Combination Effects of (Tri)Azole Fungicides on Hormone Production and Xenobiotic Metabolism in a Human Placental Cell Line

Abstract
:1. Introduction
2. Experimental Section

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primer | Reverse Primer | |
|---|---|---|
| 18sRNA | CGGCTACCACATCCACGGAA | GCTGGAATTACCGCGGCT |
| GAPDH | CCACTCCTCCACCTTTGAC | ACCCTGTTGCTGTAGCCA |
| CYP11A1 | GAGACATGGCCACGATGCTA | CCACTTGCACCAGTGTCTTG |
| CYP1A1 | TTTGAGAAGGGCCACATCCG | AGGCCTCCATATAGGGCAGAT |
| CYP17A | CTATGCTCATCCCCCACAAGG | GGATTCAAGAAACGCTCAGGC |
| CYP19 | TCCCTGTGGACTCTAAATTGCC | TGGGAGATGAGGGGTCCAAT |
| CYP1A2 | CGGTGATTGGCAGAGATCGG | GTCCCTCGTTGTGCTGTGG |
| ABCC1 | GATCCGCTCTGGGATTGGAA | GTAGAAGAGGTCTGCCCAGC |
| UGT2B4 | TTCGGGTTGCAGCCCACGAC | TGGGTTTCCCAGCTTCCAGCCT |
| UGT2B17 | AAGCCAAGGGAGCAGCCCTCA | CGCAGGCCAGCAGGAATGCT |
3. Results
