
Molecular Detection of a Potentially Toxic Diatom Species


Abstract
:1. Introduction
2. Materials and Methods
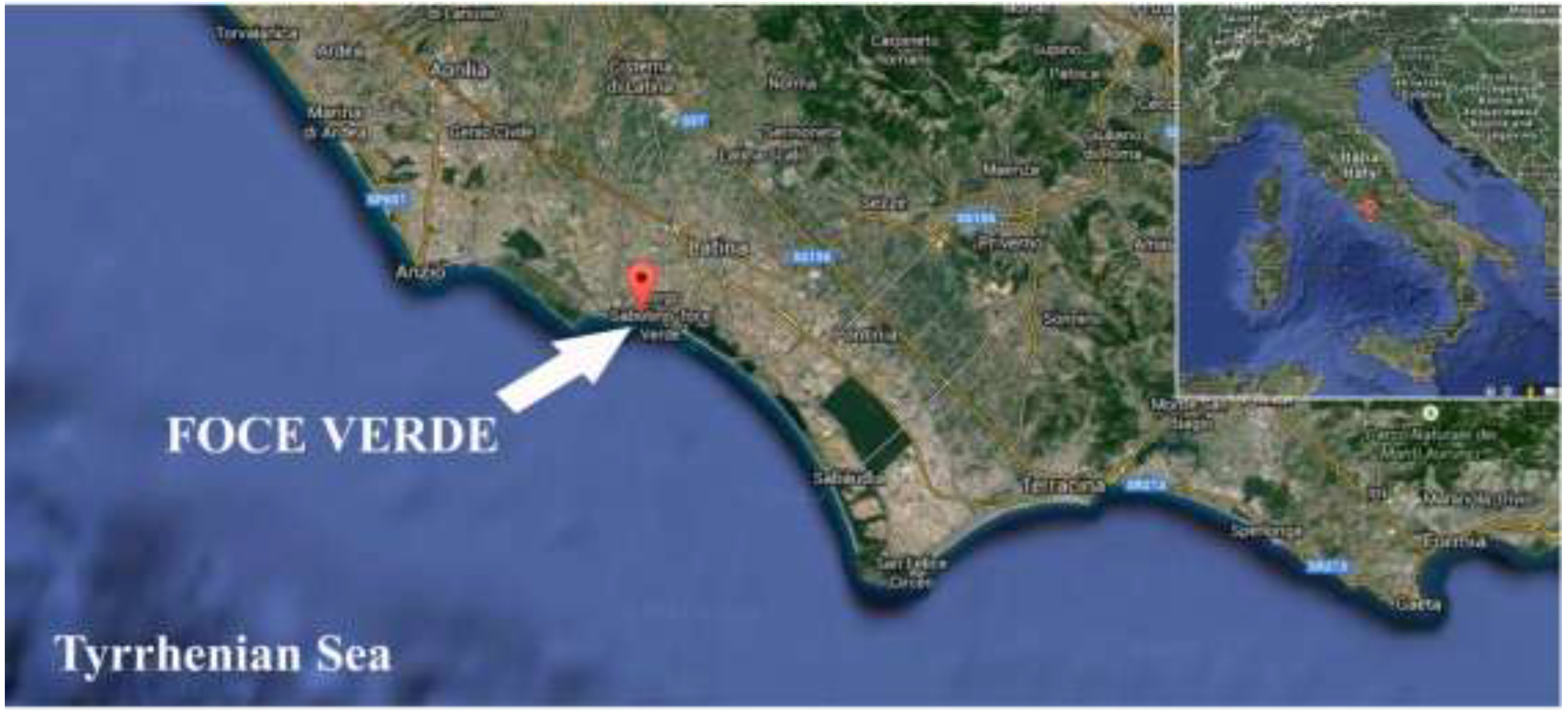
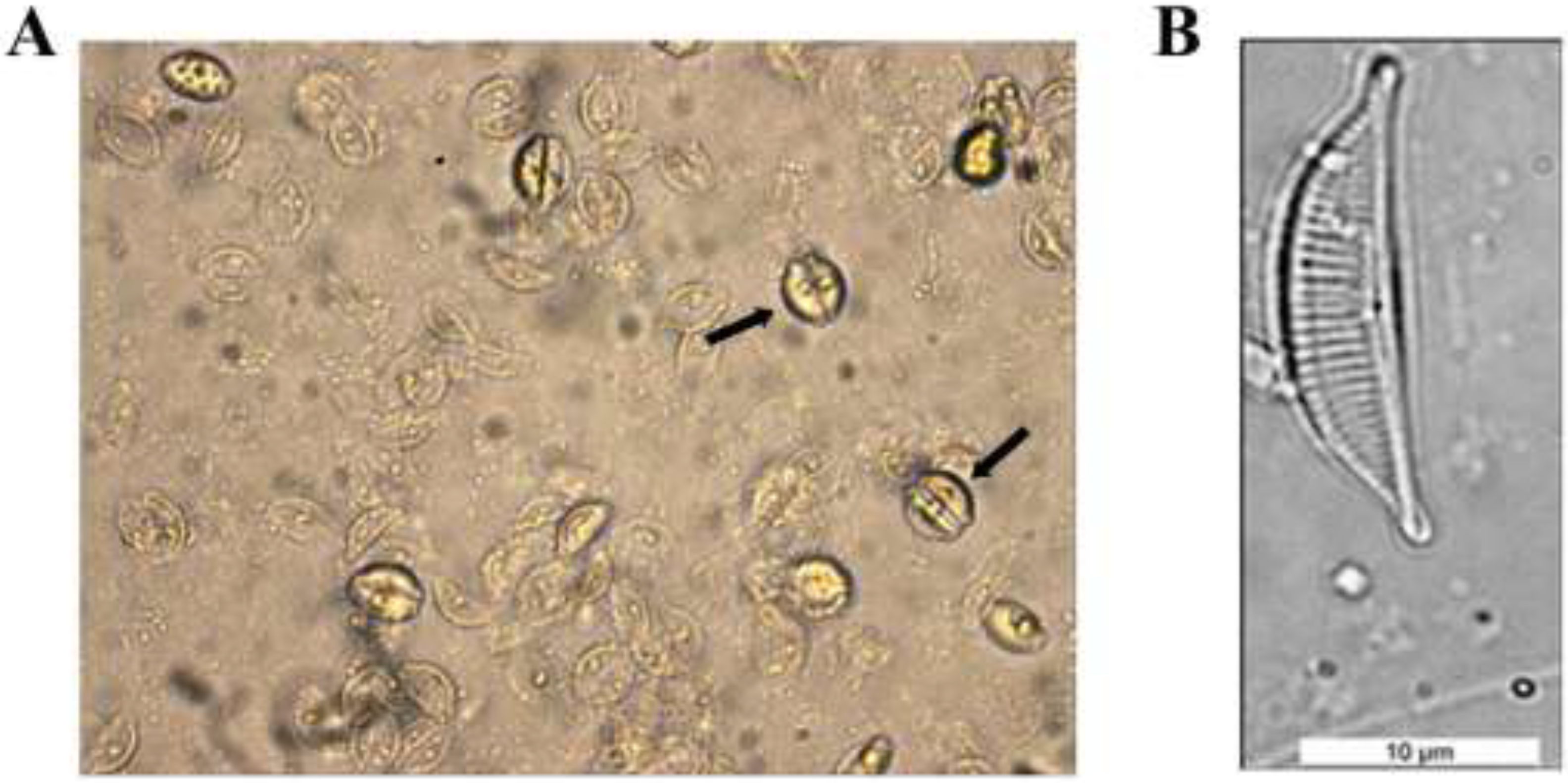
2.1. Diatom Cell Cultures and Identification

2.2. Preparation of Genomic DNA
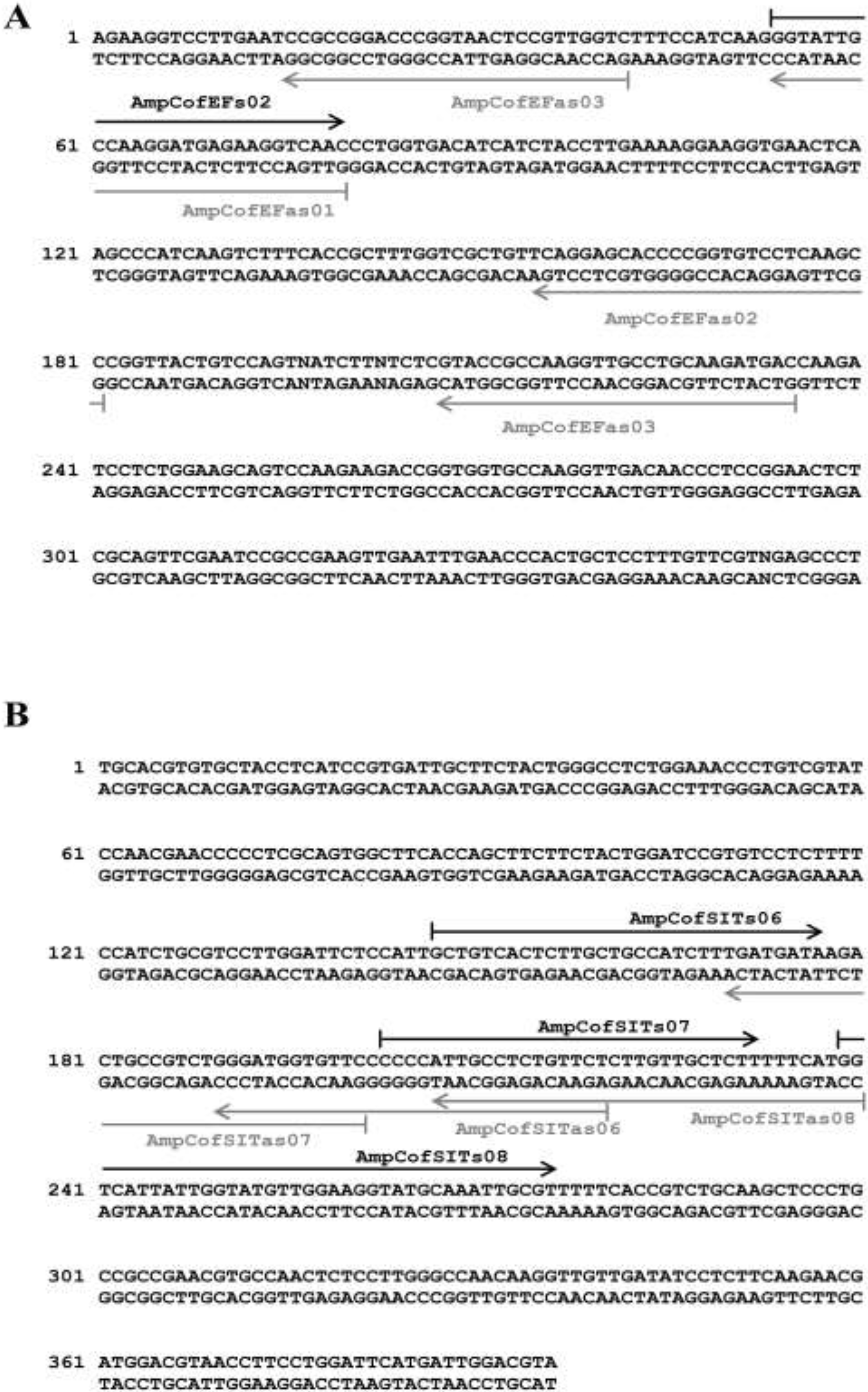
2.3. Primer Design and Oligonucleotide Probe Design
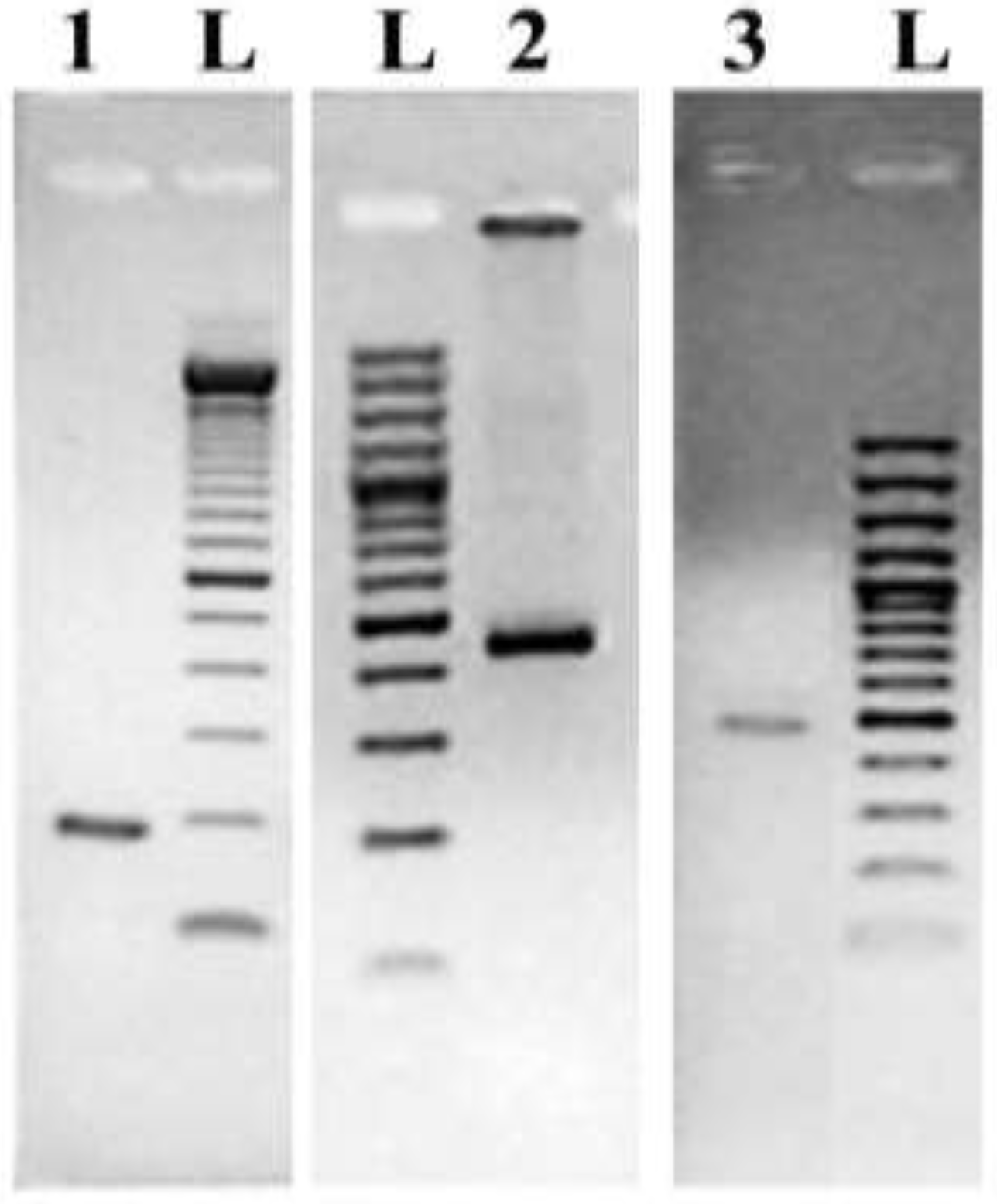
2.4. DNA Amplification of Marker Genes from Genomic DNA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer | Source | Nucleotide Sequence (5’-3’) | Amplicon Size (bp) |
|---|---|---|---|---|
| SSU rRNA | 528F | [42] | GCGGTAATTCCAGCTCCAA | 190 |
| 650R | [42] | AACACTCTAATTTTTTCACAG | ||
| Elongation Factor gene (eEF1-a) | UniE-F | This work | ATCGAACAACACAAGAAGGT | 442 |
| UniE-R | This work | ACCTTTCCAAGCATCTTCAA | ||
| Silicic acid Transporter gene (SIT) | UniS-F1 | This work | GACTTCATCAACAACTACTTCG | 474 |
| UniS-R1 | This work | ACGTCCAATCATGAATCCAG |
| Target | Primers | Initial Denaturation | Denaturation | Annealing | Extension | Final extension | N° of Cycles | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T (°C) | Time (min) | T (°C) | Time (sec) | T (°C) | Time (Sec) | T(°C) | Time (sec) | T (°C) | Time (min) | |||
