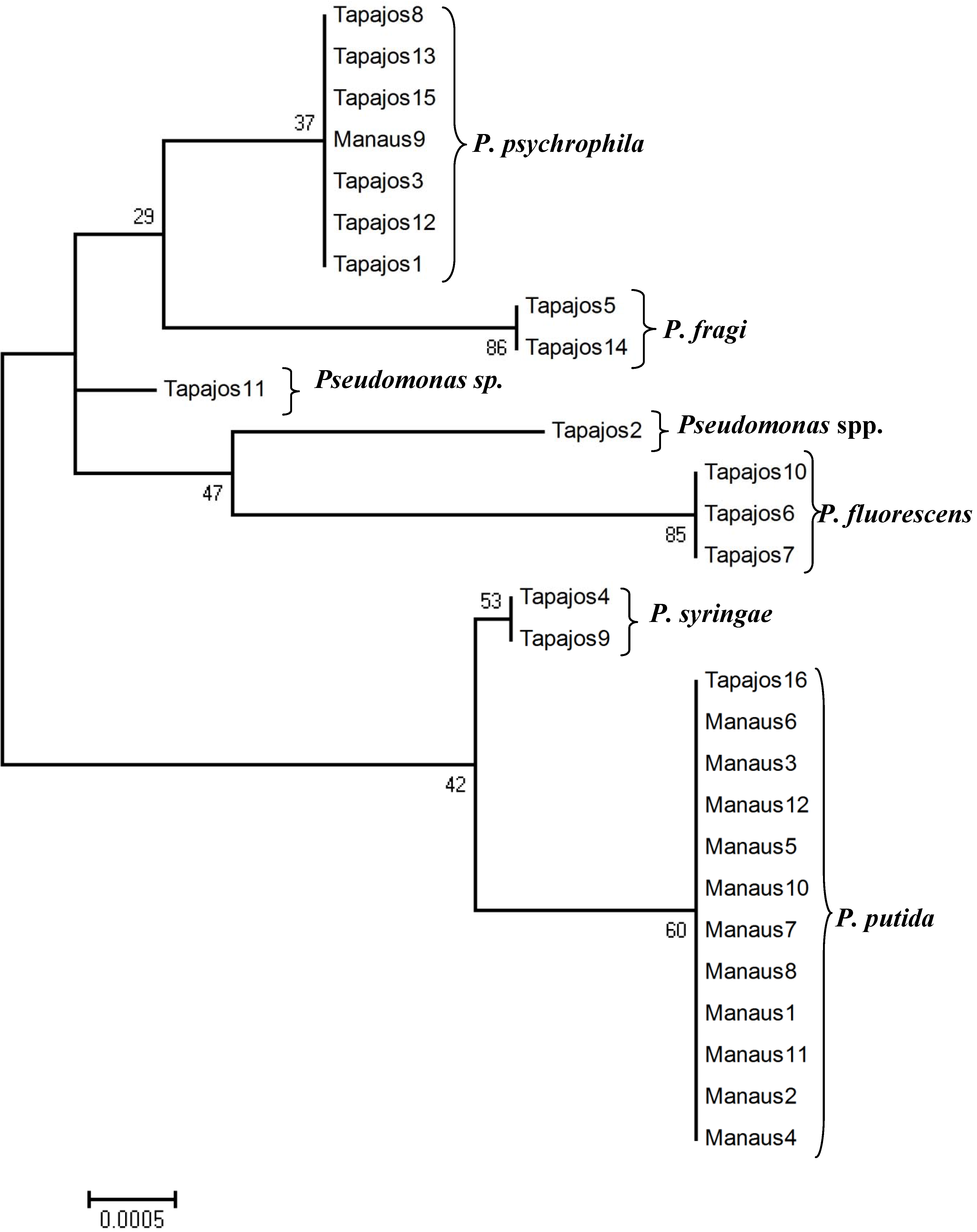
2.3. Sequence Edition and Phylogenetic Analysis
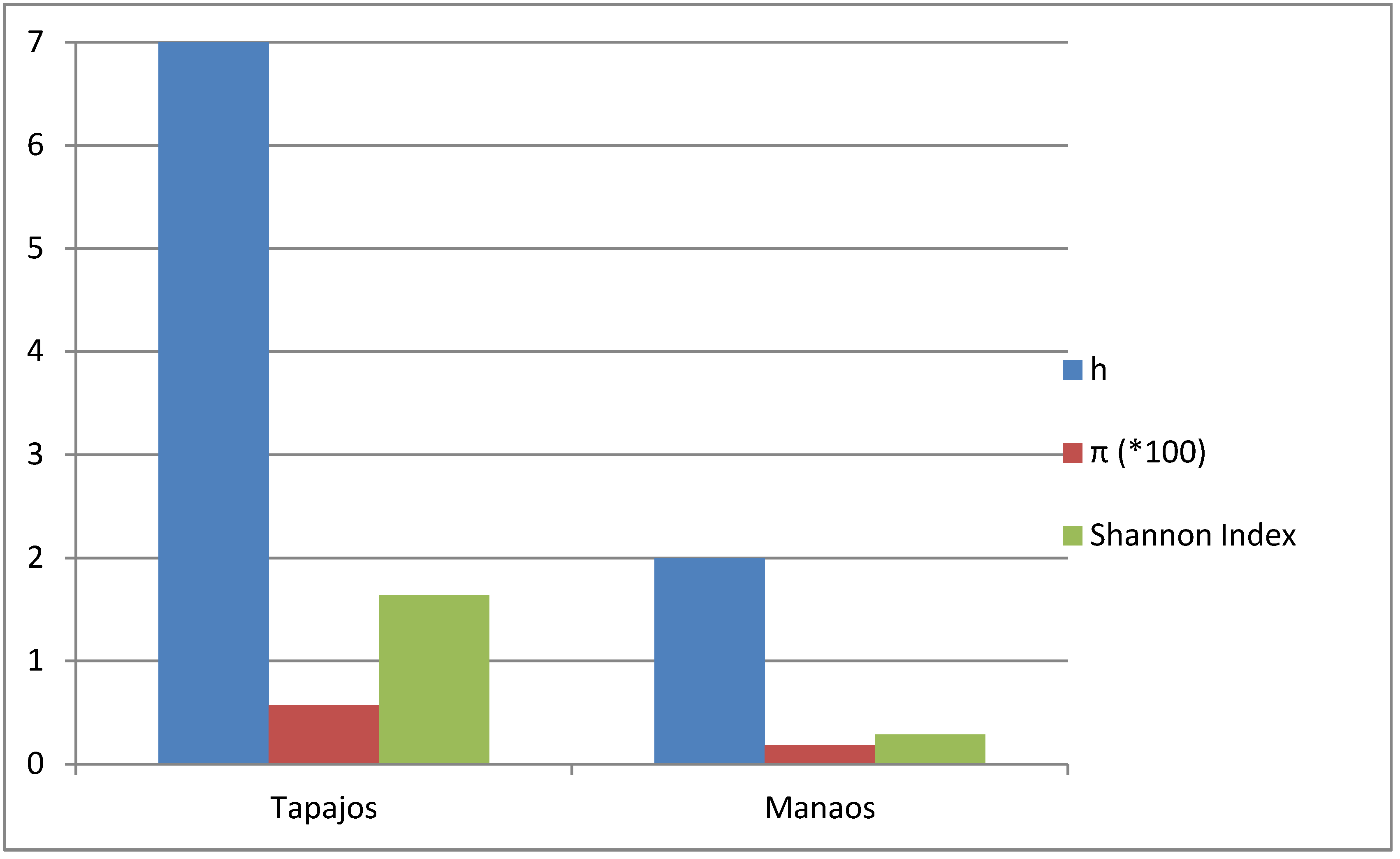
2.4. Pseudomonas Diversity Estimates
2.5. Statistics
3. Results and Discussion



| Pseudomonas species | Characteristics | Pathogenesis reported for humans |
|---|---|---|
| P. fluorescens | Opportunistic pathogen in fish [42] | Oncology patients [43] |
| P. fragi | Seafood spoilage [6,44] | No published data about this |
| Opportunistic microbiota [6] | ||
| P. psychrophila | No * | |
| P. putida | Seafood spoilage [44] | Immunodepressed patients [45] |
| Cosmopolitan opportunist [46] | Nosocomial infections [47] | |
| P. syringae | No * |
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Saint-Paul, U.; Zuanon, J.; Villacorta-Correa, M.A.; García, M.; Fabré, N.N.; Berger, U.; Junk, W.J. Fish communities in central amazonian white- and blackwater floodplains. Environ. Biol. Fishes 2000, 57, 235–250. [Google Scholar]
- Fernandes, C.C.; Podos, J.; Lundberg, J.G. Amazonian ecology: Tributaries enhance the diversity of electric fishes. Science 2004, 305, 1960–1962. [Google Scholar] [CrossRef]
- Ardura, A.; Pola, I.G.; Linde, A.R.; Garcia-Vazquez, E. DNA-based methods for species authentication of Amazonian commercial fish. Food Res. Int. 2010, 43, 2295–2302. [Google Scholar] [CrossRef]
- FAO/WHO: Food and Agriculture Organization of the United Nations/ World Health Organization, Microbiological Risk Assessment Series. In Risk Characterization of Microbiological Hazards in Food. Guidelines; WHO: Geneva, Switzerland, 2009; Volume 17, p. 116.