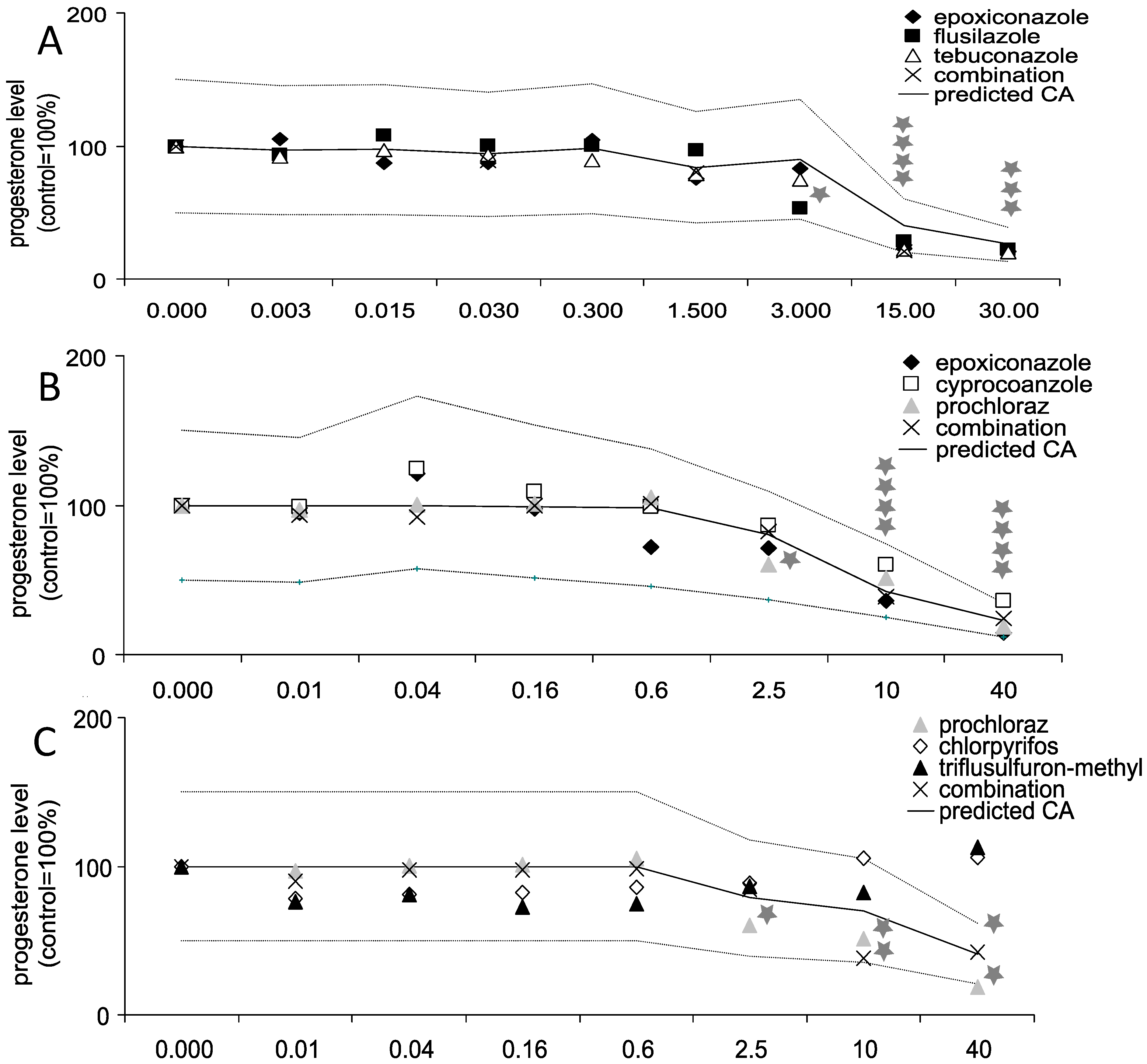
3.1. Effects of Triazole Fungicides Individually and in Combination on Progesterone Production by Jeg-3 Cells
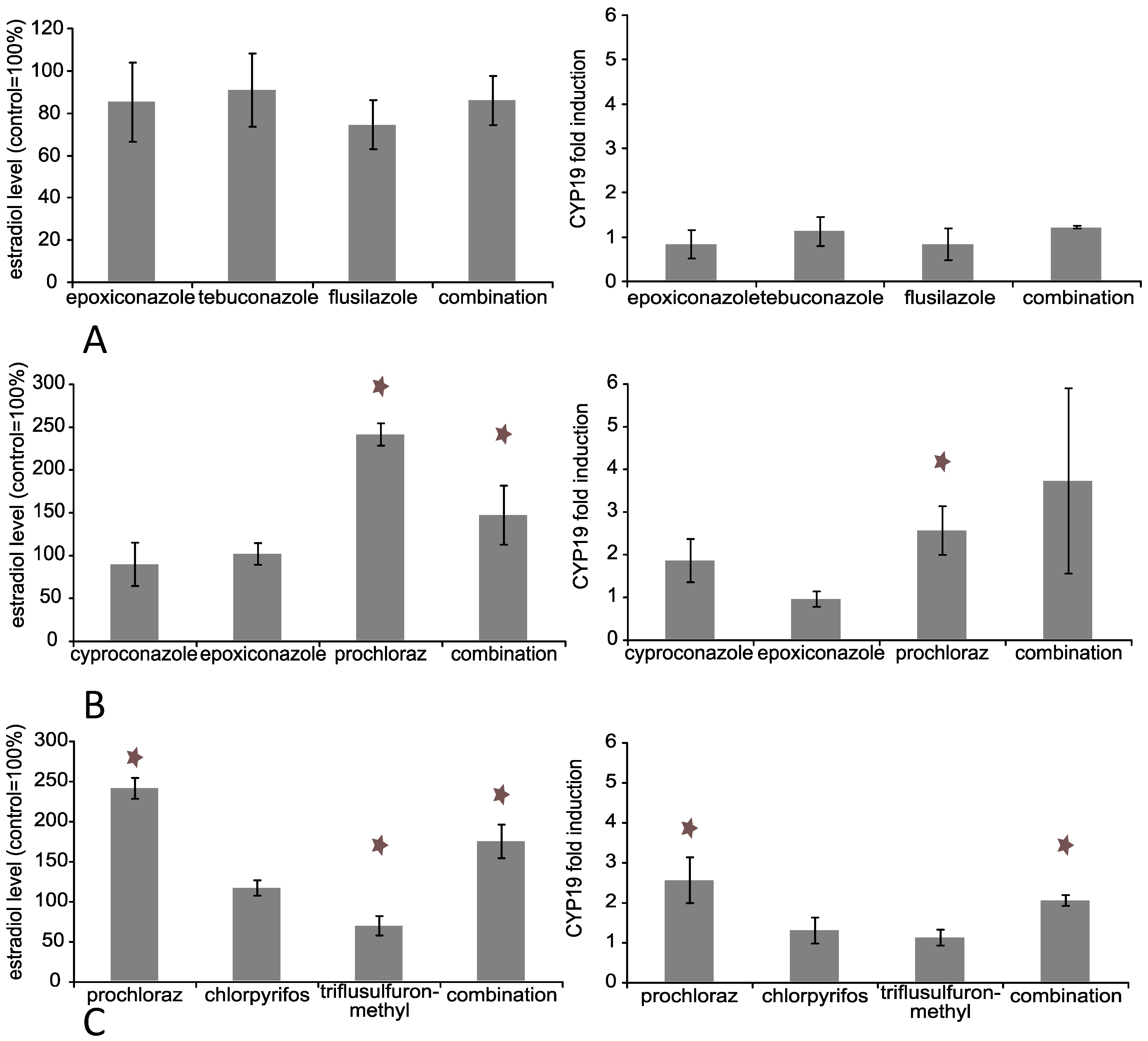
3.2. Effects of Triazole Fungicides Individually and in Combination on Estradiol Production by Jeg-3 Cells


3.3. Effects of Triazole Fungicides Individually and in Combination on Steroid Biosynthesis Dependent Gene Expression

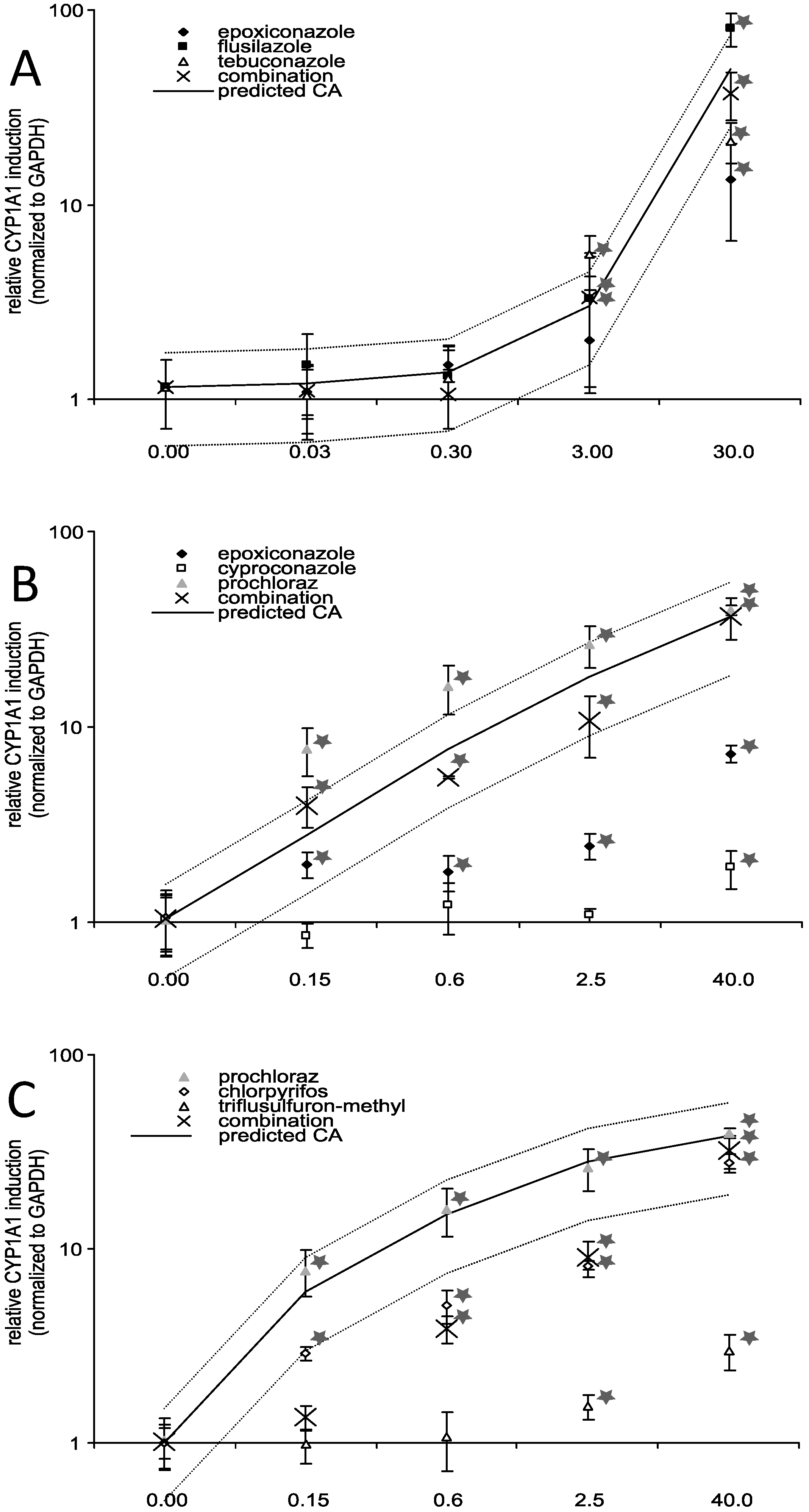
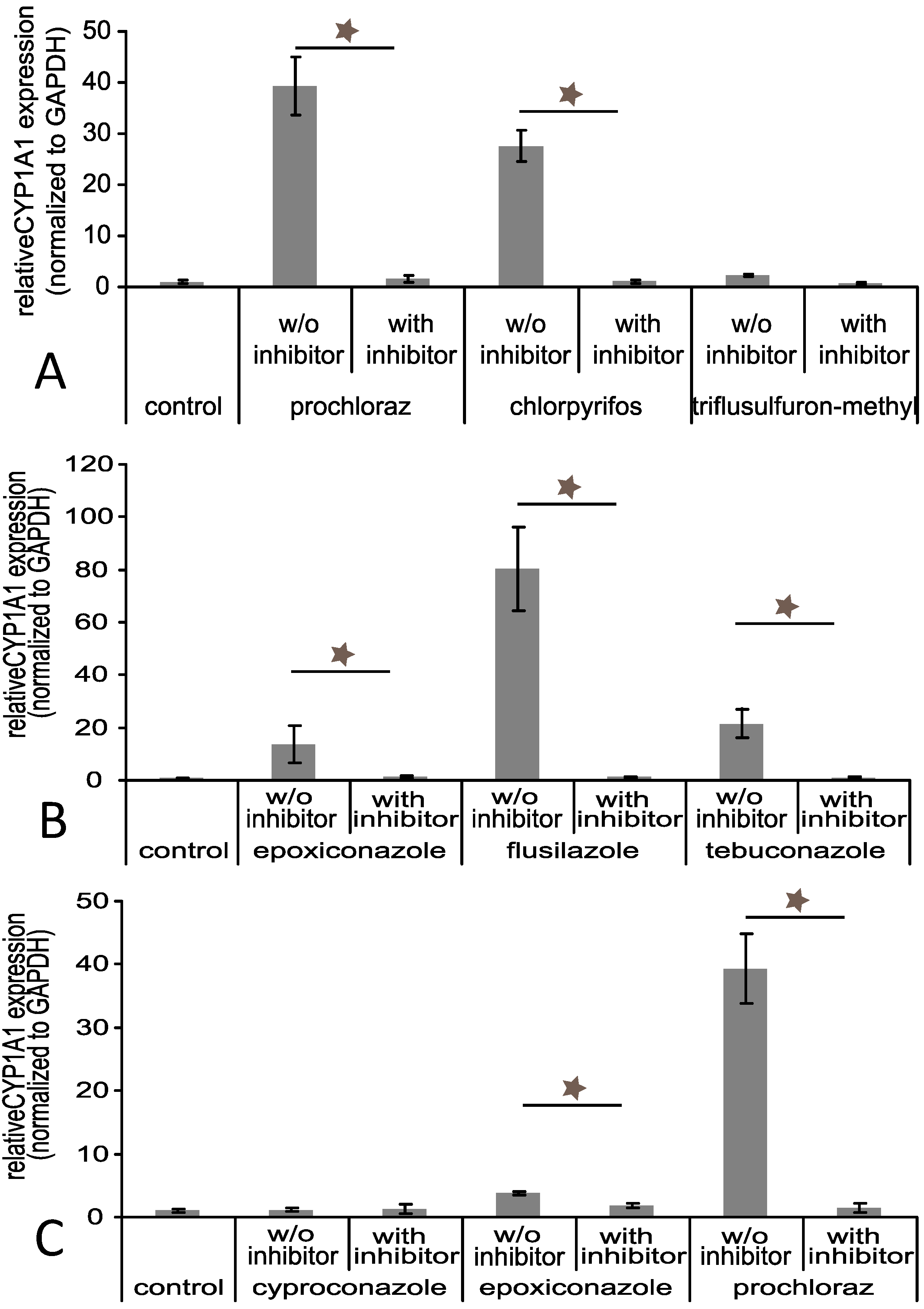
3.4. Effects of Triazole Fungicides Individually and in Combination on Gene Expression of Selected Xenobiotic Metabolising Enzymes
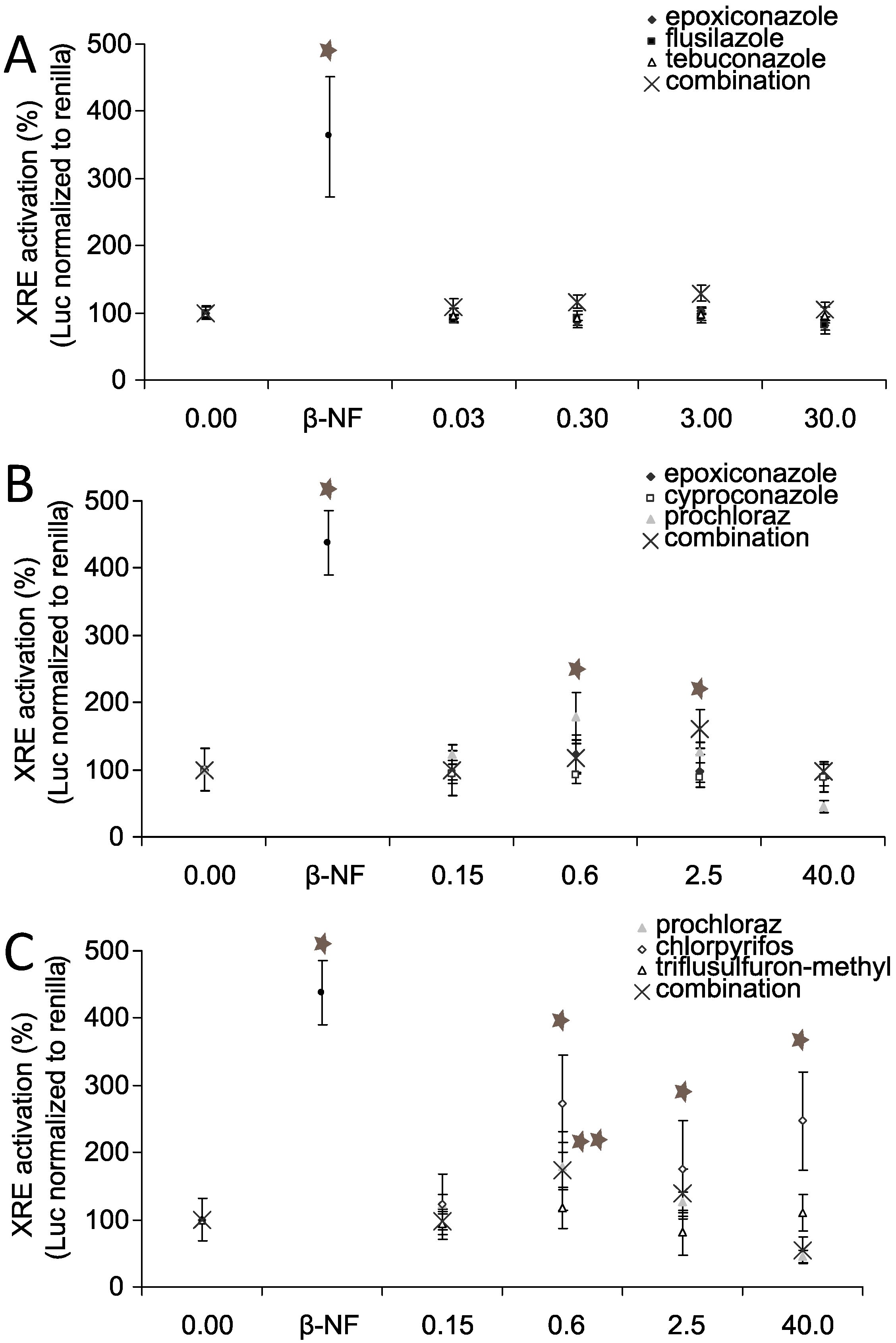
3.5. AhR Activation and AhR Dependent Gene Expression



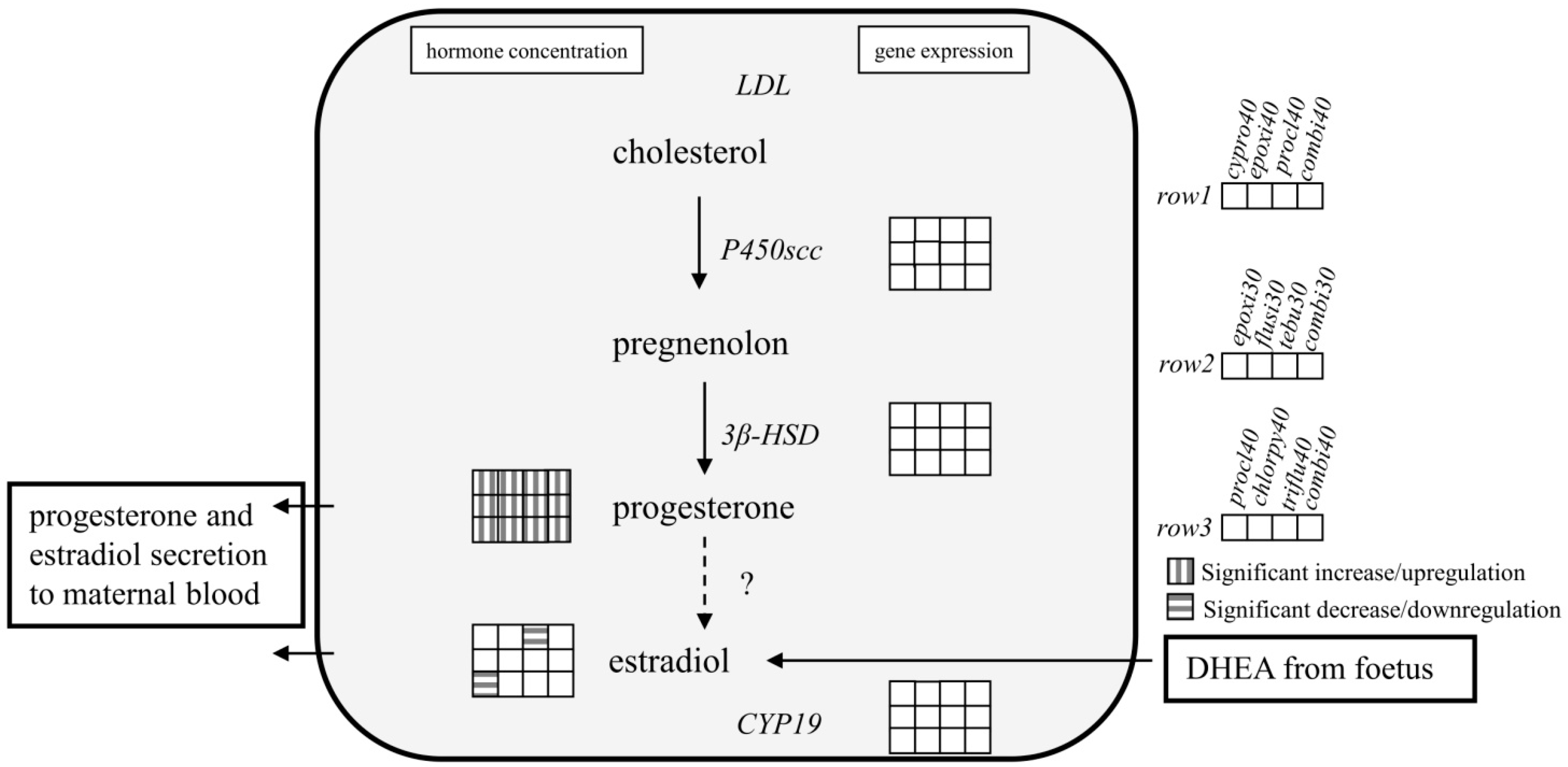
4. Discussion