| eEF1-a | UniE-F/UniE-R | 95 | 2.5 | 94 | 30 | 58 | 60 | 72 | 30 | 72 | 2 | 24–30 |
| SIT | UniS-F1/UniS-R1 | 3 | 30 | 50–58 | 40–50 | 40 | 2 | 30 | ||||
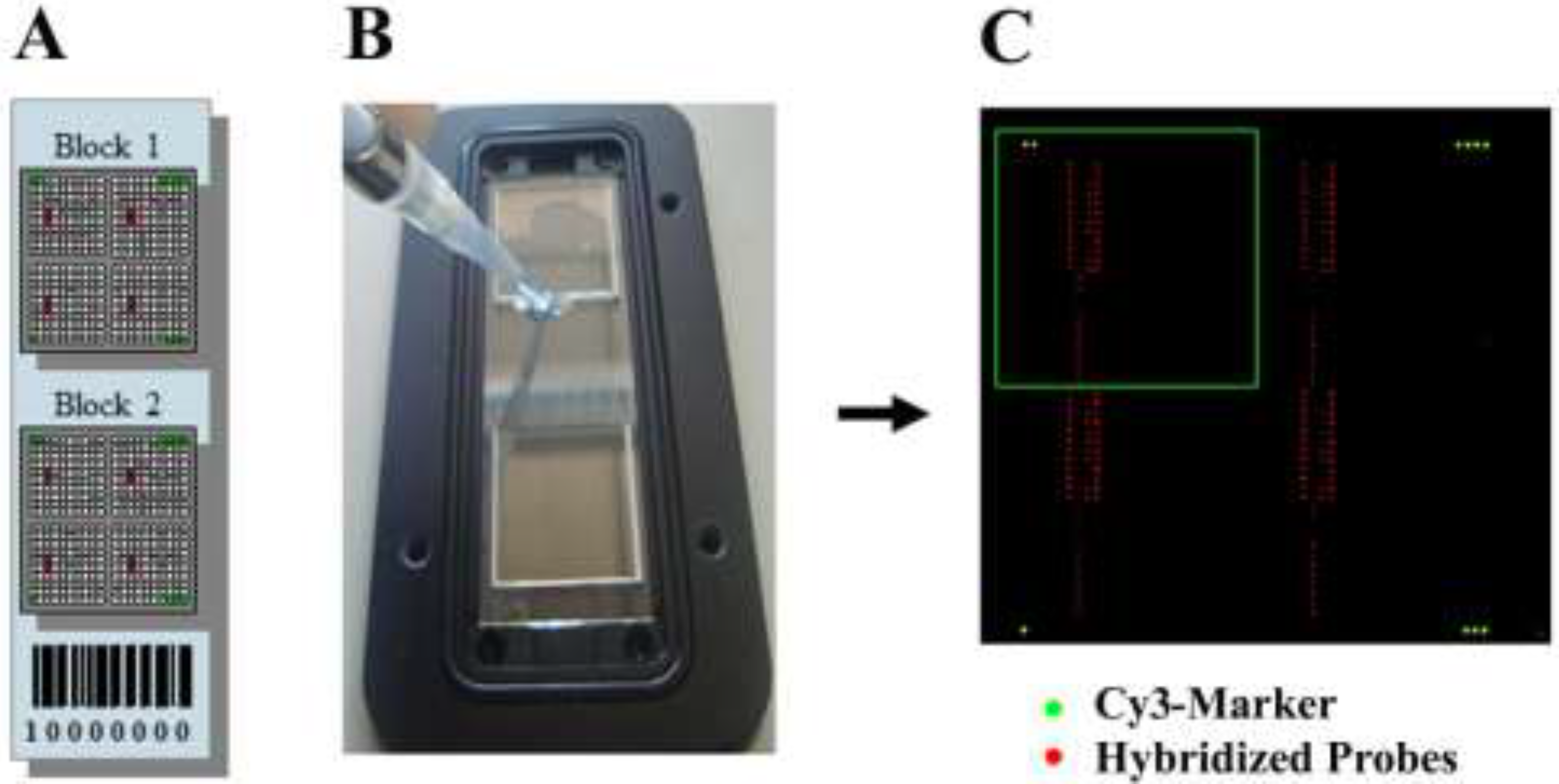
2.5. Set-Up of the Microarray Format

2.6. Microarray Experiments
3. Results
3.1. Isolation and Identification of Diatom Cells from Environmental Samples

3.2. Design of Universal Primers Targeting Marker Genes
3.3. Design of Species-Specific Oligonucleotide Probes

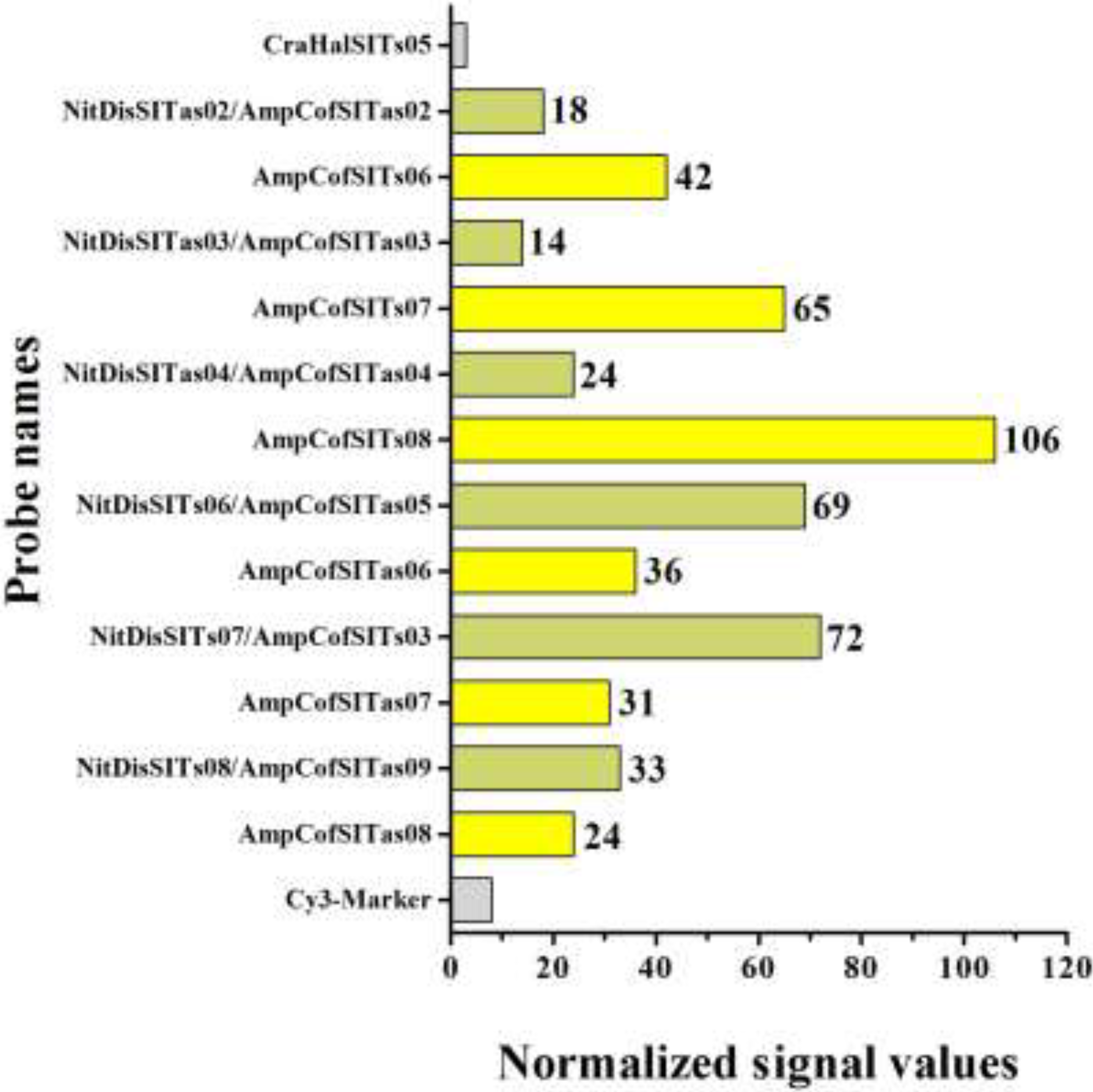
3.4. Use of Microarrays for the Detection of Amphora Coffeaeformis


4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of thebiosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Müller-Navarra, D.C.; Brett, M.T.; Liston, A.M.; Goldman, C.R. A highly unsaturated fatty acid predicts carbon transfer between primary producers and consumers. Nature 2000, 403, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.L.C.; Boyd, R.K.; de Freitas, A.S.W.; Falk, M.; Foxall, R.A.; Jamieson, W.D.; Laycock, M.V.; McCulloch, A.W.; McInnes, A.G.; Odense, P.; et al. Identification of domoic acid, a neuroexcitatory amino acid, in toxic mussels from eastern Prince Edward Island. Can. J. Chem. 1989, 67, 481–490. [Google Scholar] [CrossRef]
