Genetic Diversity of Rotavirus Strains Circulating in Environmental Water and Bivalve Shellfish in Thailand

Abstract
:1. Introduction
2. Experimental Section
2.1. Water and Oyster Samples
2.2. Virus Processing
2.2.1. Water Samples
2.2.2. Oyster Samples
2.3. RNA Extraction and RT-nested PCR
2.4. Phylogenetic Analysis of Rotavirus-positive Samples
3. Results
3.1. Rotavirus Detection in Environmental Samples

{kind=link}
| Source of Samples | No. of Samples | No. of Rotavirus-positive Samples (%) |
|---|---|---|
| River | 59 | 16 (27.1) |
| Irrigation canals | 55 | 5 (9.1) |
| Oysters | 110 | 6 (5.4) |
| Total | 224 | 27 (12.0) |
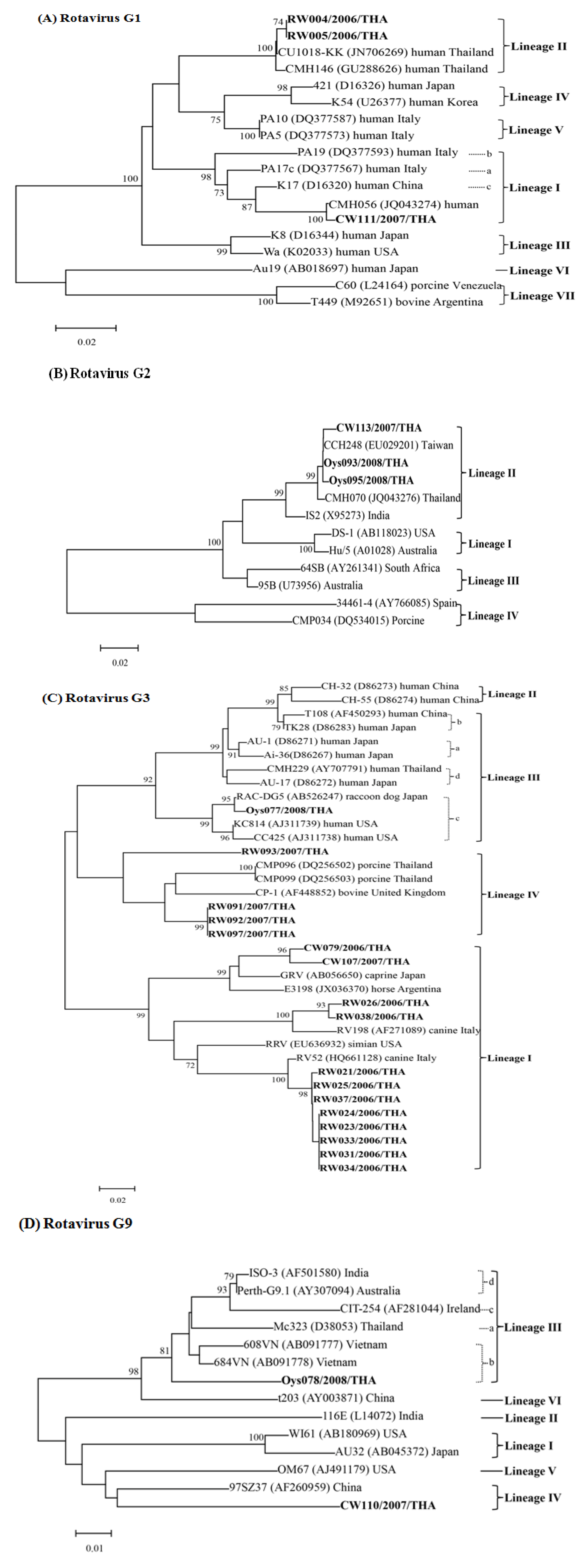
3.2. Molecular Analysis of Rotaviruses

4. Discussion
5. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Steele, A.D.; Duque, J.; Parashar, U.D.; WHO-coordinated Global Rotavirus Surveillance Network. 2008 estimate of worldwide rotavirus-associated mortality in children younger than 5 years before the introduction of universal rotavirus vaccination programmes: A systematic review and meta-analysis. Lancet Infect. Dis. 2012, 12, 136–141. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Rotavirus surveillance—Worldwide, 2009. MMWR 2011, 60, 514–516. [Google Scholar]
- Estes, M.K.; Kapikian, A.Z. Rotaviruses. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Griffin, D.E., Lamb, R.A., Straus, S.E., Martin, M.A., Roizman, B., Eds.; Lippincott, Williams & Wilkins: Philadelphia, PA, USA, 2007; pp. 1917–1974. [Google Scholar]
- Matthijnssens, J.; Otto, P.H.; Ciarlet, M.; Desselberger, U.; van Ranst, M.; Johne, R. VP6-sequence-based cutoff values as a criterion for rotavirus species demarcation. Arch. Virol. 2012, 157, 1177–1182. [Google Scholar] [CrossRef]
- Matthijnssens, J.; Ciarlet, M.; McDonald, S.M.; Attoui, H.; Bányai, K.; Brister, J.R.; Buesa, J.; Esona, M.D.; Estes, M.K.; Gentsch, J.R.; et al. Uniformity of rotavirus strain nomenclature proposed by the Rotavirus Classification Working Group (RCWG). Arch. Virol. 2011, 156, 1397–1413. [Google Scholar] [CrossRef]
- Trojnar, E.; Sachsenröder, J.; Twardziok, S.; Reetz, J.; Otto, P.H.; Johne, R. Identification of an avian group A rotavirus containing a novel VP4 gene with a close relationship to those of mammalian rotaviruses. J. Gen. Virol. 2013, 94, 136–142. [Google Scholar] [CrossRef]
- Bishop, R. Discovery of rotavirus: Implications for child health. J. Gastroenterol. Hepatol. 2009, 24, S81–S85. [Google Scholar] [CrossRef]
- Le Guyader, F.S.; le Saux, J.C.; Ambert-Balay, K.; Krol, J.; Serais, O.; Parnaudeau, S.; Giraudon, H.; Delmas, G.; Pommepuy, M.; Pothier, P.; Atmar, R.L. Aichi virus, norovirus, astrovirus, enterovirus, and rotavirus involved in clinical cases from a French oyster-related gastroenteritis outbreak. J. Clin. Microbiol. 2008, 46, 4011–4017. [Google Scholar] [CrossRef]
- Räsänen, S.; Lappalainen, S.; Kaikkonen, S.; Hämäläinen, M.; Salminen, M.; Vesikari, T. Mixed viral infections causing acute gastroenteritis in children in a waterborne outbreak. Epidemiol. Infect. 2010, 138, 1227–1234. [Google Scholar] [CrossRef]