| Substance Class | Progesterone Synthesis | Endpoint Specific Additive Effect? | CYP1A1 Expression | Endpoint Specific Additive Effect? | |
|---|---|---|---|---|---|
| Cyproconazole | Triazole (DMI-fungicide) | ↓ | Yes | = | No |
| Epoxiconazole | Triazole (DMI-fungicide) | ↓ | Yes | (↑) | Yes |
| Flusilazole | Triazole (DMI-fungicide) | ↓ | Yes | ↑ | Yes |
| Tebuconazole | Triazole (DMI-fungicide) | ↓ | Yes | ↑ | Yes |
| Prochloraz | Imidazole (DMI-fungicide) | ↓ | Yes | ↑ | Yes |
| Chlorpyrifos | Organophosphate- insecticide | = | No | ↑ | Yes |
| Triflusulfuron-methyl | Triazinylsulfonyl-urea-herbicide | = | No | = | No |
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- EFSA. The 2010 European Union Report on pesticide residues in food. EFSA J. 2013, 11. [Google Scholar] [CrossRef]
- Kortenkamp, A.; Faust, M. Combined exposures to anti-androgenic chemicals: Steps towards cumulative risk assessment. Int. J. Androl. 2010, 33, 463–474. [Google Scholar] [CrossRef]