- Bates, S.S.; Bird, C.J.; de Freitas, A.S.W.; Foxall, R.; Gilgan, M.; Hanic, L.A.; Johnson, G.R.; McCulloch, A.W.; Odense, P.; Pocklington, R.; et al. Pennate diatom Nitzschia pungens as the primary source of domoic acid, a toxin in shellfish from eastern Prince Edward Island, Canada. Can. J. Fish. Aquat. Sci. 1989, 46, 1203–1215. [Google Scholar] [CrossRef]
- Bates, S.S. Domoic acid producing diatoms: Another genus added. J. Phycol. 2000, 36, 978–983. [Google Scholar] [CrossRef]
- Landsberg, J.H. The effects of harmful algal blooms on aquatic organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar] [CrossRef]
- Trainer, V.L.; Hickey, B.M.; Bates, S.S. Toxic diatoms. In Oceans and Human Health: Risks and Remedies from the Sea; Walsh, P.J., Smith, S.L., Fleming, L.E., Solo-Gabriele, H., Gerwick, W.H., Eds.; Elsevier Science Publishers: New York, NY, USA, 2008; pp. 219–238. [Google Scholar]
- Fernandes, L.F.; Hubbard, K.A.; Richlen, M.L.; Smith, J.; Bates, S.S.; Ehrman, J.; Léger, C.; Mafra, L.L., Jr.; Kulis, D.; Quilliam, M.; et al. Diversity and toxicity of the diatom Pseudo-nitzschia Peragallo in the Gulf of Maine, Northwestern Atlantic Ocean. Deep-Sea Res. Pt. II 2014, 103, 139–162. [Google Scholar]