- Sakata, T. Microflora of Healthy Animals. In Methods for the Microbiological Examination of Fish and Shellfish Chichester; Austin, B., Austin, D.A., Eds.; Ellis Horwood Ltd.: England, UK, 1989; pp. 141–163. [Google Scholar]
- Gram, L.; Huss, H.H. Fresh and Processed Fish and Shellfish. In The Microbiological Safety and Quality of Foods; Lund, B.M., Baird-Parker, A.C., Gould, G.W., Eds.; Chapman & Hall: London, UK, 2000; pp. 472–506. [Google Scholar]
- Gram, L.; Ravn, L.; Rasch, M.; Bruhn, J.B.; Christensen, A.B.; Givskov, M. Food spoilage-interactions between food spoilage bacteria. Int. J. Food Microbiol. 2002, 78, 79–97. [Google Scholar] [CrossRef]
- Nyenje, M.E.; Odjadjare, C.E.; Odjadjare, L.; Tanih, N.F.; Green, E.; Ndip, R.N. Foodborne pathogens recovered from ready-to-eat foods from roadside cafeterias and retail outlets in Alice, Eastern Cape Province, South Africa: Public health implications. Int. J. Environ. Res. Public Health 2012, 9, 2608–2619. [Google Scholar] [CrossRef]
- Bagshaw, S.M.; Laupland, K.B. Epidemiology of intensive care unit-acquired urinary tract infections. Curr. Opin. Infect. Dis. 2006, 19, 67–71. [Google Scholar] [CrossRef]
- Zilberberg, M.D.; Shorr, A.F. Epidemiology of healthcare-associated pneumonia (HCAP). Semin. Respir. Crit. Care Med. 2009, 30, 10–15. [Google Scholar] [CrossRef]
- Mena, K.D.; Gerba, C.P. Risk assessment of Pseudomonas aeruginosa in water. Rev. Environ. Contam. Toxicol. 2009, 201, 71–115. [Google Scholar] [CrossRef]
- Jiwa, S.F.H.; Krovacek, K.; Wadstrom, T. Enterotoxigenic bacteria in food and water from an ethiopian community. Appl. Environ. Microbiol. 1981, 41, 1010–1019. [Google Scholar]
- Jertborn, M.; Svennerholm, A.M. Enterotoxin-producing bacteria isolated from Swedish travellers with diarrhoea. Scand. J. Infect. Dis. 1991, 23, 473–479. [Google Scholar] [CrossRef]
- Bockemühl, J.; Fleischer, K.; Bednarek, I. A cholera-like illness in a traveller due to a mixed infection with enterotoxigenic Escherichia coli, Vibrio parahaemolyticus and Pseudomonas aeruginosa. Infection 1983, 11, 272–274. [Google Scholar] [CrossRef]
- Adlard, P.A.; Kirov, S.M.; Sanderson, K.; Cox, G.E. Pseudomonas aeruginosa as a cause of infectious diarrhoea. Epidemiol. Infect. 1998, 121, 237–241. [Google Scholar] [CrossRef]
- Wong, S.; Street, D.; Delgado, S.I.; Klontz, K.C. Recalls of foods and cosmetics due to microbial contamination reported to the U.S. Food and Drug Administration. J. Food Prot. 2000, 63, 1113–1116. [Google Scholar]
- Craun, G.F.; Brunkard, J.M.; Yoder, J.S.; Roberts, V.A.; Carpenter, J.; Wade, T.; Calderon, R.L.; Roberts, J.M.; Beach, M.J.; Roy, S.L. Causes of outbreaks associated with drinking water in the United States from 1971 to 2006. Clin. Microbiol. Rev. 2010, 23, 507–528. [Google Scholar] [CrossRef]
- Product Inspection of Imported Fish. Csnsdian Food Inspection Agency. 2013. Available online: http://www.inspection.gc.ca/english/fssa/fispoi/import/pol/procprode.shtml (accessed on 26 August 2013).
- Microbiological Criteria. Health and Consumers. European Commission. 2011. Available online: http://ec.europa.eu/food/food/biosafety/salmonella/microbio_en.htm (accessed on 26 August 2013).