- Hadrup, N.; Taxvig, C.; Pedersen, M.; Nellemann, C.; Hass, U.; Vinggaard, A.M. Concentration addition, independent action and generalized concentration addition models for mixture effect prediction of sex hormone synthesis in vitro. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Hass, U.; Boberg, J.; Christiansen, S.; Jacobsen, P.R.; Vinggaard, A.M.; Taxvig, C.; Poulsen, M.E.; Herrmann, S.S.; Jensen, B.H.; Petersen, A.; et al. Adverse effects on sexual development in rat offspring after low dose exposure to a mixture of endocrine disrupting pesticides. Reprod. Toxicol. 2012, 34, 261–274. [Google Scholar] [CrossRef]
- Jacobsen, P.R.; Christiansen, S.; Boberg, J.; Nellemann, C.; Hass, U. Combined exposure to endocrine disrupting pesticides impairs parturition, causes pup mortality and affects sexual differentiation in rats. Int. J. Androl. 2010, 33, 434–442. [Google Scholar] [CrossRef]
- Kjaerstad, M.B.; Taxvig, C.; Andersen, H.R.; Nellemann, C. Mixture effects of endocrine disrupting compounds in vitro. Int. J. Androl. 2010, 33, 425–433. [Google Scholar] [CrossRef]
- Moser, V.C.; Simmons, J.E.; Gennings, C. Neurotoxicological interactions of a five-pesticide mixture in preweanling rats. Toxicol. Sci. 2006, 92, 235–245. [Google Scholar] [CrossRef]
- Payne, J.; Scholze, M.; Kortenkamp, A. Mixtures of four organochlorines enhance human breast cancer cell proliferation. Environ. Health Perspect. 2001, 109, 391–397. [Google Scholar] [CrossRef]
- Kraemer, W.; Ulrich, S. Modern Crop Protection Compounds. Available online: http://onlinelibrary.wiley.com/doi/10.1002/9783527619580.fmatter/pdf (accessed on 5 February 2008).