- Vale, P.; Sampayo, M.A.M. Domoic acid in Portuguese shellfish and fish. Toxicon 2001, 39, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Penna, A.; Bertozzini, E.; Battocchi, C.; Galluzzi, L.; Giacobbe, M.G.; Vila, M.; Garces, E.; Luglie, A.; Magnani, M. Monitoring of HAB species in the Mediterranean Sea through molecular methods. J. Plankton Res. 2007, 29, 19–38. [Google Scholar] [CrossRef]
- Amzil, Z.; Fresnel, J.; Le Gal, D.; Billard, C. Domoic acid accumulation in French shellfish in relation to toxic species of Pseudo-nitzschia multiseries and P. pseudodelicatissima. Toxicon 2001, 39, 1245–1251. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Magano, G.S.; Tartaglione, L.; Quilliam, M.A.; Tubaro, A.; Polleti, R. Hydrophilic interaction liquid chromatography/mass spectrometry for determination of domoic acid in Adriatic shellfish. Rapid. Commun. Mass Spectrom. 2005, 19, 2030–2038. [Google Scholar] [CrossRef] [PubMed]
- Facca, C.; Bilaničovà, D.; Pojana, G.; Sfriso, A.; Marcomini, A. Harmful algae records in Venice lagoon and in Po River Delta (Northern Adriatic Sea, Italy). Sci. World J. 2014, 1–11. [Google Scholar]
