
Isolation and Molecular Characterization of Free-Living Amoebae from Different Water Sources in Italy


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
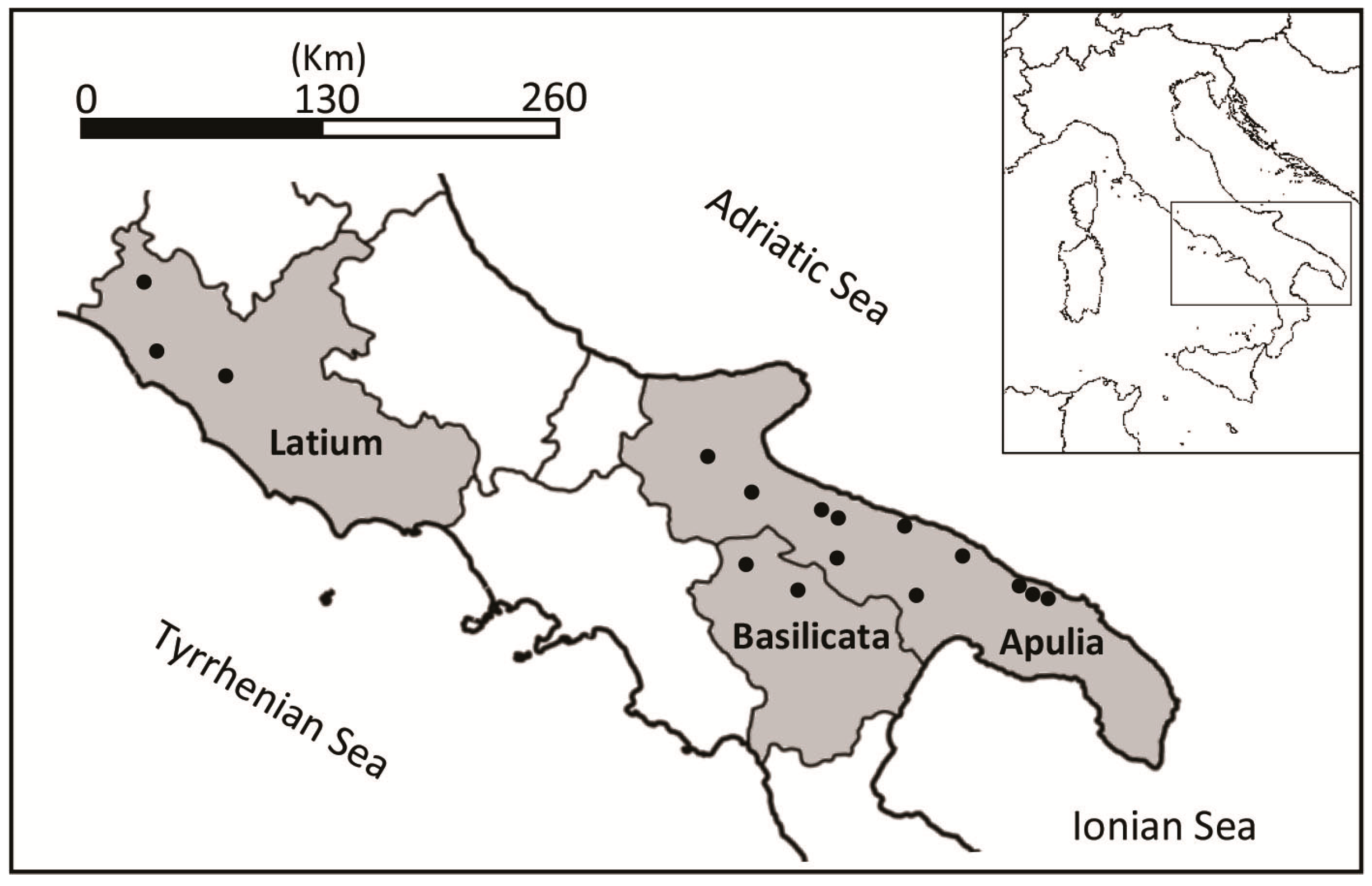
2.1. Sample Collection and Culture of FLA


{kind=link}
{kind=link}
{kind=link}
| Water Samples * | Samples Examined | Cultured Positive Samples | Molecular Identification | |
|---|---|---|---|---|
| Total | Number (%) | Acanthamoeba (Genotype) | V. vermiformis | |
| Well water (W) | 4 | 2 (50.0) | 2 (T15) | - |
| Thermal water (T) | 9 | 3 (33.3) | 2 (T4) | - |
| Tap water (TW) | 84 | 15 (17.8) | 4 (T4) | 8 |
| Bottled mineral water (M) | 19 | 3 (15.8) | - | 3 |
| Ornamental fountain water (F) | 37 | 17 (45.9) | 6 (T4) | 10 |
| Groundwater (G) | 7 | 6 (85.7) | 1 (T4); 3 (T15) | - |
| Total | 160 | 46 (28.7) | 18 (13 T4; 5 T15) | 21 |
2.2. Molecular Methods
3. Results
3.1. Microscopy
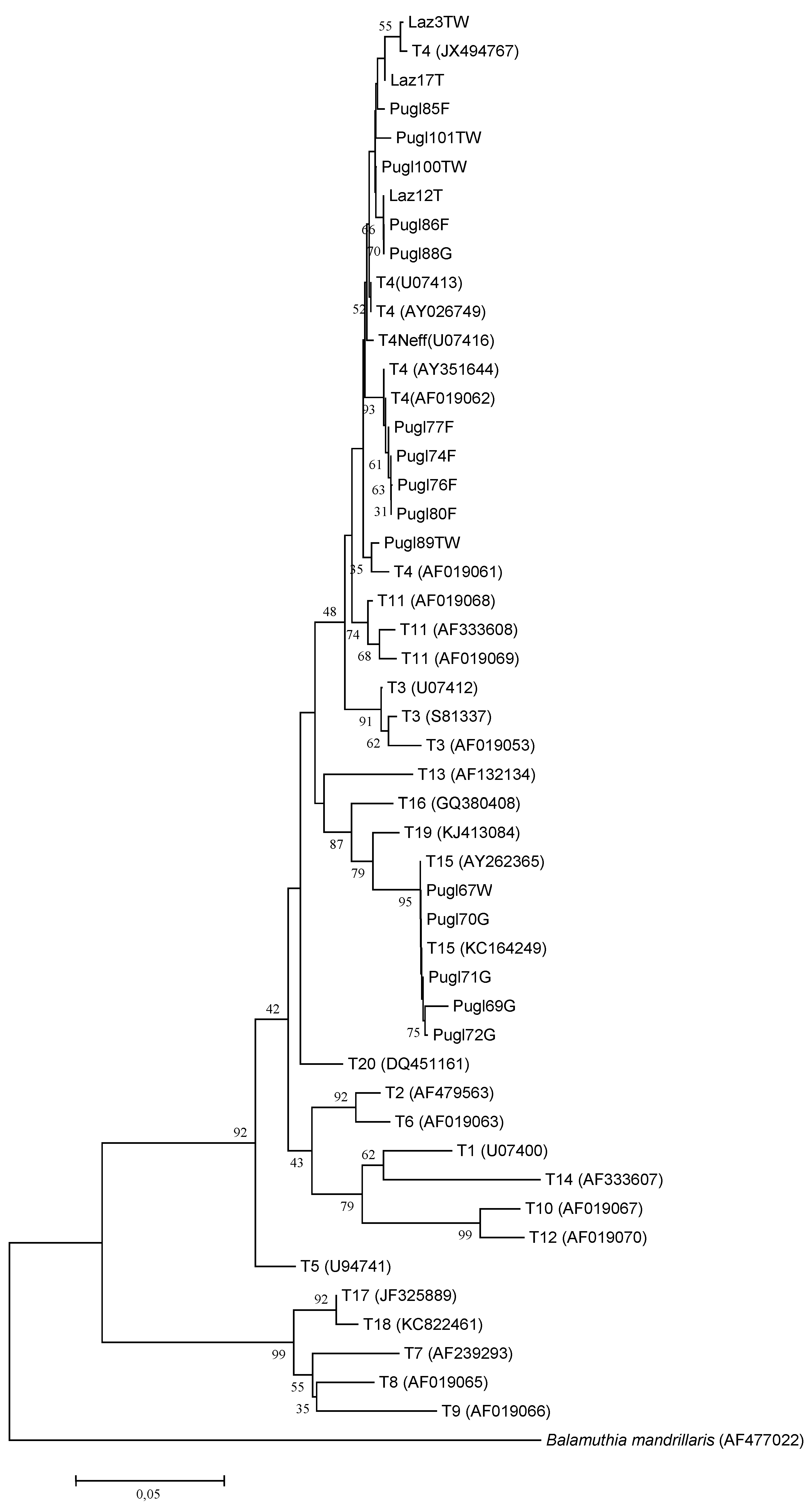
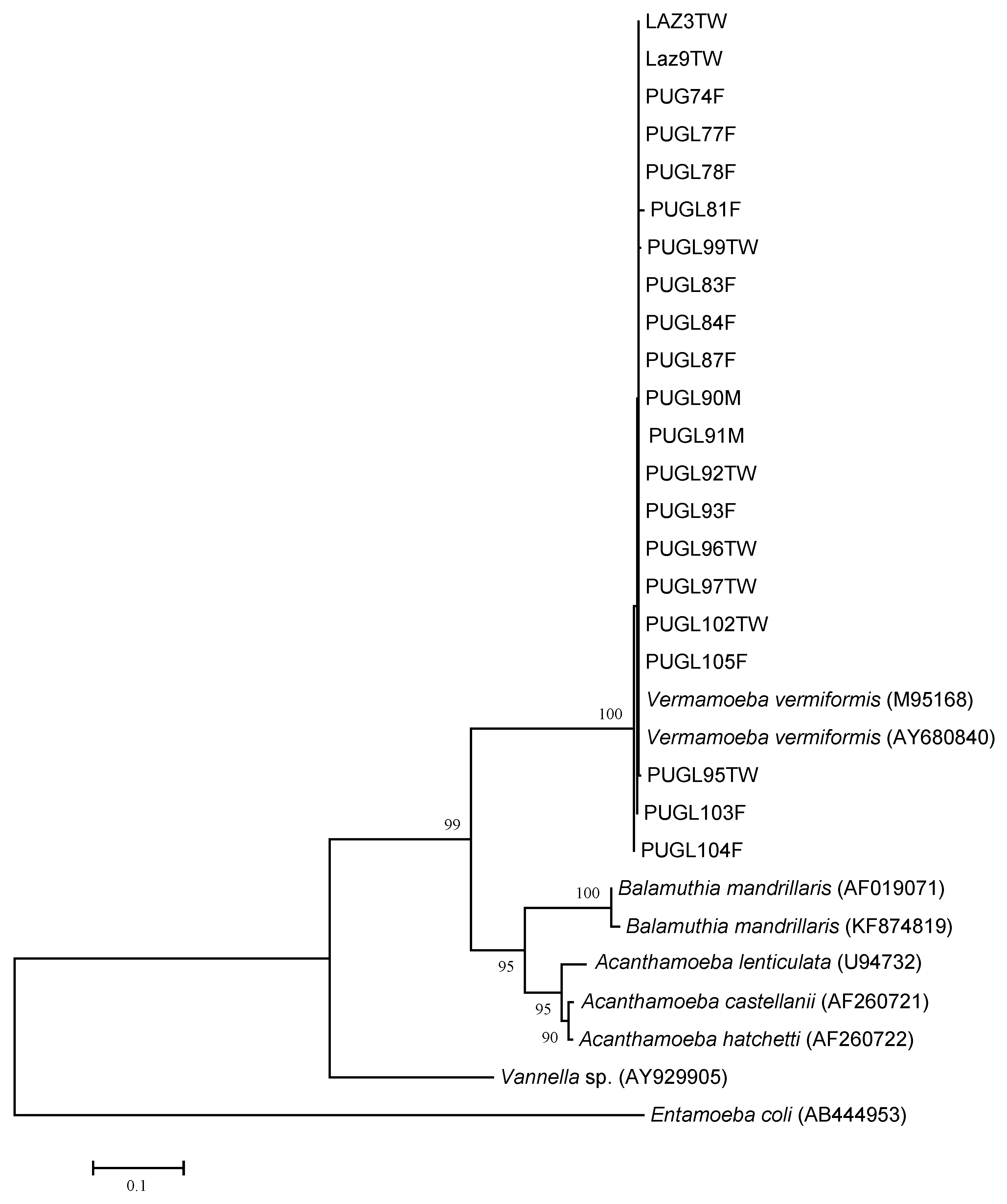
3.2. Molecular Identification and Genotyping


4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Visvesvara, G.S.; Moura, H.; Schuster, F.L. Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immunol. Med. Microbiol. 2007, 50, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, A.V.; Chao, E.; Nassonova, E.S.; Cavalier-Smith, T. A Revised Classification of Naked Lobose Amoebae (Amoebozoa: Lobosa). Protist 2011, 162, 545–570. [Google Scholar] [CrossRef] [PubMed]
- Trabelsi, H.; Dendana, F.; Sellami, A.; Sellami, H.; Cheikhrouhou, F.; Neji, S.; Makni, F.; Ayadi, A. Pathogenic free-living amoebae: Epidemiology and clinical review. Pathol. Biologie 2012, 60, 1–7. [Google Scholar] [CrossRef]