- Sanderson, J.T.; Boerma, J.; Lansbergen, G.W.; van den Berg, M. Induction and inhibition of aromatase (CYP19) activity by various classes of pesticides in H295R human adrenocortical carcinoma cells. Toxicol. Appl. Pharmacol. 2002, 182, 44–54. [Google Scholar] [CrossRef]
- Vinggaard, A.M.; Hass, U.; Dalgaard, M.; Andersen, H.R.; Bonefeld-Jorgensen, E.; Christiansen, S.; Laier, P.; Poulsen, M.E. Prochloraz: An imidazole fungicide with multiple mechanisms of action. Int. J. Androl. 2006, 29, 186–192. [Google Scholar] [CrossRef]
- Hass, U.; Scholze, M.; Christiansen, S.; Dalgaard, M.; Vinggaard, A.M.; Axelstad, M.; Metzdorff, S.B.; Kortenkamp, A. Combined exposure to anti-androgens exacerbates disruption of sexual differentiation in the rat. Environ. Health Perspect. 2007, 115, 122–128. [Google Scholar] [CrossRef]
- Prutner, W.; Nicken, P.; Haunhorst, E.; Hamscher, G.; Steinberg, P. Effects of single pesticides and binary pesticide mixtures on estrone production in H295R cells. Arch. Toxicol. 2013, 87, 2201–2214. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion on risk assessment for a selected group of pesticides from the triazole group to test possible methodologies to assess cumulative effects from exposure through food from these pesticides on human health. EFSA J. 2009, 7. [Google Scholar] [CrossRef]
- Schneider, S.; Hofmann, T.; Stinchcombe, S.; Moreno, M.C.; Fegert, I.; Strauss, V.; Groters, S.; Fabian, E.; Thiaener, J.; Fussell, K.C.; et al. Species differences in developmental toxicity of epoxiconazole and its relevance to humans. Birth Defects Res. B Dev. Reprod. Toxicol. 2013, 98, 230–246. [Google Scholar] [CrossRef]
- Taxvig, C.; Hass, U.; Axelstad, M.; Dalgaard, M.; Boberg, J.; Andeasen, H.R.; Vinggaard, A.M. Endocrine-disrupting activities in vivo of the fungicides tebuconazole and epoxiconazole. Toxicol. Sci. 2007, 100, 464–473. [Google Scholar] [CrossRef]
- EFSA. Conclusion on the peer review of epoxiconazole. EFSA Scientific Rep. 2008, 138, 1–80. [Google Scholar]
- Samson, M.; Labrie, F.; Luu-The, V. Specific estradiol biosynthetic pathway in choriocarcinoma (JEG-3) cell line. J. Steroid. Biochem. Mol. Biol. 2009, 116, 154–159. [Google Scholar] [CrossRef]
- Kortenkamp, A.; Backhaus, T.; Faust, M. State of the Art Report on Mixture Toxicity—Final Report, Executive Summary; 070307/2007/485103/ETU/D.1. The School of Pharmacy, University of London (ULSOP): Lomdon, UK, 2009. Available online: http://ec.europa.eu/environment/chemicals/effects/pdf/report_mixture_toxicity.pdf (accessed on 22 December 2009).