- Ujević, I.; Ninčević-Gladan, Z.; Roje, R.; Skejić, S.; Arapov, J.; Marasović, I. Domoic acid—A new toxin in the Croatian Adriatic shellfish toxin profile. Molecules 2010, 15, 6835–6849. [Google Scholar] [CrossRef] [PubMed]
- Kaniou-Grigoriadou, I.; Mouratidou, T.; Katikou, P. Investigation on the presence of the domoic acid in Greek shellfish. Harmful Algae 2005, 4, 717–723. [Google Scholar] [CrossRef]
- James, K.J.; Gillman, M.; Amandi, M.F.; López-Rivera, A.; Puente, P.F.; Lehane, M.; Mitrović, S.; Furey, A. Amnesic shellfish poisoning toxin in bivalve molluscs in Ireland. Toxicon 2005, 46, 852–858. [Google Scholar] [CrossRef] [PubMed]
- Ajani, P.; Murray, S.; Hallegraeff, G.; Brett, S.; Armand, L. First reports of Pseudo-nitzschia micropora and P. hasleana (Bacillariaceae) from the Southern Hemisphere: Morphological, molecular and toxicological characterization. Phycol. Res. 2013, 61, 237–248. [Google Scholar] [CrossRef]
- Ajani, P.; Murray, S.; Hallegraeff, G.; Lundholm, N.; Gillings, M.; Brett, S.; Armand, L. The diatom genus Pseudo-nitzschia (Bacillariophyceae) in new south wales, Australia: Morphotaxonomy, molecular phylogeny, toxicity, and distribution. J. Phycol. 2013, 49, 765–785. [Google Scholar] [CrossRef]