- Fuerst, P.A.; Booton, G.C.; Crary, M. Phylogenetic analysis and the evolution of the 18S rRNA gene typing system of Acanthamoeba. J. Eukaryot. Microbiol. 2015, 62, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Maciver, S.K.; Asif, M.; Simmen, M.W.; Lorenzo-Morales, J. A systematic analysis of Acanthamoeba genotype frequency correlated with source and pathogenicity: T4 is confirmed as a pathogen-rich genotype. Eur. J. Protistol. 2013, 49, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Magnet, A.; Henriques-Gil, N.; Galván-Diaz, A.L.; Izquiedo, F.; Fenoy, S.; del Aguila, C. Novel Acanthamoeba 18S rRNA gene sequence type from an environmental isolate. Parasitol. Res. 2014, 113, 2845–2850. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, S.M.; Devine, P.; Hurley, C.; Ooi, Y.S.; Collum, L.M.T. Corneal infection associated with Hartmannella vermiformis in contact-lens wearer. Lancet 1995, 346, 637–638. [Google Scholar] [CrossRef] [PubMed]
- Niyyati, M.; Rahimi, F.; Lasejerdi, Z.; Rezaeian, M. Potentially pathogenic free-living amoebae in contact lenses of the asymptomatic contact lens wearers. Iranian J. Parasitol. 2014, 9, 14–19. [Google Scholar]
- Scaglia, M.; Strosselli, M.; Grazioli, V.; Gatti, S.; Bernuzzi, A.M.; de Jonckheere, J.F. Isolation and identification of pathogenic Naegleria australiensis (Amoebida, Vahlkampfiidae) from a Spa in northern Italy. Appl. Environ. Microbiol. 1983, 46, 1282–1285. [Google Scholar] [PubMed]
- Scaglia, M.; Gatti, S.; Brustia, R.; Strosselli, M.; Bernuzzi, A.M.; Cevini, C. Pathogenic and non-pathogenic Naegleria and Acanthamoeba spp.: A new autochthonous isolate from an Italian thermal area. Microbiologica 1987, 10, 171–182. [Google Scholar] [PubMed]
