Interference in Pheromone-Responsive Conjugation of a High-Level Bacitracin Resistant Enterococcus faecalis Plasmid of Poultry Origin

Abstract
:1. Introduction
2. Material and Methods
2.1. Bacteria, Media and Antibiotics
2.2. PCR, Specific Primers and DNA Sequencing
2.3. Filter Mating
2.4. Pheromone Induction and Aggregation
2.5. Pheromone-Responsive Conjugation Experiments
2.6. Minimum Inhibitory Concentration (MIC) Determination
2.7. Production of Antiserum and Interference in Pheromone-Responsive Conjugation
2.8. DNA Microarrays
2.9. Pulsed-Field Gel Electrophoresis (PFGE)
2.10. Southern Hybridization
2.11. SDS-PAGE and Western Blotting
2.12. Statistical Analysis
3. Results
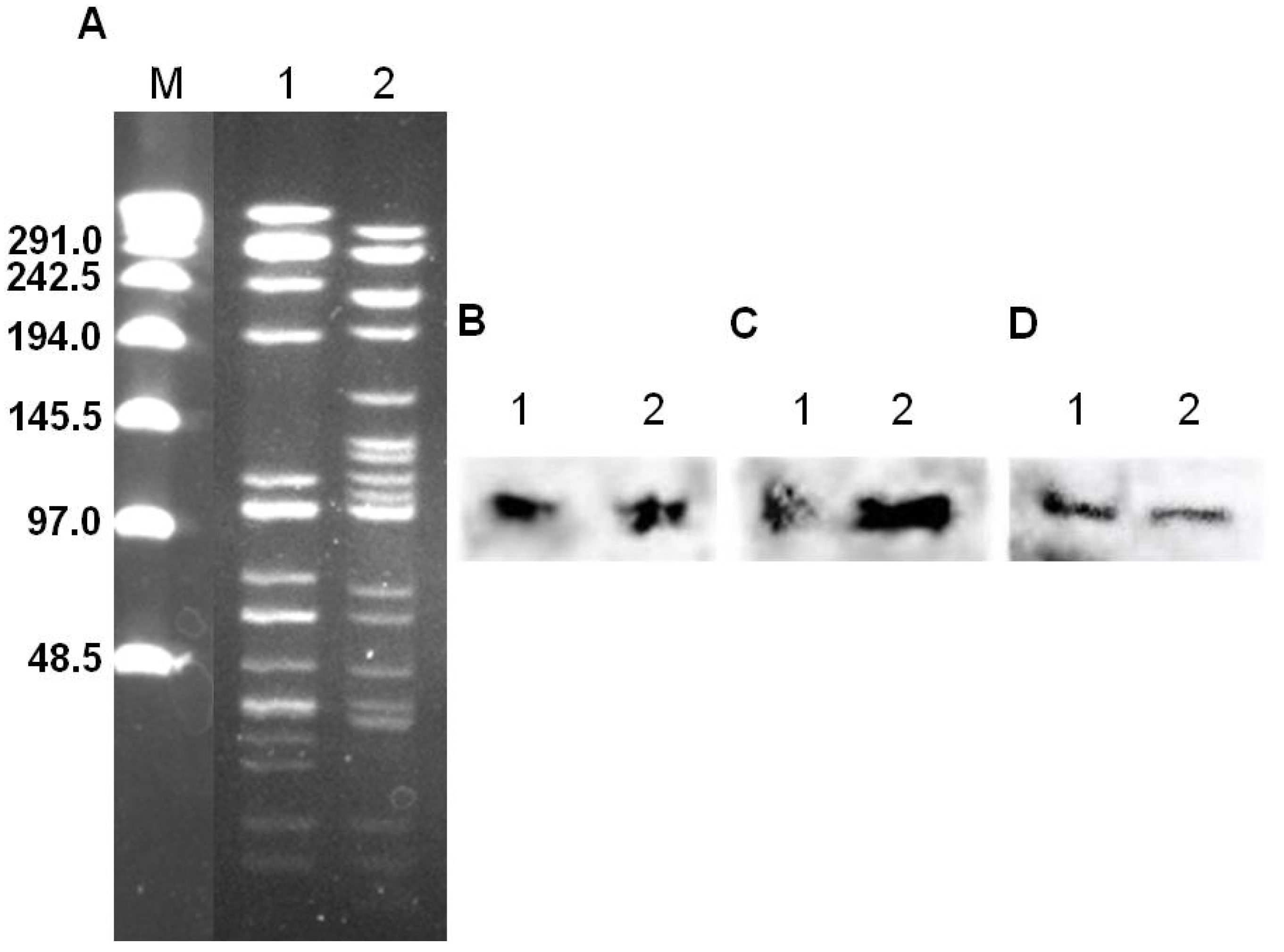