- Shimizu, Y.; Gupta, S.; Masuda, K.; Maranda, L.; Walker, C.K.; Wang, R. Dinoflagellate and other microalgal toxins: Chemistry and biochemistry. Pure. Appl. Chem. 1989, 61, 513–516. [Google Scholar] [CrossRef]
- Maranda, L.; Wang, R.; Masuda, K.; Shimizu, Y. Investigation of the source of domoic acid in mussels. In Toxic Marine Phytoplankton; Granéli, E., Sundström, B., Edler, L., Anderson, D.M., Eds.; Elsevier Science Publishing Co.: New York, NY, USA, 1990; pp. 300–304. [Google Scholar]
- Sala, S.E.; Sar, E.A.; Ferrario, M.E. Review of materials reported as containing Amphora coffeaeformis (Agardh) Kützing in Argentina. Diatom Res. 1998, 13, 323–336. [Google Scholar] [CrossRef]
- Lee, K.; Round, F.E. Studies on freshwater Amphora species. In Amphora ovalis. Diatom Res. 1987, 2, 193–203. [Google Scholar] [CrossRef]
- Sala, S.E.; Guerrero, J.M.; Coste, M. Valve morphology of Amphora chilensis Hustedt (Bacillariophyceae). Nova Hedwigia 2007, 85, 353–364. [Google Scholar] [CrossRef]
- Fourtanier, E.; Kociolek, J.P. Catalogue of Diatom Names. Part II: Abas. through Bruniopsis. Occas. pap. Calif. Acad. Sci. 2009, 156. [Google Scholar]
