
Early Exposure to Intermediate-Frequency Magnetic Fields Alters Brain Biomarkers without Histopathological Changes in Adult Mice



Abstract
:1. Introduction
2. Materials and Methods
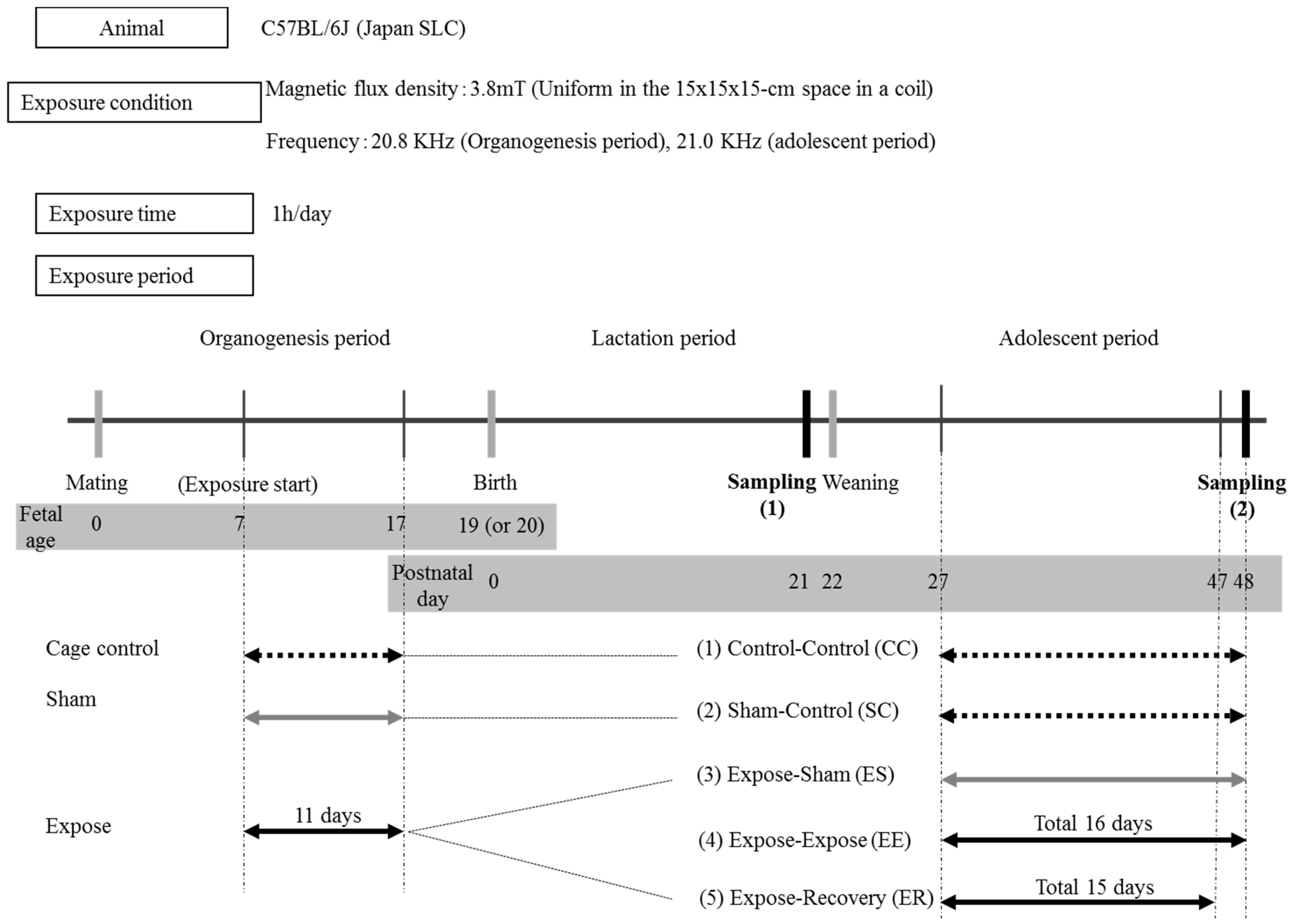
2.1. Animals
2.2. IF-MF Generation
2.3. Exposure to IF-MF

2.4. Quantification of mRNA Expression Levels
2.5. Histological Examination
2.6. Immunohistochemistry
2.7. Statistical Analysis
3. Results
3.1. Body Weight and Organ Weights
3.2. Immediate Early Gene, Memory Function-Related Genes, Their Signal Transduction Pathway Genes and Neurotrophins in the Hippocampus

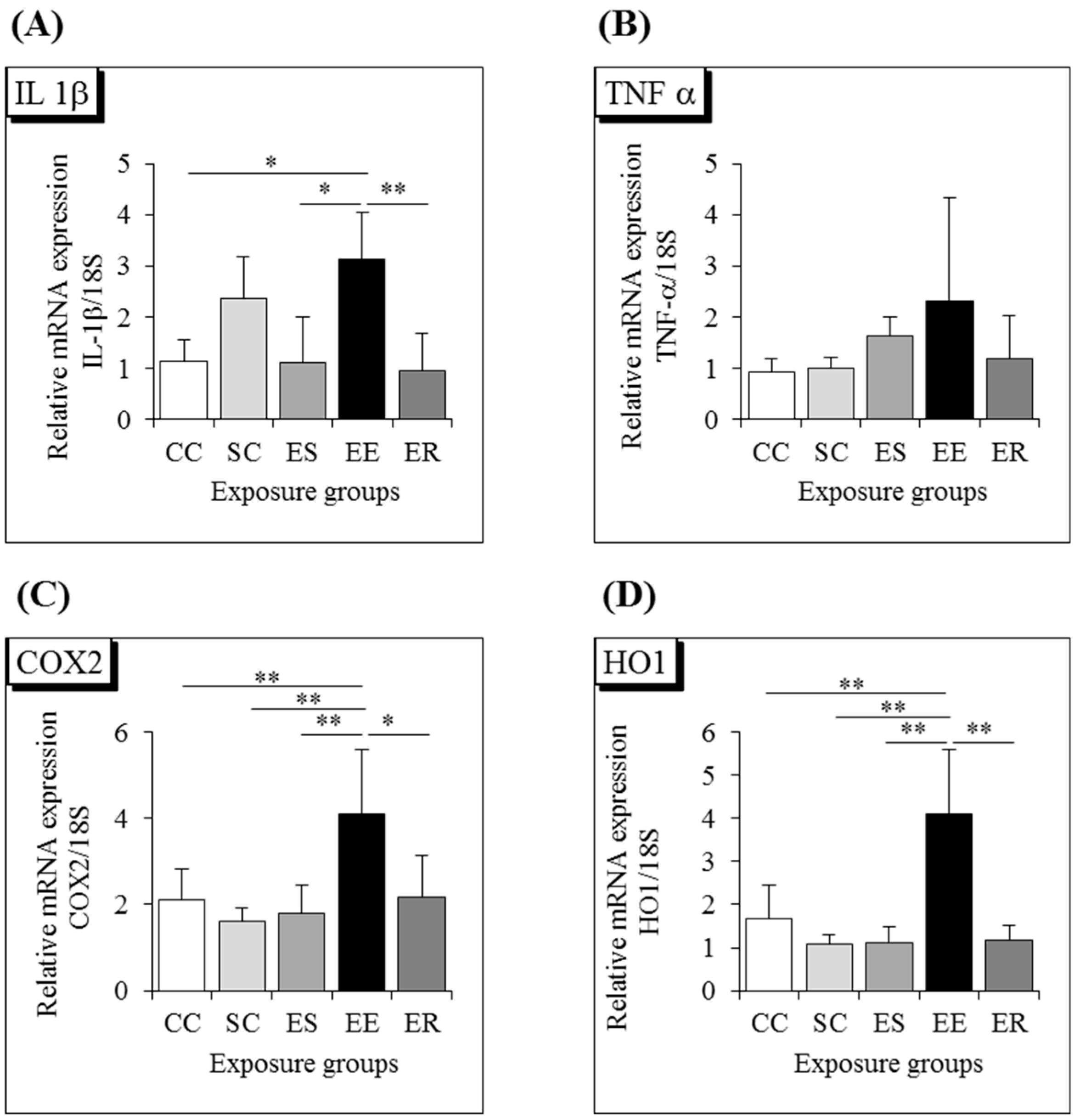
3.3. Proinflammatory Cytokines and Oxidative Stress Markers in the Hippocampus

3.4. Effect of IF-MF Exposure on Histology of Hippocampus


3.5. Effect of IF-MF Exposure on Microglia Activation in the Hippocampus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | In utero Exposure | 7-Week-Old | 7-Week-Old |
|---|---|---|---|
| Biomarker | Organogenesis Exposure | Organogenesis and Adolescent Exposure (EE) | Organogenesis and Adolescent Exposure (ER) |
| c-Fos | − | + | − |
| NR1 | + | + | − |
| NR2A | − | − | − |
| NR2B | + | + | # |
| CaMKIV | − | + | − |
| CREB1 | − | + | # |
| NGF | − | − | − |
| BDNF | − | − | − |
| IL-1β | − | + | # |
| TNF-α | − | + | − |
| COX2 | − | + | # |
| HO1 | − | + | # |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- ICNIRP (International Commission on Non-Ionizing Radiation Protection). ICNIRP statement on EMF-emitting new technologies. Health Phys. 2008, 94, 376–392. [Google Scholar]
- Floderus, B.; Stenlund, C.; Carlgren, F. Occupational exposures to high frequency electromagnetic fields in the intermediate range (4300Hz–10 MHz). Bioelectromagnetics 2002, 23, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Decat, G.; Deckx, L.; Meynen, G.; De Graef, E.; Jonlet, F. European Parliament and of the Council. Magnetic fields of induction heaters in the framework of directive 2004/40/EC of the European Parliament and of the Council. Int. J. Occup. Saf. Ergon. 2006, 12, 169–176. [Google Scholar] [CrossRef] [PubMed]
- WHO (World Health Organization). Extremely Low Frequency Fields. Environmental Health Criteria Monograph No. 238. 2007. Available online: http://www.who.int/peh-emf/publications/elf_ehc/en/index.html (accessed on 25 September 2014).