- Corsaro, D.; Venditti, D. Phylogenetic evidence for a new genotype of Acanthamoeba (Amoebozoa, Acanthamoebida). Parasitol. Res. 2010, 107, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, J.M.; Booton, J.C.; Hay, J.; Niszl, I.A.; Seal, D.V.; Markus, M.B.; Fuerst, P.A.; Byers, T.J. Use of subgenic 18S ribosomal DNA PCR and sequencing for genus and genotype identification of acanthamoebae from humans with keratitis and from sewage sludge. J. Clin. Microbiol. 2001, 39, 1903–1911. [Google Scholar] [CrossRef] [PubMed]
- Tsvetkova, N.; Schild, M.; Panaiotov, S.; Kurdova-Mintcheva, R.; Gottstein, B.; Walochnik, J.; Aspock, H.; Lucas, M.S.; Muller, N. The identification of free-living environmental isolates of amoebae from Bulgaria. Parasitol. Res. 2004, 92, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Posada, D. Selection of models of DNA evolution with jModelTest. Method. Mol. Biol. 2009, 537, 93–112. [Google Scholar]
- Corsaro, D.; Venditti, D. Molecular phylogenetics evidence for a novel lineage of amoebae within Discosea (Amoebozoa: Lobosa). Acta Protozool. 2013, 52, 309–316. [Google Scholar]
- Coşkun, K.A.; Özçelik, S.; Tutar, L.; Elaldı, N.; Tutar, Y. Isolation and identification of free-living amoebae from tap water in Sivas, Turkey. BioMed Res. Int. 2013, 1–8. [Google Scholar] [CrossRef]
- Garcia, A.; Goñi, P.; Cieloszyk, J.; Fernandez, M.T.; Calvo-Beguería, L.; Rubio, E.; Fillat, M.F.; Peleato, M.L.; Clavel, A. Identification of free-living amoebae and amoeba-associated bacteria from reservoirs and water treatment plants by molecular techniques. Environ. Sci. Technol. 2013, 47, 3132–3140. [Google Scholar] [CrossRef] [PubMed]