- Food Safety and Inspection Service. United States Department of Agriculture. Available online: http://www.fsis.usda.gov/ (accessed on 26 August 2013).
- Bennett, A.R.; Greenwood, D.; Tennant, C.; Banks, J.G.; Betts, R.P. Rapid and definitive detection of Salmonella in foods by PCR. Lett. Appl. Microbiol. 1998, 26, 437–441. [Google Scholar]
- Wang, R.-F.; Cao, W.W.; Cerniglia, C.E. A universal protocol for PCR detection of 13 species of foodborne pathogens in foods. J. Appl. Microbiol. 1997, 83, 727–736. [Google Scholar]
- Estoup, A.; Largiader, C.R.; Perrot, E.; Chourrout, D. Rapid one-tube DNA extraction for reliable PCR detection of fish polymorphic markers and transgenes. Mol. Mar. Biol. Biotechnol. 1996, 5, 295–298. [Google Scholar]
- Spilker, T.; Coenye, T.; Vandamme, P.; LiPuma, J.J. PCR-based assay for differentiation of Pseudomonas aeruginosa from other Pseudomonas species recovered from cystic fibrosis patients. J. Clin. Microbiol. 2004, 42, 2074–2079. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. Clustal-W, Improving the sensitivity of progressive multiple sequence alignment trough sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evolut. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Hebert, P.; Cywinska, A.; Ball, S.; deWaard, J. Biological identification through DNA barcodes. Proc. R. Soc. B: Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Kumar, S.; Gadadkar, S. Efficiency of the neighbour-joining method in reconstructing deep and shallow evolutionary relationships in large phylogenies. J. Mol. Evolut. 2000, 51, 544–553. [Google Scholar]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evolut. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov/ (accessed on 26 August 2013).
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987; p. 512. [Google Scholar]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. J. Evol. Bioinform. Online 2005, 1, 47–50. [Google Scholar]
- Shiose, J.; Wakabayashi, H.; Tominaga, M.; Egusa, S. A report on a disease of cultured carp due to a capsulated Pseudomonas. Fish Pathol. 1974, 9, 79–83. [Google Scholar] [CrossRef]
- Alderman, D.J.; Polglase, J.L. Pathogens, Parasites and Commensals. In Freshwater Crayfish—Biology, Management and Exploitation; Holdich, D.M., Lowry, R.S., Eds.; Timber Press: Portland, OR, USA, 1998; pp. 168–187. [Google Scholar]
- Kusuda, R.; Toyoshima, R. Characteristics of a pathogenic Pseudomonas isolated from cultured yellowtail. Fish Pathol. 1976, 1, 133–139. [Google Scholar] [CrossRef]
- Uryu, Y.; Malm, O.; Thornton, I.; Payne, I.; Cleary, D. Mercury contamination of fish and its implications for other wildlife of the Tapajós Basin, Brazilian Amazon. Conserv. Biol. 2001, 15, 438–446. [Google Scholar] [CrossRef]
- Altinok, I.; Kayisa, S.; Capkin, E. Pseudomonas putida infection in rainbow trout. Aquaculture 2006, 261, 850–855. [Google Scholar] [CrossRef]
- Yumoto, I; Kusano, T.; Shingyo, T.; Nodasaka, Y.; Matsuyama, H.; Okuyama, H. Assignment of Pseudomonas sp. strain E-3 to Pseudomonas psychropdomonas sp. strain E-3 to Pseudomonas psychrophila spp. nov., a new facultatively psychrophilic bacterium. Extremophiles 2001, 5, 343–349. [Google Scholar] [CrossRef]
- Morita, R.Y. Psychrophilic bacteria. Bacteriol. Rev. 1975, 39, 144–167. [Google Scholar]
- Hirano, S.S.; Upper, C.D. Population biology and epidemiology of Pseudomonas syringae. Annu. Rev. Phytopathol. 1990, 28, 155–177. [Google Scholar] [CrossRef]
- Bruno, D.W.; Ellis, A.E. Salmonid Disease Management. Dev. Aquac. Fish. Sci. 1996, 29, 729–824. [Google Scholar]
- Hsueh, P.R.; Teng, L.J.; Pan, H.J.; Chen, Y.C.; Sun, C.C.; Ho, S.W.; Luh, K.T. Outbreak of Pseudomonas fluorescens bacteremia among oncology patients. J. Clin. Microbiol. 1998, 36, 2914–2917. [Google Scholar]