- Davanipour, Z.; Tseng, C.C.; Lee, P.J.; Sobel, E. A case-control study of occupational magnetic field exposure and Alzheimer’s disease: Results from the California Alzheimer’s Disease Diagnosis and Treatment Centers. BMC Neurol. 2007, 7. [Google Scholar] [CrossRef]
- Davanipour, Z.; Sobel, E. Long-term exposure to magnetic fields and the risks of Alzheimer’s disease and breast cancer: Further biological research. Pathophysiology 2009, 16, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Håkansson, N.; Gustavsson, P.; Sastre, A.; Floderus, B. Occupational exposure to extremely low frequency magnetic fields and mortality from cardiovascular disease. Am. J. Epidemiol. 2003, 158, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Loomis, D.P.; Savitz, D.A. Mortality from brain cancer and leukaemia among electrical workers. Br. J. Ind. Med. 1990, 47, 633–638. [Google Scholar] [PubMed]
- Trimmel, M.; Schweiger, E. Effects of an ELF (50 Hz, 1 mT) electromagnetic field (EMF) on concentration in visual attention, perception and memory including effects of EMF sensitivity. Toxicol. Lett. 1998, 96–97, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Che, Y.; Sun, H.; Cui, Y.; Zhou, D.; Ma, Y. Effects of exposure to 50 Hz magnetic field of 1 mT on the performance of detour learning task by chicks. Brain Res. Bull. 2007, 74, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Che, Y.; Liu, X.; Zhou, D.; Miao, Y.; Ma, Y. Effects of prenatal exposure to a 50-Hz magnetic field on one-trial passive avoidance learning in 1-day-old chicks. Electromagnetics 2010, 31, 150–155. [Google Scholar]
- Nishimura, I.; Imai, S.; Negishi, T. Lack of chick embryotoxicity after 20 kHz, 1.1 mT magnetic field exposure. Bioelectromagnetics 2009, 30, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, I.; Oshima, A.; Shibuya, K.; Negishi, T. Lack of teratological effects in rats exposed to 20 or 60 kHz magnetic fields. Birth Defects Res. B. Dev. Reprod. Toxicol. 2011, 92, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Ushiyama, A.; Ohtani, S.; Suzuki, Y.; Wada, K.; Kunugita, N.; Ohkubo, C. Effects of 21-kHz Intermediate frequency magnetic Fields on blood properties and immune systems of juvenile rats. Int. J. Radiat. Biol. 2014, 90, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Ramundo-Orlando, A.; Mattia, F.; Palombo, A.; D’Inzeo, G. Effect of low frequency, low amplitude magnetic fields on the permeability of cationic liposomes entrapping carbonic anhydrase: II. No evidence for surface enzyme involvement. Bioelectromagnetics. 2000, 21, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Piacentini, R.; Ripoli, C.; Mezzogori, D.; Azzena, G.B.; Grassi, C. Extremely low-frequency electromagnetic fields promote in vitro neurogenesis via upregulation of Ca(v)1-channel activity. J. Cell. Physiol. 2008, 215, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Marchionni, I.; Paffi, A.; Pellegrino, M.; Liberti, M.; Apollonio, F.; Abeti, R.; Fontana, F.; D’Inzeo, G.; Mazzanti, M. Comparison between low-level 50 Hz and 900 MHz electromagnetic stimulation on single channel ionic currents and on firing frequency in dorsal root ganglion isolated neurons. Biochim. Biophys. Acta 2006, 1758, 597–605. [Google Scholar] [CrossRef] [PubMed]
- He, Y.L.; Liu, D.D.; Fang, Y.J.; Zhan, X.Q.; Yao, J.J.; Mei, Y.A. Exposure to extremely low-frequency electromagnetic fields modulates Na+ currents in rat cerebellar granule cells through increase of AA/PGE2 and EP receptor-mediated cAMP/PKA pathway. PLoS ONE. 2013, 8, e54376. [Google Scholar] [CrossRef] [PubMed]