- Magnet, A.; Fenoy, S.; Galvàn, A.L.; Izquierdo, F.; Rueda, C.; Fernandez Vadillo, C.; del Aguila, C. A year long study of the presence of free living amoeba in Spain. Water Res. 2013, 47, 6966–6972. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Visvesvara, G.S.; Martinez, A.J.; Theodore, F.H.; Daggett, P.M.; Sawyer, T.K. Naegleria and Acanthamoeba infections: Review. Rev. Infect. Dis. 1990, 12, 490–513. [Google Scholar] [CrossRef] [PubMed]
- Martinez, A.J.; Visvesvara, G.S. Free-living, amphizoic and opportunistic amebas. Brain Pathol. 1997, 7, 583–598. [Google Scholar] [CrossRef] [PubMed]
- Thomas, V.; McDonnell, G.; Denyer, S.P.; Maillard, J.Y. Free-living amoebae and their intracellular pathogenic microorganisms: Risks for water quality. FEMS Microbiol. Rev. 2010, 34, 231–259. [Google Scholar] [CrossRef] [PubMed]
- Goñi, P.; Fernández, M.T.; Rubio, E. Identifying endosymbiont bacteria associated with free-living amoebae. Environ. Microbiol. 2014, 16, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Scheid, P. Relevance of free-living amoebae as hosts for phylogenetically diverse microorganisms. Parasitol. Res. 2014, 113, 2407–2414. [Google Scholar] [CrossRef] [PubMed]
- Cogo, P.E.; Scagli, M.; Gatti, S.; Rossetti, F.; Alaggio, R.; Laverda, A.M.; Zhou, L.; Xiao, L.; Visvesvara, G.S. Fatal Naegleria fowleri meningoencephalitis, Italy. Emerg. Infect. Dis. 2004, 10, 1835–1837. [Google Scholar] [CrossRef]
- Gianinazzi, C.; Schild, M.; Zumkehr, B.; Wüthrich, F.; Nüesch, I.; Ryter, R.; Schürch, N.; Gottstein, B.; Müller, N. Screening of Swiss hot spring resorts for potentially pathogenic free-living amoebae. Exp. Parasitol. 