- Koroglu, M.; Yakupogullari, Y.; Otlu, B.; Ozturk, S.; Ozden, M.; Ozer, A.; Sener, K.; Durmaz, R. A waterborne outbreak of epidemic diarrhea due to group A rotavirus in Malatya, Turkey. New Microbiol. 2011, 34, 17–24. [Google Scholar]
- Mellou, K.; Katsioulis, A.; Potamiti-Komi, M.; Pournaras, S.; Kyritsi, M.; Katsiaflaka, A.; Kallimani, A.; Kokkinos, P.; Petinaki, E.; Sideroglou, T.; Georgakopoulou, T.; Vantarakis, A.; Hadjichristodoulou, C. A large waterborne gastroenteritis outbreak in central Greece, March 2012: Challenges for the investigation and management. Epidemiol. Infect. 2014, 142, 40–50. [Google Scholar]
- Gabrieli, R.; Macaluso, A.; Lanni, L.; Saccares, S.; di Giamberardino, F.; Cencioni, B.; Petrinca, A.R.; Divizia, M. Enteric viruses in molluscan shellfish. New Microbiol. 2007, 30, 471–475. [Google Scholar]
- Hansman, G.S.; Oka, T.; Li, T.C.; Nishio, O.; Noda, M.; Takeda, N. Detection of human enteric viruses in Japanese clams. J. Food Prot. 2008, 71, 1689–1695. [Google Scholar]
- Sdiri-Loulizi, K.; Hassine, M.; Aouni, Z.; Gharbi-Khelifi, H.; Chouchane, S.; Sakly, N.; Neji-Guédiche, M.; Pothier, P.; Aouni, M.; Ambert-Balay, K. Detection and molecular characterization of enteric viruses in environmental samples in Monastir, Tunisia between January 2003 and April 2007. J. Appl. Microbiol. 2010, 109, 1093–1104. [Google Scholar] [CrossRef]
- Rigotto, C.; Victoria, M.; Moresco, V.; Kolesnikovas, C.K.; Corrêa, A.A.; Souza, D.S.; Miagostovich, M.P.; Simões, C.M.; Barardi, C.R. Assessment of adenovirus, hepatitis A virus and rotavirus presence in environmental samples in Florianopolis, south Brazil. J. Appl. Microbiol. 2010, 109, 1979–1987. [Google Scholar] [CrossRef]
- Keller, R.; Justino, J.F.; Cassini, S.T. Assessment of water and seafood microbiology quality in a mangrove region in Vitória, Brazil. J. Water Health 2013, 11, 573–580. [Google Scholar] [CrossRef]
- Bagordo, F.; Grassi, T.; Idolo, A.; Serio, F.; Gabutti, G.; de Donno, A. Rotavirus occurrence in shellfish with low levels of E. coli. Food Environ. Virol. 2013, 5, 169–175. [Google Scholar] [CrossRef]
- Kirkwood, C.D.; Boniface, K.; Barnes, G.L.; Bishop, R.F. Distribution of rotavirus genotypes after introduction of rotavirus vaccines, Rotarix® and RotaTeq®, into the national immunization program of Australia. Pediatr. Infect. Dis. J. 2011, 30, S48–S53. [Google Scholar] [CrossRef]
- McDermid, A.; le Saux, N.; Grudeski, E.; Bettinger, J.A.; Manguiat, K.; Halperin, S.A.; Macdonald, L.; Déry, P.; Embree, J.; Vaudry, W.; Booth, T.F.; Members of the Canadian immunization monitoring program, active. Molecular characterization of rotavirus isolates from select Canadian pediatric hospitals. BMC Infect. Dis. 2012, 12. [Google Scholar] [CrossRef]
- Enweronu-Laryea, C.C.; Sagoe, K.W.; Damanka, S.; Lartey, B.; Armah, G.E. Rotavirus genotypes associated with childhood severe acute diarrhoea in southern Ghana: A cross-sectional study. Virol. J. 2013, 10. [Google Scholar] [CrossRef]
