Effects of Dietary Selenium on Histopathological Changes and T Cells of Spleen in Broilers Exposed to Aflatoxin B1

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chickens and Diets
2.2. Relative Weight of Spleen
2.3. Pathological Observation
2.4. Determination of Splenic T-cell Subsets
2.5. Statistical Analysis
3. Results
3.1. Changes of Relative Weight of Spleen

{kind=link}
| Group | 7 Days | 14 Days | 21 Days |
|---|---|---|---|
| Control group | 0.61 ± 0.06 | 0.55 ± 0.12 | 0.73 ± 0.07 Be |
| AFB1 group | 0.53 ± 0.06 | 0.51 ± 0.10 | 0.57 ± 0.05 AcDE |
| +Se group I | 0.53 ± 0.09 | 0.63 ± 0.11 | 0.66 ± 0.06 bDE |
| +Se group II | 0.53 ± 0.08 | 0.60 ± 0.09 | 0.80 ± 0.05 BC |
| +Se group III | 0.57 ± 0.07 | 0.57 ± 0.10 | 0.81 ± 0.06 aBC |
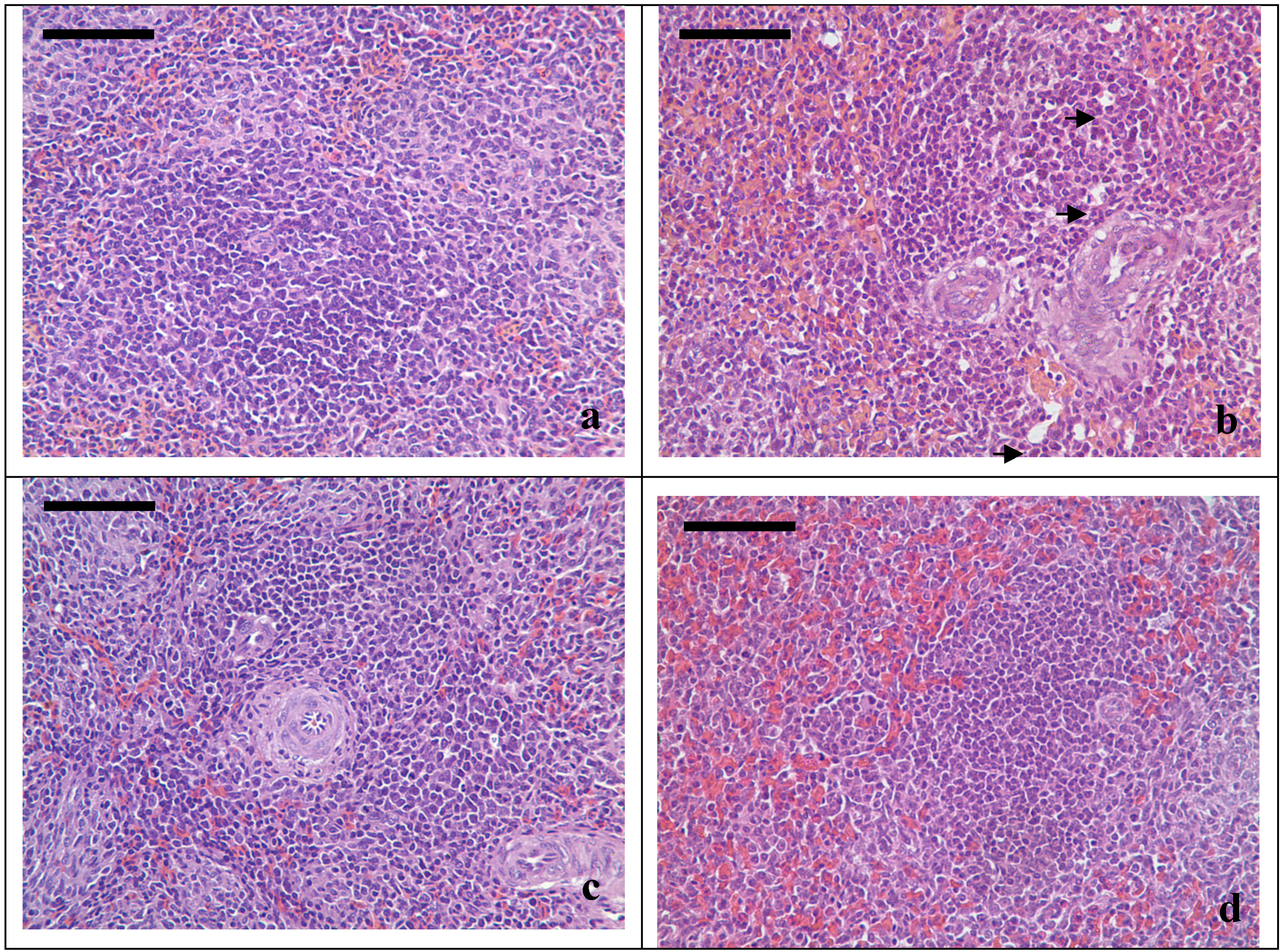
3.2. Pathological Lesions
3.3. Changes of Splenic T-cell Subsets
| Time | Pathological Lesions | Control Group | AFB1 Group | +Se Group I | +Se Group II | +Se Group III |
|---|---|---|---|---|---|---|
| 7 days | Congestion in red pulp | 1/5 | 2/5 | 1/5 | 1/5 | 1/5 |
| Vacuoles appeared | 1/5 | 1/5 | 2/5 | 1/5 | 1/5 | |
| 14 days | Congestion in red pulp | 1/5 | 1/5 | 2/5 | 1/5 | 2/5 |
| Vacuoles appeared | 1/5 | 2/5 | 1/5 | 1/5 | 1/5 | |
| 21 days | Congestion in red pulp | 1/5 | 4/5 | 4/5 | 1/5 | 3/5 |
| Vacuoles appeared | 1/5 | 4/5 | 3/5 | 1/5 | 1/5 |

4. Discussion
| Time | Items | Control Group | AFB1 Group | +Se Group I | +Se Group II | +Se Group III |
|---|---|---|---|---|---|---|