2010, 126, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Loret, J.F.; Greub, G. Free-living amoebae: Biological by-passes in water treatment. Int. J. Hyg. Environ. Health. 2010, 213, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Magnet, A.; Galvan, A.L.; Fenoy, S.; Izquierdo, F.; Rueda, C.; Fernandez Vadillo, C.; Perez-Irezabal, J.; Bandyopadhyay, K.; Visversvara, G.S.; da Silva, A.J.; del Aquila, C. Molecular characterization of Acanthamoeba isolated in water treatment plants and comparison with clinical isolates. Parasitol. Res. 2012, 111, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Hewett, M.K.; Robinson, B.S.; Monis, P.T.; Saint, C.P. Identification of a new Acanthamoeba 18S rRNA gene sequence type, corresponding to the species Acanthamoeba jacobsi Sawyer, Nerad and Visvesvara, 1992 (Lobosea: Acanthamoebidae). Acta Protozool. 2003, 42, 325–329. [Google Scholar]
- Di Cave, D.; Monno, R.; Bottalico, P.; Guerriero, S.; D’Amelio, S.; D’Orazi, C.; Berrilli, F. Acanthamoeba T4 and T15 genotypes associated with keratitis infections in Italy. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 607–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatti, S.; Rama, P.; Matuska, S.; Berrilli, F.; Cavallero, A.; Carletti, S.; Bruno, A.; Maserati, R.; di Cave, D. Isolation and genotyping of Acanthamoeba strains from corneal infections in Italy. J. Med. Microbiol. 2010, 59, 1324–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Cave, D.; D’ Alfonso, R.; Dussey Comlavi, K.A.; D’Orazi, C.; Monno, R.; Berrilli, F. Genotypic heterogeneity based on 18S-rRNA gene sequences among Acanthamoeba isolates from clinical samples in Italy. Exp. Parasitol. 2014. [Google Scholar] [CrossRef]