- Levkov, Z. Amphora sensu lato. In Diatoms of Europe: Diatoms of the European Inland Waters and Comparable Habitats; Lange-Bertalot, H., Ed.; A.R.G. Gantner Verlag K.G.: Ruggell, Liechtenstein, 2009. [Google Scholar]
- Park, J.; Koh, C.-H. Taxonomic studies on korean marine benthic diatoms—LM and SEM observations of the diatom genus Amphora (bacillariophyceae) from Korean tidal flats with the first recordings of A. arenicola, A. beaufortiana and A. maletractata var. constricta. Ocean. Sci. J. 2012, 47, 101–112. [Google Scholar]
- Stepanek, J.G.; Kociolek, J.P. Several new species of Amphora and Halamphora from the western USA. Diatom Res. 2013, 28, 61–76. [Google Scholar] [CrossRef]
- Wang, P.; Park, B.S.; Kim, J.H.; Kim, J.-H.; Lee, H.-O.; Han, M.-S. Phylogenetic position of eight Amphora sensu lato (Bacillariophyceae) species and comparative analysis of morphological characteristics. Algae 2014, 29, 57–73. [Google Scholar] [CrossRef]
- Scholin, C.A.; Marin, R.; Miller, P.E.; Doucette, G.J.; Powell, C.L.; Haydock, P.; Judith Howard, J.; Ray, J. DNA probes and a receptor-binding assay for detection of Pseudo-nitzschia (Bacillariophyceae) species and domoic acid activity in cultured and natural samples. Phycol. 1999, 35, 1356–1367. [Google Scholar] [CrossRef]