| 7days | CD3+ (%) | 50.81 ± 4.22B CD | 45.55 ± 3.19 ACDE | 58.72 ± 3.36 ABE | 57.90 ± 3.66 ABE | 55.12 ± 3.40 BCD |
| CD3+ CD4+ (%) | 10.11 ± 1.82 | 6.82 ± 1.83 ACDe | 10.60 ± 2.00 b | 11.12 ± 2.02 B | 10.14 ± 2.02 b | |
| CD3+CD8+ (%) | 25.91 ± 2.24 BCD | 19.18 ± 3.39 ACDE | 28.11 ± 2.33 aB | 30.26 ± 2.17 ABe | 26.44 ± 2.65 Bd | |
| CD4+/CD8+ | 0.39 ± 0.04 | 0.35 ± 0.04 | 0.37 ± 0.04 | 0.37 ± 0.04 | 0.38 ± 0.04 | |
| 14days | CD3+ (%) | 54.69 ± 1.97 BCDE | 41.24 ± 2.45 ACDE | 58.84 ± 1.51 aBDE | 66.65 ± 1.88 ABC | 63.66 ± 1.90 ABC |
| CD3+ CD4+ (%) | 12.19 ± 1.62 B | 8.92 ± 1.92 ACDe | 12.24 ± 1.73 B | 13.40 ± 1.72 B | 11.53 ± 1.66 b | |
| CD3+CD8+ (%) | 26.02 ± 1.98 BD | 14.97 ± 1.77 ACDE | 23.60 ± 1.97 BD | 31.31 ± 1.81 ABCE | 25.71 ± 2.40 BD | |
| CD4+/CD8+ | 0.44 ± 0.03 | 0.43 ± 0.06 | 0.43 ± 0.04 | 0.42 ± 0.03 | 0.43 ± 0.04 | |
| 21days | CD3+ (%) | 57.41 ± 1.89 BD | 42.20 ± 1.93 ACDE | 59.27 ± 2.14 BD | 65.51 ± 2.17 ABCE | 60.39 ± 2.02 BD |
| CD3+ CD4+ (%) | 15.34 ± 2.34 BD | 8.61 ± 1.62 ACDE | 13.97 ± 2.09 BD | 18.66 ± 2.03 ABCE | 15.12 ± 2.10 BD | |
| CD3+CD8+ (%) | 29.27 ± 2.23 BD | 16.84 ± 1.99 ACDE | 26.54 ± 2.21 BD | 35.22 ± 2.04 ABCE | 28.92 ± 2.70 BD | |
| CD4+/CD8+ | 0.52 ± 0.04 | 0.51 ± 0.04 | 0.52 ± 0.04 | 0.53 ± 0.03 | 0.52 ± 0.04 |
5. Conclusions
Acknowledgment
Author Contributions
Conflicts of Interest
References
- Some Traditional Herbal Medicines, Some Mycotoxins, Naphthalene and Styrene. Available online: http://monographs.iarc.fr/ENG/Monographs/vol82/mono82.pdf (accessed on 17 December 2013).