- Kjaerstad, M.B.; Taxvig, C.; Nellemann, C.; Vinggaard, A.M.; Andersen, H.R. Endocrine disrupting effects in vitro of conazole antifungals used as pesticides and pharmaceuticals. Reprod. Toxicol. 2010, 30, 573–582. [Google Scholar] [CrossRef]
- Kojima, H.; Katsura, E.; Takeuchi, S.; Niiyama, K.; Kobayashi, K. Screening for Estrogen and Androgen Receptor Activities in 200 Pesticidesby In Vitro Reporter Gene Assays Using Chinese Hamster Ovary Cells. Environ. Health Perspect. 2004, 112, 524–531. [Google Scholar] [CrossRef]
- Mankame, T.; Hokanson, R.; Chowdhary, R.; Busbee, D. Altered gene expression in human cells induced by the agricultural chemical Enable. Toxicol. Ind. Health 2004, 20, 89–102. [Google Scholar] [CrossRef]
- Birkhoj, M.; Nellemann, C.; Jarfelt, K.; Jacobsen, H.; Andersen, H.R.; Dalgaard, M.; Vinggaard, A.M. The combined antiandrogenic effects of five commonly used pesticides. Toxicol. Appl. Pharmacol. 2004, 201, 10–20. [Google Scholar] [CrossRef]
- Christiansen, S.; Scholze, M.; Axelstad, M.; Boberg, J.; Kortenkamp, A.; Hass, U. Combined exposure to anti-androgens causes markedly increased frequencies of hypospadias in the rat. Int. J. Androl. 2008, 31, 241–248. [Google Scholar] [CrossRef]
- Kortenkamp, A.; Faust, M. Combined exposures to anti-androgenic chemicals: Steps towards cumulative risk assessment. Int. J. Androl. 2010, 33, 463–474. [Google Scholar] [CrossRef]
- Pottinger, T.G.; Katsiadaki, I.; Jolly, C.; Sanders, M.; Mayer, I.; Scott, A.P.; Morris, S.; Kortenkamp, A.; Scholze, M. Anti-androgens act jointly in suppressing spiggin concentrations in androgen-primed female three-spined sticklebacks—Prediction of combined effects by concentration addition. Aquat. Toxicol. 2013, 140–141, 145–156. [Google Scholar]
- Jiang, Y.Z.; Wang, K.; Fang, R.; Zheng, J. Expression of aryl hydrocarbon receptor in human placentas and fetal tissues. J. Histochem. Cytochem. 2010, 58, 679–685. [Google Scholar] [CrossRef]
- Abbott, B.D.; Schmid, J.E.; Pitt, J.A.; Buckalew, A.R.; Wood, C.R.; Held, G.A.; Diliberto, J.J. Adverse reproductive outcomes in the transgenic Ah receptor-deficient mouse. Toxicol. Appl. Pharmacol. 1999, 155, 62–70. [Google Scholar] [CrossRef]
- Baba, T.; Mimura, J.; Nakamura, N.; Harada, N.; Yamamoto, M.; Morohashi, K.; Fujii-Kuriyama, Y. Intrinsic function of the aryl hydrocarbon (dioxin) receptor as a key factor in female reproduction. Mol. Cell. Biol. 2005, 25, 10040–10051. [Google Scholar] [CrossRef]
- Benedict, J.C.; Lin, T.M.; Loeffler, I.K.; Peterson, R.E.; Flaws, J.A. Physiological role of the aryl hydrocarbon receptor in mouse ovary development. Toxicol. Sci. 2000, 56, 382–388. [Google Scholar] [CrossRef]
- Fernandez-Salguero, P.M.; Ward, J.M.; Sundberg, J.P.; Gonzalez, F.J. Lesions of aryl-hydrocarbon receptor-deficient mice. Vet. Pathol. 1997, 34, 605–614. [Google Scholar]
- Detmar, J.; Rennie, M.Y.; Whiteley, K.J.; Qu, D.; Taniuchi, Y.; Shang, X.; Casper, R.F.; Adamson, S.L.; Sled, J.G.; Jurisicova, A. Fetal growth restriction triggered by polycyclic aromatic hydrocarbons is associated with altered placental vasculature and AhR-dependent changes in cell death. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E519–E530. [Google Scholar]
- Wu, Y.; Chen, X.; Zhou, Q.; He, Q.; Kang, J.; Zheng, J.; Wang, K.; Duan, T. ITE and TCDD differentially regulate the vascular remodeling of rat placenta via the activation of AhR. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Korashy, H.M.; Shayeganpour, A.; Brocks, D.R.; El-Kadi, A.O. Induction of cytochrome P450 1A1 by ketoconazole and itraconazole but not fluconazole in murine and human hepatoma cell lines. Toxicol. Sci. 2007, 97, 32–43. [Google Scholar] [CrossRef]
- Medjakovic, S.; Zoechling, A.; Gerster, P.; Ivanova, M.M.; Teng, Y.; Klinge, C.M.; Schildberger, B.; Gartner, M.; Jungbauer, A. Effect of nonpersistent pesticides on estrogen receptor, androgen receptor, and aryl hydrocarbon receptor. Environ. Toxicol. 2013. [Google Scholar] [CrossRef]
- Nishi, K.; Hundal, S.S. Chlorpyrifos induced toxicity in reproductive organs of female Wistar rats. Food Chem. Toxicol. 2013, 62, 732–738. [Google Scholar] [CrossRef]
- Meyer, A.; Seidler, F.J.; Slotkin, T.A. Developmental effects of chlorpyrifos extend beyond neurotoxicity: Critical periods for immediate and delayed-onset effects on cardiac and hepatic cell signaling. Environ. Health Perspect. 2004, 112, 170–178. [Google Scholar] [CrossRef]
- Tian, Y.; Ishikawa, H.; Yamaguchi, T.; Yamauchi, T.; Yokoyama, K. Teratogenicity and developmental toxicity of chlorpyrifos. Maternal exposure during organogenesis in mice. Reprod. Toxicol. 2005, 20, 267–270. [Google Scholar]
- EFSA. Conclusion regarding the peer review of the risk assessment of the active substance triflusulfuron. Available online: http://www.efsa.europa.eu/en/efsajournal/pub/195r.htm (accessed on 17 April 2009).