3.1. Conjugal Transfer and Analysis of Transconjugants

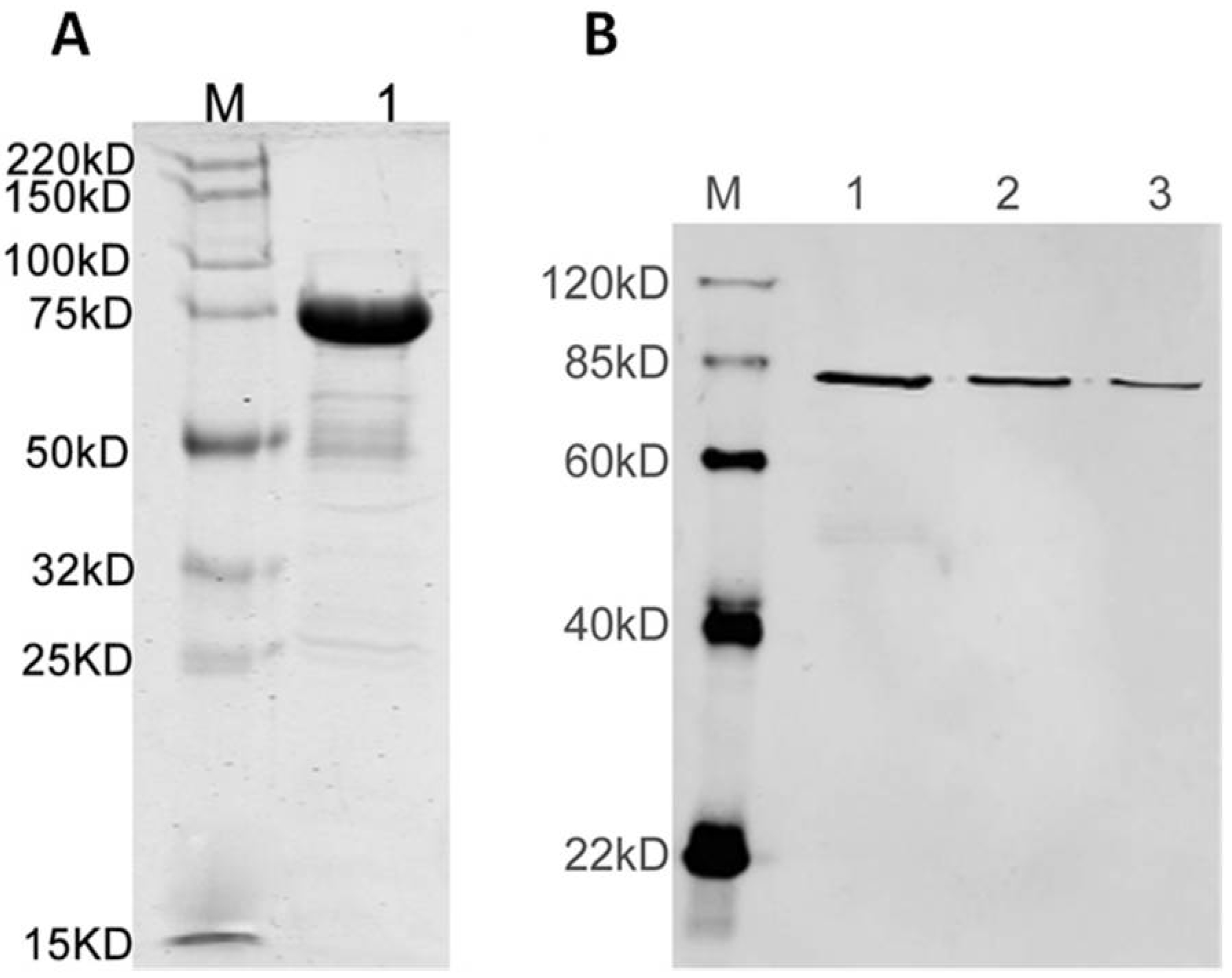
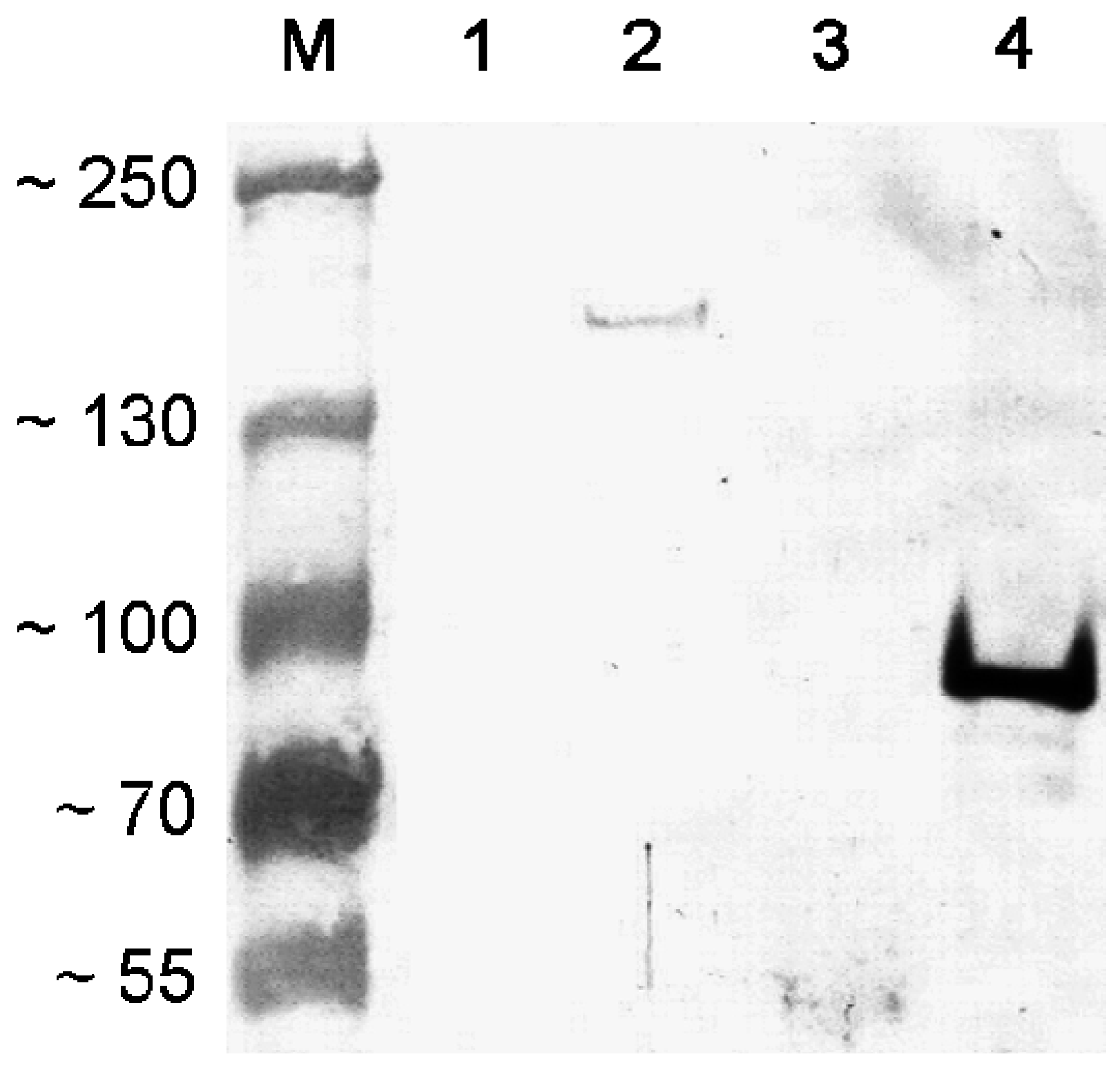
3.2. Antisera Titer and Specificity


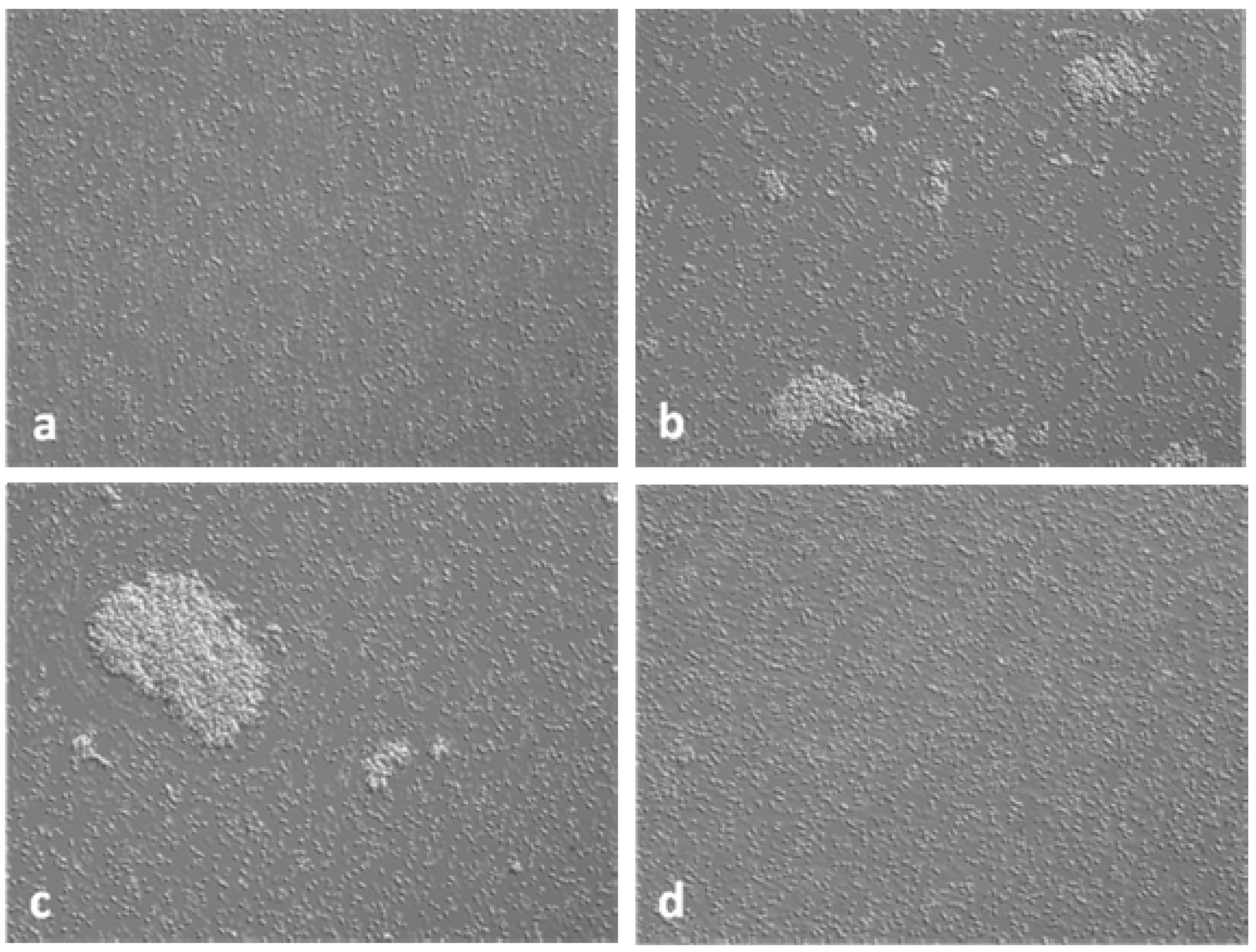
3.3. Interference in the Formation of Aggregates Using Anti-AS44–560 Antibodies


3.4. Interference in Pheromone-Responsive Conjugation Using