- Wogan, G.N. Aflatoxins as risk factors for hepatocellular carcinoma in humans. Cancer Res. 1992, 52, 2114s–2118s. [Google Scholar]
- Verma, J.; Johri, T.S.; Swain, B.K.; Ameena, S. Effect of graded levels of aflatoxin, ochratoxin and their combinations on the performance and immune response of broilers. Br. Poult. Sci. 2004, 45, 512–518. [Google Scholar] [CrossRef]
- Khan, W.A.; Khan, M.Z.; Khan, A.; Hussain, I. Pathological effects of aflatoxin and their amelioration by vitamin E in White Leghorn layers. Pak. Vet. J. 2010, 30, 155–162. [Google Scholar]
- Sumit, R.; Ji, E.K.; Roger, C.J. Aflatoxin B1 in poultry: Toxicology, metabolism and prevention. Res. Vet. Sci. 2010, 89, 325–331. [Google Scholar] [CrossRef]
- Abdel-Wahhab, M.A.; Nada, S.A.; Amra, H.A. Effect of aluminosilicates and bentonite on aflatoxin-induced developmental toxicity in rat. J. Appl. Toxico. 1999, 19, 199–204. [Google Scholar] [CrossRef]
- Hawkes, W.C.; Kelley, D.S.; Taylor, P.C. The effects of dietary selenium on the immune system in healthy men. Boil. Trace Elem. Res. 2001, 81, 189–213. [Google Scholar]
- Kiremidjian-Schumacher, L.; Roy, M.; Wishe, H.I.; Cohen, M.W.; Stotzky, G. Regulation of cellular immune responses by Selenium. Biol. Trace Elem. Res. 1992, 33, 23–35. [Google Scholar] [CrossRef]
- Whanger, P.D. Selenium and its relationship to cancer: An update. Br. J. Nutr. 2004, 91, 11–28. [Google Scholar] [CrossRef]
- Altamura, M.; Caradonna, L.; Amati, L.; Pellegrino, N.M.; Urgesi, G.; Miniello, S. Splenectomy and sepsis: The role of the spleen in the immune-mediated bacterial clearance. Immunopharm. Immunot. 2001, 23, 153–161. [Google Scholar] [CrossRef]
- Hinton, D.M.; Myers, M.J.; Raybourne, R.A.; Francke, C.S.; Sotomayor, R.E.; Shaddock, J.; Warbritton, A.; Chou, M.W. Immunotoxicity of aflatoxin B1 in rats: Effects on lymphocytes and the inflammatory response in a chronic intermittent dosing study. Toxicol. Sci. 2003, 73, 362–377. [Google Scholar] [CrossRef]
- Morris, S.M.; Aidoo, A.; Chen, J.J.; Chou, M.W.; Casciano, D.A. Aflatoxin B1-induced Hprt mutations in splenic lymphocytes of fischer 344 rats: Results of an intermittent feeding trial. Mutat. Res. 1999, 423, 33–38. [Google Scholar] [CrossRef]
- Sabourin, P.J.; Price, J.A.; Casbohm, S.L.; Perry, M.R.; Tuttle, R.S.; Rogers, J.V.; Rowell, K.S.; Estep, J.E.; Sabourin, C.L. Evaluation of acute immunotoxicity of aerosolized aflatoxin B1 in female C57BL/6N mice. J. Immunotoxicol. 2006, 3, 11–20. [Google Scholar] [CrossRef]
- Hoffmann, F.W.; Hashimoto, A.C.; Shafer, L.A.; Dow, S.; Berry, M.J.; Hoffmann, P.R. Dietary selenium modulates activation and differentiation of CD4+ T cells in mice through a mechanism involving cellular free thiols. J. Nutr. 2010, 140, 1155–1161. [Google Scholar]
- Roy, M.; Kiremidjian-Schumacher, L.; Wishe, H.I.; Cohen, M.W.; Stotzky, G. Selenium and immune cell functions. II. Effect on lymphocyte-mediated cytotoxicity. Proc. Soc. Exp. Biol. Med. 1990, 193, 143–148. [Google Scholar] [CrossRef]