- Khamrin, P.; Peerakome, S.; Tonusin, S.; Malasao, R.; Okitsu, S.; Mizuguchi, M.; Ushijima, H.; Maneekarn, N. Changing pattern of rotavirus G genotype distribution in Chiang Mai, Thailand from 2002 to 2004: Decline of G9 and reemergence of G1 and G2. J. Med. Virol. 2007, 79, 1775–1782. [Google Scholar] [CrossRef]
- Khananurak, K.; Vutithanachot, V.; Simakachorn, N.; Theamboonlers, A.; Chongsrisawat, V.; Poovorawan, Y. Prevalence and phylogenetic analysis of rotavirus genotypes in Thailand between 2007 and 2009. Infect. Genet. Evol. 2010, 10, 537–545. [Google Scholar] [CrossRef]
- Maiklang, O.; Vutithanachot, V.; Vutithanachot, C.; Hacharoen, P.; Chieochansin, T.; Poovorawan, Y. Prevalence of group A genotype human rotavirus among children with diarrhea in Thailand, 2009–2011. Southeast Asian J. Trop. Med. Public Health 2012, 43, 904–916. [Google Scholar]
- Kittigul, L.; Swangsri, T.; Pombubpa, K.; Howteerakul, N.; Diraphat, P.; Hirunpetcharat, C. Rotavirus infection in children and adults with acute gastroenteritis in Thailand. Southeast Asian J. Trop. Med. Public Health 2014, in press. [Google Scholar]
- Kittigul, L.; Khamoun, P.; Sujirarat, D.; Utrarachkij, F.; Chitpirom, K.; Chaichantanakit, N.; Vathanophas, K. An improved method for concentrating rotavirus from water samples. Mem. Inst. Oswaldo Cruz. 2001, 96, 815–821. [Google Scholar] [CrossRef]
- Kittigul, L.; Pombubpa, K.; Rattanatham, T.; Diraphat, P.; Utrarachkij, F.; Pungchitton, S.; Khamrin, P.; Ushijima, H. Development of a method for concentrating and detecting rotavirus in oysters. Int. J. Food Microbiol. 2008, 122, 204–210. [Google Scholar] [CrossRef]
- Gilgen, M.; Germann, D.; Luthy, J.; Hubner, P. Three-step isolation method for sensitive detection of enterovirus, rotavirus, hepatitis A virus, and small round structured viruses in water samples. Int. J. Food Microbiol. 1997, 37, 189–199. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Molec. Biol. 1990, 215, 403–410. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Chaimongkol, N.; Khamrin, P.; Malasao, R.; Thongprachum, A.; Ushijima, H.; Maneekarn, N. Genotypic linkages of gene segments of rotaviruses circulating in pediatric patients with acute gastroenteritis in Thailand. Infect. Genet. Evol. 2012, 12, 1381–1391. [Google Scholar] [CrossRef]
- Than, V.T.; Kang, H.; Lim, I.; Kim, W. Molecular characterization of serotype G9 rotaviruses circulating in South Korea between 2005 and 2010. J. Med. Virol. 2013, 85, 171–178. [Google Scholar] [CrossRef]
- Kittigul, L.; Ekchaloemkiet, S.; Utrarachkij, F.; Siripanichgon, K.; Sujirarat, D.; Pungchitton, S.; Boonthum, A. An efficient virus concentration method and RT-nested PCR for detection of rotaviruses in environmental water samples. J. Virol. Methods 2005, 124, 117–122. [Google Scholar] [CrossRef]
- Steyer, A.; Torkar, K.G.; Gutiérrez-Aguirre, I.; Poljšak-Prijatelj, M. High prevalence of enteric viruses in untreated individual drinking water sources and surface water in Slovenia. Int. J. Hyg. Environ. Health 2011, 214, 392–398. [Google Scholar] [CrossRef]
- Katukiza, A.Y.; Temanu, H.; Chung, J.W.; Foppen, J.W.; Lens, P.N. Genomic copy concentrations of selected waterborne viruses in a slum environment in Kampala, Uganda. J. Water Health 2013, 11, 358–370. [Google Scholar] [CrossRef]