Anti-AS44–560 Antibodies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
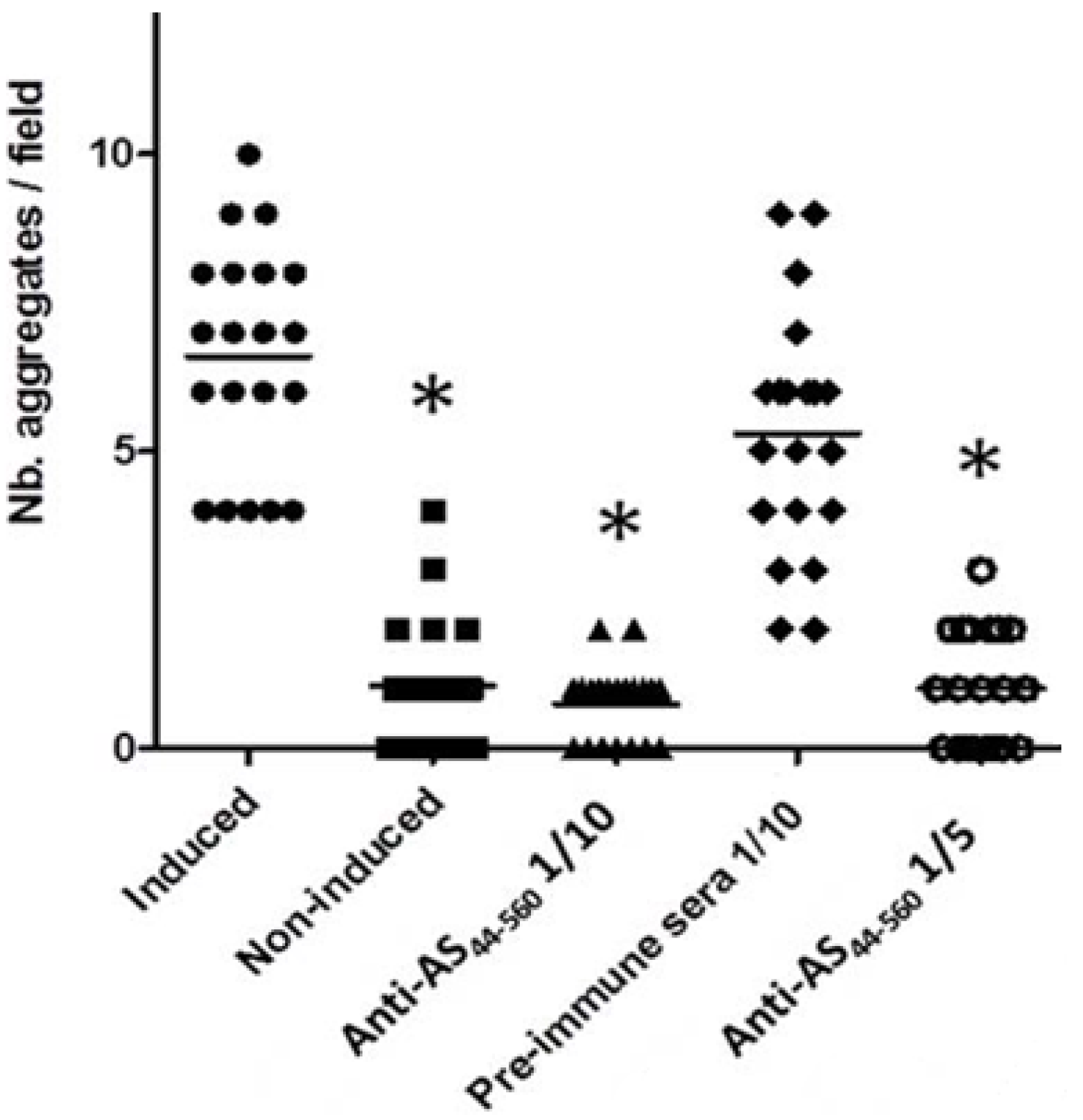{kind=link}
| Exposure to pheromone a | Antibodies | Transfer frequency in 20-min mating (no. of transconjugants per donor cell) c |
|---|---|---|
| + b | None | 4.6 × 10−3 |
| - | None | 2.2 × 10−6 |
| + | pre-immune serum | 8.3 × 10−3 |
| + | None | 4.6 × 10−3 |
| + | polyclonal anti-AS44–560 | 4.4 × 10−5 |
| + | None | 7.5 × 10−2 |
| - | None | 6.3 × 10−7 |
4. Discussion
5. Conclusions
Acknowledgements
Conflicts of Interest
References
- American Society for Microbiology, Manual of Clinical Microbiology, 8th ed.; Murray, P.R.; Baron, E.J. (Eds.) ASM Press: Washington, DC, USA, 2003; pp. 422–423.