- Perozo, F.; Rivera, S. Effect of aflatoxin B1 exposure and selenium supplementation on immune response in broilers. Ind. Vet. J. 2003, 80, 1218–1221. [Google Scholar]
- National Research Council. Nutrient Requirements of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Quist, C.F.; Bounous, D.I.; Kilburn, J.V.; Nettles, V.F.; Wyatt, R.D. The effect of dietary aflatoxin on wild turkey poults. J. Wildlife. Dis. 2000, 36, 436–444. [Google Scholar] [CrossRef]
- Guo, S.N.; Shi, D.Y.; Liao, S.Q.; Su, R.S.; Lin, Y.C.; Pan, J.Q.; Tang, Z.X. Influence of selenium on body weights and immune organ indexes in ducklings intoxicated with aflatoxin B1. Biol. Trace Elem. Res. 2012, 146, 167–170. [Google Scholar] [CrossRef]
- Omar, N.A. Effect of some aflatoxins on a lymphatic organ (spleen) of male albino rats (histopathological study). Egypt. Hosp. Med. 2012, 48, 357–367. [Google Scholar]
- Gisela, F.E.; Walter, G.B.; Tina, K.B. CD4, CD8 and TCR defined T-cell subsets in thymus and spleen of 2- and 7-week old commercial broiler chickens. Vet. Mmunol. Immunopathol. 1998, 62, 339–348. [Google Scholar] [CrossRef]
- Brisbin, J.T.; Zhou, H.; Gong, J.; Sabour, P.; Akbari, M.R.; Haghighi, H.R.; Yu, H.; Clarke, A.; Sarson, A.J.; Sharif, S. Gene expression profiling of chicken lymphoid cells after treatment with Lactobacillus acidophilus cellular components. Dev. Comp. Immunol. 2008, 32, 563–574. [Google Scholar]
- Guo, S.N.; Liao, S.Q.; Su, R.S.; Lin, R.Q.; Chen, Y.Z.; Tang, Z.X.; Wu, H.; Shi, D.Y. Influence of longdan xiegan decoction on body weights and immune organ indexes in ducklings intoxicated with aflatoxin B1. J. Anim. Vet. Adv. 2012, 11, 1162–1165. [Google Scholar] [CrossRef]
- Alpsoy, L.; Yildirim, A.; Agar, G. The antioxidant effects of vitamin A, C, and E on aflatoxin B1-induced oxidative stress in human lymphocytes. Toxicol. Ind. Health 2009, 25, 121–127. [Google Scholar]
- Rainbow, L.; Maxwell, S.M.; Hendrickse, R.G. Ultrastructural changes in murine lymphocytes induced by aflatoxin B1. Mycopathologia 1994, 125, 33–39. [Google Scholar] [CrossRef]
- Amstad, P.; Levy, A.; Emerit, I.; Cerutti, P. Evidence for membrane mediated chromosomal damage by aflatoxin B1 in human lymphocytes. Carcinogenesis 1984, 5, 719–723. [Google Scholar] [CrossRef]
- Groopman, J.D.; Kensler, T.W. The light at the end of the tunnel for chemical- specific biomarkers: Daylight or headlight? Carcinogenesis 1999, 20, 1–11. [Google Scholar] [CrossRef]
- Reddy, R.V.; Sharma, R.P. Effects of aflatoxin B1 on murine lymphocytic functions. Toxicology 1989, 54, 31–44. [Google Scholar] [CrossRef]
- Wang, F.Y.; Shu, G.; Peng, X.; Fang, J.; Chen, K.J.; Cui, H.M.; Chen, Z.L.; Zuo, Z.C.; Deng, J.L.; Geng, Y.; et al. Protective effects of sodium selenite against aflatoxin B1-induced oxidative stress and apoptosis in broiler spleen. Int. J. Environ. Res. Public Health 2013, 10, 2834–2844. [Google Scholar] [CrossRef]