- Miyakoshi, J.; Horiuchi, E.; Nakahara, T.; Sakurai, T. Magnetic fields generated by an induction heating (IH) cook top do not cause genotoxicity in vitro. Bioelectromagnetics 2007, 28, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Win-Shwe, T.T.; Ohtani, S.; Ushiyama, A.; Fujimaki, H.; Kunugita, N. Can intermediate-frequency magnetic fields affect memory function-related gene expressions in hippocampus of C57BL/6J mice? J. Toxicol. Sci. 2013, 38, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Ikehata, M.; Wada, K.; Suzuki, Y.; Yoshie, S.; Sakai, T.; Wake, K.; Nakasono, S.; Taki, M.; Ohkubo, C. Development of exposure system of intermediate frequency magnetic fields for in vitro test systems. In Proceedings of the 32nd Annual Meeting of the Bioelectromagnetics Society, Seoul, Korea, 14–18 June 2010; pp. 429–430.
- Ohtani, S.; Ushiyama, A.; Unno, A.; Hirai, Y.; Suzuki, Y.; Wada, K.; Kunugita, N.; Ohkubo, C. Development of Novel in vivo Exposure Apparatus for Intermediate Frequency Magnetic Field. In Proceedings of the 10th International Congress of the European Bioelectromagnetics Association (EBEA), Rome, Italy, 21–24 February 2011.
- Wada, K.; Kogure, S.; Suzuki, Y. Optimal design of a magnetic-field generator at 20 kHz for biological research. In Proceedings of the 32nd Annual Meeting of the Bioelectromagnetics Society, Seoul, Korea, 14–18 June 2010; pp. 455–456.
- Win-Shwe, T.T.; Yamamoto, S.; Fujitani, Y.; Hirano, S.; Fujimaki, H. Spatial learning and memory function-related gene expression in the hippocampus of mouse exposed to nanoparticle-rich diesel exhaust. Neurotoxicology 2008, 29, 940–947. [Google Scholar] [CrossRef] [PubMed]
- Van Elzakke, M.; Fevurly, R.D.; Breindel, T.; Spencer, R.L. Environmental novelty is associated with a selective increase in Fos expression in the output elements of the hippocampal formation and the perirhinal cortex. Learn. Mem. 2008, 15, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Dragunow, M.; Faull, R. The use of c-fos as a metabolic marker in neuronal pathway tracing. J. Neurosci. Methods 1989, 29, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Day, H.E.; Kryskow, E.M.; Nyhuis, T.J.; Herlihy, L.; Campeau, S. Conditioned Fear Inhibits c-fos mRNA Expression in the Central Extended Amygdala. Brain Res. 2008, 1229, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Dawson, B.V.; Robertson, I.G.; Wilson, W.R.; Zwi, L.J.; Boys, J.T.; Green, A.W. Evaluation of potential health effects of 10 kHz magnetic fields: A rodent reproductive study. Bioelectromagnetics 1998, 19, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, I.; Oshima, A.; Shibuya, K.; Mitani, T.; Negishi, T. Absence of reproductive and developmental toxicity in rats following exposure to a 20-kHz or 60-kHz magnetic field. Regul. Toxicol. Pharmacol. 2012, 64, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Bannerman, D.M.; Good, M.A.; Butcher, S.P.; Ramsay, M.; Morris, R.G. Distinct components of spatial learning revealed by prior training and NMDA receptor blockade. Nature 1995, 378, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Tsien, J.Z.; Huerta, P.T.; Tonegawa, S. The essential role of hippocampal CA1 NMDA receptor-dependent synaptic plasticity in spatial memory. Cell 1996, 87, 1327–1338. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, Z.; Wieraszko, A. The mechanism of magnetic field-induced increase of excitability in hippocampal neurons. Brain Res. 2008, 1221, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, S.; Persinger, M.A. Perinatal exposures to rotating magnetic fields ‘demasculinize’ neuronal density in the medial preoptic nucleus of male rats. Neurosci. Lett. 1998, 253, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Whissell, P.D.; Persinger, M.A. Developmental effects of perinatal exposure to extremely weak 7 Hz magnetic fields and nitric oxide modulation in the Wistar albino rat. Int. J. Dev. Neurosci. 2007, 25, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Zhang, B.; Conti, P. Decreased mitochondrial function and increased brain inflammation in bipolar disorder and other neuropsychiatric diseases. J. Clin. Psychopharmacol. 2011, 31, 685–687. [Google Scholar] [CrossRef] [PubMed]