- Von Graevenitz, A.; Weinstein, J. Pathogenic significance of Pseudomonas fluorescens and Pseudomonas putida. Yale J. Biol. Med. 1971, 44, 265–273. [Google Scholar]
- Yoshino, Y.; Kitazawa, T.; Kamimura, M.; Tatsuno, K.; Ota, Y.; Yotsuyanagi, H. Pseudomonas putida bacteremia in adult patients: five case reports and a review of the literature. J. Infect. Chemother. 2011, 17, 278–282. [Google Scholar] [CrossRef]
- Timmis, K.N. Pseudomonas putida: A cosmopolitan opportunist par excellence. Environ. Microbiol. 2002, 4, 779–781. [Google Scholar] [CrossRef]
- Yamamoto, S.; Kasai, H.; Arnold, D.L.; Jackson, R.W.; Vivian, A.; Harayama, S. Phylogeny of the genus Pseudomonas: Intrageneric structure reconstructed from the nucleotide sequences of gyrB and rpoD genes. Microbiology 2000, 146, 2385–2394. [Google Scholar]
- Franzetti, L.; Scarpellini, M. Characterisation of Pseudomonas spp. isolated from foods. Ann. Microbiol. 2007, 57, 39–47. [Google Scholar]
- Mulet, M.; Lalucat, J.; García-Valdés, E. DNA sequence-based analysis of the Pseudomonas species. Environ. Microbiol. 2010, 12, 1513–1530. [Google Scholar]
- Picot, L.; Mezghani-Abdelmoula, S.; Merieaua, A.; Lerouxb, P.; Cazina, L.; Orangea, N.; Feuilloley, M.G.J. Pseudomonas fluorescens as a potential pathogen: Adherence to nerve cells. Microbes Infect. 2001, 3, 985–995. [Google Scholar] [CrossRef]
- Kim, S.E.; Park, S.H.; Park, H.B.; Park, K.H.; Kim, S.H.; Jung, S.I.; Shin, J.H.; Jang, H.C.; Kang, S.J. Nosocomial Pseudomonas putida bacteremia: High rates of carbapenem resistance and mortality. Chonnam Med. J. 2012, 48, 91–95. [Google Scholar] [CrossRef]
- Gilardi, G.L. Infrequently encountered Pseudomonas species causing infection in humans. Ann. Int. Med. 1972, 77, 211–215. [Google Scholar] [CrossRef]
- Wilson, M.R.; Allard, M.W.; Brown, E.W. The forensic analysis of foodborne bacterial pathogens in the age of whole-genome sequencing. Cladistics 2013, 29, 449–461. [Google Scholar] [CrossRef]
- LeChevallier, M.W.; Seidler, R.J.; Evans, T.M. Enumeration and characterization of standard plate count bacteria in chlorinated and raw water supplies. Appl. Environ. Microbiol. 1980, 40, 922–930. [Google Scholar]
- Hihgsmith, A.K.; Abshire, R.L. Evaluation of most-probable-number technique for the enumeration of Pseudomonas aeruginosa. Appl. Microbiol. 1975, 30, 596–601. [Google Scholar]
- Breeuwer, P.; Abee, T. Assessment of viability of microorganisms employing fluorescence techniques. Int. J. Food Microbiol. 2000, 55, 193–200. [Google Scholar] [CrossRef]
- Kogure, K.; Simidu, U.; Taga, N. A tentative direct microscopic method for counting living marine bacteria. Can. J. Microbiol. 1979, 25, 415–420. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ardura, A.; Linde, A.R.; Garcia-Vazquez, E. Genetic Detection of Pseudomonas spp. in Commercial Amazonian Fish. Int. J. Environ. Res. Public Health 2013, 10, 3954-3966. https://doi.org/10.3390/ijerph10093954
Ardura A, Linde AR, Garcia-Vazquez E. Genetic Detection of Pseudomonas spp. in Commercial Amazonian Fish. International Journal of Environmental Research and Public Health. 2013; 10(9):3954-3966. https://doi.org/10.3390/ijerph10093954
Chicago/Turabian StyleArdura, Alba, Ana R. Linde, and Eva Garcia-Vazquez. 2013. "Genetic Detection of Pseudomonas spp. in Commercial Amazonian Fish" International Journal of Environmental Research and Public Health 10, no. 9: 3954-3966. https://doi.org/10.3390/ijerph10093954
APA StyleArdura, A., Linde, A. R., & Garcia-Vazquez, E. (2013). Genetic Detection of Pseudomonas spp. in Commercial Amazonian Fish. International Journal of Environmental Research and Public Health, 10(9), 3954-3966. https://doi.org/10.3390/ijerph10093954