- Conforti, L.; Petrovic, M.; Mohammad, D.; Lee, S.; Ma, Q.; Barone, S.; Filipovich, A.H. Hypoxia regulates expression and activity of Kv1.3 channels in T lymphocytes: A possible role in T cell proliferation. J. Immunol. 2003, 170, 695–702. [Google Scholar]
- Robbins, J.R.; Lee, S.M.; Filipovich, A.H.; Szigligeti, P.; Neumeier, L.; Petrovic, M.; Conforti, L. Hypoxia modulates early events in T cell receptor-mediated activation in human T lymphocytes via Kv1.3 channels. J. Physiol. 2005, 564, 131–143. [Google Scholar]
- Agar, G.; Alpsoy, L.; Bozari, S.; Erturk, F.A.; Yildirim, N. Determination of protective role of selenium against aflatoxin B1-induced DNA damage. Toxicol. Ind. Health 2013, 29, 396–403. [Google Scholar]
- Methenitou, G.; Maravelias, C.; Koutsogeorgopoulou, L.; Athanaselis, S.; Koutselinis, A. Immunomodulative effects of aflatoxins and selenium on human peripheral blood lymphocytes. Vet. Hum. Toxicol. 1996, 38, 274–277. [Google Scholar]
- Peng, X.; Cui, H.M.; Deng, J.L.; Zuo, Z.C.; Lai, W.M. Histological lesion of spleen and inhibition of splenocyte proliferation in broilers fed on diets excess in selenium. Biol. Trace Elem. Res. 2011, 140, 66–72. [Google Scholar] [CrossRef]
- Vega, L.; Rodriguez-Sosa, M.; Garcia-Montalvo, E.A.; Razo, L.M.D.; Elizondo, G. Non-optimallevels of dietary selenomethionine alter splenocyte response and modify oxidative stress markers in female mice. Food Chem. Toxicol. 2007, 145, 1147–1153. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, K.; Peng, X.; Fang, J.; Cui, H.; Zuo, Z.; Deng, J.; Chen, Z.; Geng, Y.; Lai, W.; Tang, L.; et al. Effects of Dietary Selenium on Histopathological Changes and T Cells of Spleen in Broilers Exposed to Aflatoxin B1. Int. J. Environ. Res. Public Health 2014, 11, 1904-1913. https://doi.org/10.3390/ijerph110201904
Chen K, Peng X, Fang J, Cui H, Zuo Z, Deng J, Chen Z, Geng Y, Lai W, Tang L, et al. Effects of Dietary Selenium on Histopathological Changes and T Cells of Spleen in Broilers Exposed to Aflatoxin B1. International Journal of Environmental Research and Public Health. 2014; 11(2):1904-1913. https://doi.org/10.3390/ijerph110201904
Chicago/Turabian StyleChen, Kejie, Xi Peng, Jing Fang, Hengmin Cui, Zhicai Zuo, Junliang Deng, Zhengli Chen, Yi Geng, Weimin Lai, Li Tang, and et al. 2014. "Effects of Dietary Selenium on Histopathological Changes and T Cells of Spleen in Broilers Exposed to Aflatoxin B1" International Journal of Environmental Research and Public Health 11, no. 2: 1904-1913. https://doi.org/10.3390/ijerph110201904
APA StyleChen, K., Peng, X., Fang, J., Cui, H., Zuo, Z., Deng, J., Chen, Z., Geng, Y., Lai, W., Tang, L., & Yang, Q. (2014). Effects of Dietary Selenium on Histopathological Changes and T Cells of Spleen in Broilers Exposed to Aflatoxin B1. International Journal of Environmental Research and Public Health, 11(2), 1904-1913. https://doi.org/10.3390/ijerph110201904