- Gregersen, R.H.; Petersen, A.; Christensen, H.; Bisgaard, M. Multilocus sequence typing of Enterococcus faecalis isolates demonstrating different lesion types in broiler breeders. Avian Pathol. 2010, 39, 435–440. [Google Scholar] [CrossRef]
- Tankson, J.D.; Thaxton, J.P.; Vizzier-Thaxton, Y. Pulmonary hypertension syndrome in broilers caused by Enterococcus faecalis. Infect. Immun. 2001, 69, 6318–6322. [Google Scholar] [CrossRef]
- Landman, W.J.; Gruys, E.; Dwars, R.M. A syndrome associated with growth depression and amyloid arthropathy in layers: A preliminary report. Avian Pathol. 1994, 23, 461–470. [Google Scholar] [CrossRef]
- Fertner, M.E.; Olsen, R.H.; Bisgaard, M.; Christensen, H. Transmission and genetic diversity of Enterococcus faecalis among layer chickens during hatch. Acta Vet. Scand. 2011, 53, 56. [Google Scholar] [CrossRef]
- Olsen, R.H.; Schonheyder, H.C.; Christensen, H.; Bisgaard, M. Enterococcus faecalis of human and poultry origin share virulence genes supporting the zoonotic potential of E. faecalis. Zoonoses Public Health 2011, 59, 256–263. [Google Scholar]
- Coque, T.M. Evolutionary Biology of Pathogenic Enterococci. In Evolutionary Biology of Bacteria and Fungal Pathogens; Baquero, F., Nombela, C., Cassell, G.H., Guitierrez, J.A., Eds.; ASM Press: Washington, DC, USA, 2008; pp. 501–521. [Google Scholar]
- Aarestrup, F.M. Occurrence of glycopeptide resistance among Enterococcus faecium isolates from conventional and ecological poultry farms. Microb. Drug Resist. 1995, 1, 255–257. [Google Scholar] [CrossRef]
- Ghidan, A.; Dobay, O.; Kaszanyitzky, E.J.; Samu, P.; Amyes, S.G.; Nagy, K.; Rozgonyi, F. Vancomycin resistant enterococci (VRE) still persist in slaughtered poultry in hungary 8 years after the ban on avoparcin. Acta Microbiol. Immunol. Hung. 2008, 55, 409–417. [Google Scholar] [CrossRef]
- Donelli, G.; Paoletti, C.; Baldassarri, L.; Guaglianone, E.; di Rosa, R.; Magi, G.; Spinaci, C.; Facinelli, B. Sex pheromone response, clumping, and slime production in enterococcal strains isolated from occluded biliary stents. J. Clin. Microbiol. 2004, 42, 3419–3427. [Google Scholar] [CrossRef]
- Dunny, G.M.; Brown, B.L.; Clewell, D.B. Induced cell aggregation and mating in Streptococcus faecalis: Evidence for a bacterial sex pheromone. Proc. Natl. Acad. Sci. USA 1978, 75, 3479–3483. [Google Scholar] [CrossRef]
- Wirth, R. The sex pheromone system of Enterococcus faecalis. More than just a plasmid-collection mechanism? Eur. J. Biochem. 1994, 222, 235–246. [Google Scholar] [CrossRef]
- Dunny, G.M.; Antiporta, M.H.; Hirt, H. Peptide pheromone-induced transfer of plasmid pCF10 in Enterococcus faecalis: Probing the genetic and molecular basis for specificity of the pheromone response. Peptides 2001, 22, 1529–1539. [Google Scholar] [CrossRef]
- Chung, J.W.; Bensing, B.A.; Dunny, G.M. Genetic analysis of a region of the Enterococcus faecalis plasmid pCF10 involved in positive regulation of conjugative transfer functions. J. Bacteriol. 1995, 177, 2107–2117. [Google Scholar]
- Olmsted, S.B.; Kao, S.M.; van Putte, L.J.; Gallo, J.C.; Dunny, G.M. Role of the pheromone-inducible surface protein Asc10 in mating aggregate formation and conjugal transfer of the Enterococcus faecalis plasmid pCF10. J. Bacteriol. 1991, 173, 7665–7672. [Google Scholar]
- Manson, J.M.; Keis, S.; Smith, J.M.; Cook, G.M. Acquired bacitracin resistance in Enterococcus faecalis is mediated by an ABC transporter and a novel regulatory protein, BcrR. Antimicrob. Agents Chemother. 2004, 48, 3743–3748. [Google Scholar] [CrossRef]
- Dufour, M.; Manson, J.M.; Bremer, P.J.; Dufour, J.P.; Cook, G.M.; Simmonds, R.S. Characterization of monolaurin resistance in Enterococcus faecalis. Appl. Environ. Microbiol. 2007, 73, 5507–5515. [Google Scholar] [CrossRef]
- Waters, C.M.; Dunny, G.M. Analysis of functional domains of the Enterococcus faecalis pheromone-induced surface protein aggregation substance. J. Bacteriol. 2001, 183, 5659–5667. [Google Scholar] [CrossRef]
- Tremblay, C.L.; Letellier, A.; Quessy, S.; Boulianne, M.; Daignault, D.; Archambault, M. Multiple-antibiotic resistance of Enterococcus faecalis and Enterococcus faecium from cecal contents in broiler chicken and turkey flocks slaughtered in Canada and plasmid colocalization of tetO and ermB genes. J. Food Prot. 2011, 74, 1639–1648. [Google Scholar] [CrossRef]
- Jacob, A.E.; Hobbs, S.J. Conjugal transfer of plasmid-borne multiple antibiotic resistance in Streptococcus faecalis var. zymogenes. J. Bacteriol. 1974, 117, 360–372. [Google Scholar]
- Quirantes, R.; Galvez, A.; Valdivia, E.; Martin, I.; Martinez-Bueno, M.; Mendez, E.; Maqueda, M. Purification of sex pheromones specific for pMB1 and pMB2 plasmids of Enterococcus faecalis S-48. Can. J. Microbiol. 1995, 41, 629–632. [Google Scholar]
- Seno, Y.; Kariyama, R.; Mitsuhata, R.; Monden, K.; Kumon, H. Clinical implications of biofilm formation by Enterococcus faecalis in the urinary tract. Acta Med. Okayama 2005, 59, 79–87. [Google Scholar]
- Basic Local Alignment Search Tool (BLAST) Database. Available online: http://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 14 December 2012).