- Le Guyader, F.; Haugarreau, L.; Miossec, L.; Dubois, E.; Pommepuy, M. Three-year study to assess human enteric viruses in shellfish. Appl. Environ. Microbiol. 2000, 66, 3241–3248. [Google Scholar] [CrossRef]
- Kittigul, L.; Pombubpa, K.; Taweekate, Y.; Yeephoo, T.; Khamrin, P.; Ushijima, H. Molecular characterization of rotaviruses, noroviruses, sapovirus, and adenoviruses in patients with acute gastroenteritis in Thailand. J. Med. Virol. 2009, 81, 345–353. [Google Scholar] [CrossRef]
- Afrad, M.H.; Hassan, Z.; Farjana, S.; Moni, S.; Barua, S.; Das, S.K.; Faruque, A.S.; Azim, T.; Rahman, M. Changing profile of rotavirus genotypes in Bangladesh, 2006–2012. BMC Infect. Dis. 2013, 13. [Google Scholar] [CrossRef]
- Matthijnssens, J.; de Grazia, S.; Piessens, J.; Heylen, E.; Zeller, M.; Giammanco, G.M.; Bányai, K.; Buonavoglia, C.; Ciarlet, M.; Martella, V.; van Ranst, M. Multiple reassortment and interspecies transmission events contribute to the diversity of feline, canine and feline/canine-like human group A rotavirus strains. Infect. Genet. Evol. 2011, 11, 1396–1406. [Google Scholar] [CrossRef]
- Okitsu, S.; Khamrin, P.; Thongprachum, A.; Kongkaew, A.; Maneekarn, N.; Mizuguchi, M.; Hayakawa, S.; Ushijima, H. Whole-genomic analysis of G3P[23], G9P[23] and G3P[13] rotavirus strains isolated from piglets with diarrhea in Thailand, 2006–2008. Infect. Genet. Evol. 2013, 18, 74–86. [Google Scholar] [CrossRef]
- Zeller, M.; Patton, J.T.; Heylen, E.; de Coster, S.; Ciarlet, M.; van Ranst, M.; Matthijnssens, J. Genetic analyses reveal differences in the VP7 and VP4 antigenic epitopes between human rotaviruses circulating in Belgium and rotaviruses in Rotarix and RotaTeq. J. Clin. Microbiol. 2012, 50, 966–976. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kittigul, L.; Panjangampatthana, A.; Rupprom, K.; Pombubpa, K. Genetic Diversity of Rotavirus Strains Circulating in Environmental Water and Bivalve Shellfish in Thailand. Int. J. Environ. Res. Public Health 2014, 11, 1299-1311. https://doi.org/10.3390/ijerph110201299
Kittigul L, Panjangampatthana A, Rupprom K, Pombubpa K. Genetic Diversity of Rotavirus Strains Circulating in Environmental Water and Bivalve Shellfish in Thailand. International Journal of Environmental Research and Public Health. 2014; 11(2):1299-1311. https://doi.org/10.3390/ijerph110201299
Chicago/Turabian StyleKittigul, Leera, Apinya Panjangampatthana, Kitwadee Rupprom, and Kannika Pombubpa. 2014. "Genetic Diversity of Rotavirus Strains Circulating in Environmental Water and Bivalve Shellfish in Thailand" International Journal of Environmental Research and Public Health 11, no. 2: 1299-1311. https://doi.org/10.3390/ijerph110201299
APA StyleKittigul, L., Panjangampatthana, A., Rupprom, K., & Pombubpa, K. (2014). Genetic Diversity of Rotavirus Strains Circulating in Environmental Water and Bivalve Shellfish in Thailand. International Journal of Environmental Research and Public Health, 11(2), 1299-1311. https://doi.org/10.3390/ijerph110201299