- Thomas, J.M.; Ashbolt, N.J. Do free-living amoebae in treated drinking water systems present an emerging health risk? Environ. Sci. Technol. 2011, 45, 860–869. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montalbano Di Filippo, M.; Santoro, M.; Lovreglio, P.; Monno, R.; Capolongo, C.; Calia, C.; Fumarola, L.; D'Alfonso, R.; Berrilli, F.; Di Cave, D. Isolation and Molecular Characterization of Free-Living Amoebae from Different Water Sources in Italy. Int. J. Environ. Res. Public Health 2015, 12, 3417-3427. https://doi.org/10.3390/ijerph120403417
Montalbano Di Filippo M, Santoro M, Lovreglio P, Monno R, Capolongo C, Calia C, Fumarola L, D'Alfonso R, Berrilli F, Di Cave D. Isolation and Molecular Characterization of Free-Living Amoebae from Different Water Sources in Italy. International Journal of Environmental Research and Public Health. 2015; 12(4):3417-3427. https://doi.org/10.3390/ijerph120403417
Chicago/Turabian StyleMontalbano Di Filippo, Margherita, Maristella Santoro, Piero Lovreglio, Rosa Monno, Carmen Capolongo, Carla Calia, Luciana Fumarola, Rossella D'Alfonso, Federica Berrilli, and David Di Cave. 2015. "Isolation and Molecular Characterization of Free-Living Amoebae from Different Water Sources in Italy" International Journal of Environmental Research and Public Health 12, no. 4: 3417-3427. https://doi.org/10.3390/ijerph120403417
APA StyleMontalbano Di Filippo, M., Santoro, M., Lovreglio, P., Monno, R., Capolongo, C., Calia, C., Fumarola, L., D'Alfonso, R., Berrilli, F., & Di Cave, D. (2015). Isolation and Molecular Characterization of Free-Living Amoebae from Different Water Sources in Italy. International Journal of Environmental Research and Public Health, 12(4), 3417-3427. https://doi.org/10.3390/ijerph120403417