- Clark, N.C.; Olsvik, O.; Swenson, J.M.; Spiegel, C.A.; Tenover, F.C. Detection of a streptomycin/spectinomycin adenylyltransferase gene (aadA) in Enterococcus faecalis. Antimicrob. Agents Chemother. 1999, 43, 157–160. [Google Scholar] [CrossRef]
- Shryock, T.R. National Committee for Clinical Laboratory Standards. In Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals: Approved Standard: M31A3, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]
- Champagne, J.; Diarra, M.S.; Rempel, H.; Topp, E.; Greer, C.W.; Harel, J.; Masson, L. Development of a DNA microarray for enterococcal species, virulence, and antibiotic resistance gene determinations among isolates from poultry. Appl. Environ. Microbiol. 2011, 77, 2625–2633. [Google Scholar] [CrossRef]
- Diarra, M.S.; Rempel, H.; Champagne, J.; Masson, L.; Pritchard, J.; Topp, E. Distribution of antimicrobial resistance and virulence genes in Enterococcus spp. and Characterization of isolates from broiler chickens. Appl. Environ. Microbiol. 2010, 76, 8033–8043. [Google Scholar]
- Matushek, M.G.; Bonten, M.J.; Hayden, M.K. Rapid preparation of bacterial DNA for pulsed-field gel electrophoresis. J. Clin. Microbiol. 1996, 34, 2598–2600. [Google Scholar]
- Garcia-Migura, L.; Pleydell, E.; Barnes, S.; Davies, R.H.; Liebana, E. Characterization of vancomycin-resistant Enterococcus faecium isolates from broiler poultry and pig farms in England and Wales. J. Clin. Microbiol. 2005, 43, 3283–3289. [Google Scholar]
- Galli, D.; Lottspeich, F.; Wirth, R. Sequence analysis of Enterococcus faecalis aggregation substance encoded by the sex pheromone plasmid pAD1. Mol. Microbiol. 1990, 4, 895–904. [Google Scholar] [CrossRef]
- An, F.Y.; Clewell, D.B. Characterization of the determinant (traB) encoding sex pheromone shutdown by the hemolysin/bacteriocin plasmid pAD1 in Enterococcus faecalis. Plasmid 1994, 31, 215–221. [Google Scholar] [CrossRef]
- Paulsen, I.T.; Banerjei, L.; Myers, G.S.; Nelson, K.E.; Seshadri, R.; Read, T.D.; Fouts, D.E.; Eisen, J.A.; Gill, S.R.; Heidelberg, J.F.; et al. Role of mobile DNA in the evolution of vancomycin-resistant Enterococcus faecalis. Science 2003, 299, 2071–2074. [Google Scholar] [CrossRef]
- Palmer, K.L.; Kos, V.N.; Gilmore, M.S. Horizontal gene transfer and the genomics of enterococcal antibiotic resistance. Curr. Opin. Microbiol. 2010, 13, 632–639. [Google Scholar] [CrossRef]
- Hirt, H.; Wanner, G.; Galli, D.; Wirth, R. Biochemical, immunological and ultrastructural characterization of aggregation substances encoded by Enterococcus faecalis sex-pheromone plasmids. Eur. J. Biochem. 1993, 211, 711–716. [Google Scholar] [CrossRef]
- Vignaroli, C.; Zandri, G.; Aquilanti, L.; Pasquaroli, S.; Biavasco, F. Multidrug-resistant enterococci in animal meat and faeces and co-transfer of resistance from an Enterococcus durans to a human Enterococcus faecium. Curr. Microbiol. 2011, 62, 1438–1447. [Google Scholar] [CrossRef]
- Clewell, D.B. Properties of Enterococcus faecalis plasmid pAD1, a member of a widely disseminated family of pheromone-responding, conjugative, virulence elements encoding cytolysin. Plasmid 2007, 58, 205–227. [Google Scholar] [CrossRef]