- Hagberg, H.; Gressens, P.; Mallard, C. Inflammation during fetal and neonatal life: Implications for neurologic and neuropsychiatric disease in children and adults. Ann. Neurol. 2012, 71, 444–457. [Google Scholar] [CrossRef] [PubMed]
- Herbert, M.R. Contributions of the environment and environmentally vulnerable physiology to autism spectrum disorders. Curr. Opin. Neurol. 2010, 23, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Goines, P.E.; Ashwood, P. Cytokine dysregulation in autism spectrum disorders (ASD): Possible role of the environment. Neurotoxicol. Teratol. 2013, 36, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Dimberg, Y. Neurochemical effects of a 20 kHz magnetic field on the central nervous system in prenatally exposed mice. Bioelectromagnetics 1995, 16, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Kavaliers, M.; Innes, D.G. Sex differences in magnetic field inhibition of morphine-induced responses of wild deer mice, Peromyscus maniculatus triangularis. Physiol. Behav. 1987, 40, 559–562. [Google Scholar] [CrossRef] [PubMed]
- Choleris, E.; Thomas, A.W.; Ossenkopp, K.; Kavaliers, M.; Valsecchi, P.; Prato, F.S. Sex differences in conditioned taste aversion and in the effects of exposure to a specific pulsed magnetic field in deer mice Peromyscus maniculatus. Physiol. Behav. 2000, 71, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Bawin, S.M.; Adey, W.R.; Sabbot, I.M. Ionic factors in release of 45Ca2+ from chicken cerebral tissue by electromagnetic fields. Proc. Natl. Acad. Sci. USA 1978, 75, 6314–6318. [Google Scholar] [CrossRef] [PubMed]
- Blackman, C.F.; Benane, S.G.; Rabinowitz, J.R.; House, D.E.; Joines, W.T. A role for the magnetic field in the radiation-induced efflux of calcium ions from brain tissue in vitro. Bioelectromagnetics 1985, 6, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Kavaliers, M.; Ossenkopp, K.P. Magnetic fields differentially inhibit mu, delta, kappa and sigma opiate-induced analgesia in mice. Peptides 1986, 7, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Ge, Z.; Rizak, J.D.; Zhai, C.; Zhou, Z.; Gong, S.; Che, Y. Deficits in water maze performance and oxidative stress in the hippocampus and striatum induced by extremely low frequency magnetic field exposure. PLoS One 2012, 7, e32196. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Win-Shwe, T.-T.; Ohtani, S.; Ushiyama, A.; Kunugita, N. Early Exposure to Intermediate-Frequency Magnetic Fields Alters Brain Biomarkers without Histopathological Changes in Adult Mice. Int. J. Environ. Res. Public Health 2015, 12, 4406-4421. https://doi.org/10.3390/ijerph120404406
Win-Shwe T-T, Ohtani S, Ushiyama A, Kunugita N. Early Exposure to Intermediate-Frequency Magnetic Fields Alters Brain Biomarkers without Histopathological Changes in Adult Mice. International Journal of Environmental Research and Public Health. 2015; 12(4):4406-4421. https://doi.org/10.3390/ijerph120404406
Chicago/Turabian StyleWin-Shwe, Tin-Tin, Shin Ohtani, Akira Ushiyama, and Naoki Kunugita. 2015. "Early Exposure to Intermediate-Frequency Magnetic Fields Alters Brain Biomarkers without Histopathological Changes in Adult Mice" International Journal of Environmental Research and Public Health 12, no. 4: 4406-4421. https://doi.org/10.3390/ijerph120404406
APA StyleWin-Shwe, T. -T., Ohtani, S., Ushiyama, A., & Kunugita, N. (2015). Early Exposure to Intermediate-Frequency Magnetic Fields Alters Brain Biomarkers without Histopathological Changes in Adult Mice. International Journal of Environmental Research and Public Health, 12(4), 4406-4421. https://doi.org/10.3390/ijerph120404406