- EFSA Panel on Plant Protection Products and their Residues (PPR). Scientific Opinion on the identification of pesticides to be included in cumulative assessment groups on the basis of their toxicological profile. EFSA J. 2013, 11. [Google Scholar] [CrossRef]
- Belden, J.B.; Gilliom, R.J.; Lydy, M.J. How well can we predict the toxicity of pesticide mixtures to aquatic life? Integr. Environ. Assess. Manag. 2007, 3, 364–372. [Google Scholar] [CrossRef]
- Goetz, A.K.; Rockett, J.C.; Ren, H.; Thillainadarajah, I.; Dix, D.J. Inhibition of rat and human steroidogenesis by triazole antifungals. Syst. Biol. Reprod. Med. 2009, 55, 214–226. [Google Scholar] [CrossRef]
- Escobar, J.C.; Patel, S.S.; Beshay, V.E.; Suzuki, T.; Carr, B.R. The human placenta expresses CYP17 and generates androgens de novo. J. Clin. Endocrinol. Metab. 2011, 96, 1385–1392. [Google Scholar] [CrossRef]
- Escobar, J.C.; Carr, B.R. The protein kinase a pathway regulates CYP17 expression and androgen production in the human placenta. J. Clin. Endocrinol. Metab. 2011, 96, 2869–2873. [Google Scholar] [CrossRef]
- Stinchcombe, S.; Schneider, S.; Fegert, I.; Rey Moreno, M.C.; Strauss, V.; Groters, S.; Fabian, E.; Fussell, K.C.; Pigott, G.H.; van Ravenzwaay, B. Effects of estrogen coadministration on epoxiconazole toxicity in rats. Birth Defects Res. B Dev. Reprod. Toxicol. 2013, 98, 247–259. [Google Scholar] [CrossRef]
- Buse, E.; Haeger, J.D.; Svensson-Arvelund, J.; Markert, U.R.; Faas, M.M.; Ernerudh, J.; Dixon, D.; Cline, J.M.; Pfarrer, C. The placenta in toxicology. Part I: Animal models in toxicology: Placental morphology and tolerance molecules in the cynomolgus monkey (Macaca fascicularis). Toxicol. Pathol. 2014, 42, 314–326. [Google Scholar]
- Myllynen, P.; Pasanen, M.; Pelkonen, O. Human placenta: A human organ for developmental toxicology research and biomonitoring. Placenta 2005, 26, 361–371. [Google Scholar] [CrossRef]
- Albrecht, E.D.; Pepe, G.J. Placental steroid hormone biosynthesis in primate pregnancy. Endocr. Rev. 1990, 11, 124–150. [Google Scholar] [CrossRef]
- Strauss, J.F., III; Martinez, F.; Kiriakidou, M. Placental steroid hormone synthesis: Unique features and unanswered questions. Biol. Reprod. 1996, 54, 303–311. [Google Scholar]
- Stejskalova, L.; Vecerova, L.; Perez, L.M.; Vrzal, R.; Dvorak, Z.; Nachtigal, P.; Pavek, P. Aryl hydrocarbon receptor and aryl hydrocarbon nuclear translocator expression in human and rat placentas and transcription activity in human trophoblast cultures. Toxicol. Sci. 2011, 123, 26–36. [Google Scholar] [CrossRef]
- Stejskalova, L.; Pavek, P. The function of cytochrome P450 1A1 enzyme (CYP1A1) and aryl hydrocarbon receptor (AhR) in the placenta. Curr. Pharm. Biotechnol. 2011, 12, 715–730. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rieke, S.; Koehn, S.; Hirsch-Ernst, K.; Pfeil, R.; Kneuer, C.; Marx-Stoelting, P. Combination Effects of (Tri)Azole Fungicides on Hormone Production and Xenobiotic Metabolism in a Human Placental Cell Line. Int. J. Environ. Res. Public Health 2014, 11, 9660-9679. https://doi.org/10.3390/ijerph110909660
Rieke S, Koehn S, Hirsch-Ernst K, Pfeil R, Kneuer C, Marx-Stoelting P. Combination Effects of (Tri)Azole Fungicides on Hormone Production and Xenobiotic Metabolism in a Human Placental Cell Line. International Journal of Environmental Research and Public Health. 2014; 11(9):9660-9679. https://doi.org/10.3390/ijerph110909660
Chicago/Turabian StyleRieke, Svenja, Sophie Koehn, Karen Hirsch-Ernst, Rudolf Pfeil, Carsten Kneuer, and Philip Marx-Stoelting. 2014. "Combination Effects of (Tri)Azole Fungicides on Hormone Production and Xenobiotic Metabolism in a Human Placental Cell Line" International Journal of Environmental Research and Public Health 11, no. 9: 9660-9679. https://doi.org/10.3390/ijerph110909660
APA StyleRieke, S., Koehn, S., Hirsch-Ernst, K., Pfeil, R., Kneuer, C., & Marx-Stoelting, P. (2014). Combination Effects of (Tri)Azole Fungicides on Hormone Production and Xenobiotic Metabolism in a Human Placental Cell Line. International Journal of Environmental Research and Public Health, 11(9), 9660-9679. https://doi.org/10.3390/ijerph110909660