- Weaver, K.E.; Jensen, K.D.; Colwell, A.; Sriram, S.I. Functional analysis of the Enterococcus faecalis plasmid pAD1-encoded stability determinant par. Mol. Microbiol. 1996, 20, 53–63. [Google Scholar] [CrossRef]
- Christie, P.J.; Korman, R.Z.; Zahler, S.A.; Adsit, J.C.; Dunny, G.M. Two conjugation systems associated with Streptococcus faecalis plasmid pCF10: Identification of a conjugative transposon that transfers between S. faecalis and Bacillus subtilis. J. Bacteriol. 1987, 169, 2529–2536. [Google Scholar]
- Francia, M.V.; Clewell, D.B. Transfer origins in the conjugative Enterococcus faecalis plasmids pAD1 and pAM373: Identification of the pAD1 nic site, a specific relaxase and a possible TraG-like protein. Mol. Microbiol. 2002, 45, 375–395. [Google Scholar] [CrossRef]
- Ike, Y.; Tanimoto, K.; Tomita, H.; Takeuchi, K.; Fujimoto, S. Efficient transfer of the pheromone-independent Enterococcus faecium plasmid pMG1 (Gmr) (65.1 kilobases) to Enterococcus strains during broth mating. J. Bacteriol. 1998, 180, 4886–4892. [Google Scholar]
- Makinen, P.L.; Clewell, D.B.; An, F.; Makinen, K.K. Purification and substrate specificity of a strongly hydrophobic extracellular metalloendopeptidase (“gelatinase”) from Streptococcus faecalis (strain 0G1–10). J. Biol. Chem. 1989, 264, 3325–3334. [Google Scholar]
- Waters, C.M.; Antiporta, M.H.; Murray, B.E.; Dunny, G.M. Role of the Enterococcus faecalis GelE protease in determination of cellular chain length, supernatant pheromone levels, and degradation of fibrin and misfolded surface proteins. J. Bacteriol. 2003, 185, 3613–3623. [Google Scholar] [CrossRef]
- Schlievert, P.M.; Chuang-Smith, O.N.; Peterson, M.L.; Cook, L.C.; Dunny, G.M. Enterococcus faecalis endocarditis severity in rabbits is reduced by IgG Fabs interfering with aggregation substance. PLoS One 2010, 5, e13194. [Google Scholar] [CrossRef]
- Laverde Gomez, J.A.; Hendrickx, A.P.; Willems, R.J.; Top, J.; Sava, I.; Huebner, J.; Witte, W.; Werner, G. Intra- and interspecies genomic transfer of the Enterococcus faecalis pathogenicity island. PLoS One 2011, 6, e16720. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tremblay, C.-L.; Archambault, M. Interference in Pheromone-Responsive Conjugation of a High-Level Bacitracin Resistant Enterococcus faecalis Plasmid of Poultry Origin. Int. J. Environ. Res. Public Health 2013, 10, 4245-4260. https://doi.org/10.3390/ijerph10094245
Tremblay C-L, Archambault M. Interference in Pheromone-Responsive Conjugation of a High-Level Bacitracin Resistant Enterococcus faecalis Plasmid of Poultry Origin. International Journal of Environmental Research and Public Health. 2013; 10(9):4245-4260. https://doi.org/10.3390/ijerph10094245
Chicago/Turabian StyleTremblay, Cindy-Love, and Marie Archambault. 2013. "Interference in Pheromone-Responsive Conjugation of a High-Level Bacitracin Resistant Enterococcus faecalis Plasmid of Poultry Origin" International Journal of Environmental Research and Public Health 10, no. 9: 4245-4260. https://doi.org/10.3390/ijerph10094245
APA StyleTremblay, C. -L., & Archambault, M. (2013). Interference in Pheromone-Responsive Conjugation of a High-Level Bacitracin Resistant Enterococcus faecalis Plasmid of Poultry Origin. International Journal of Environmental Research and Public Health, 10(9), 4245-4260. https://doi.org/10.3390/ijerph10094245