- Penna, A.; Galluzzi, L. The quantitative real-time PCR applications in the monitoring of marine harmful algal bloom (HAB) species. Environ. Sci. Pollut. Res. 2013, 20, 6851–6862. [Google Scholar] [CrossRef]
- Lelong, A.; Hégaret, H.; Soudant, P.; Bates, S.S. Pseudo-nitzschia (Bacillariophyceae) species, domoic acid and amnesic shellfish poisoning: revisiting previous paradigms. Phycologia 2012, 51, 168–216. [Google Scholar] [CrossRef]
- Trainer, V.L.; Bates, S.S.; Lundholm, N.; Thessen, A.E.; Cochlan, W.P.; Adams, N.G.; Trick, C.G. Pseudo-nitzschia physiological ecology, phylogeny, toxicity, monitoring and impacts on ecosystem health. Harmful Algae 2012, 14, 271–300. [Google Scholar] [CrossRef]
- Lim, H.C.; Teng, S.T.; Leaw, C.P.; Lim, P.T. Three novel species in the Pseudo-nitzschia pseudodelicatissima complex: P. batesiana sp. nov., P. lundholmiae sp. nov. and P. fukuyoi sp. nov. (Bacillariophyceae), from the Strait of Malacca, Malaysia. J. Phycol. 2013, 49, 902–916. [Google Scholar]
- Orive, E.; Perez-Aicua, L.; David, H. The genus Pseudo-nitzschia (Bacillariophyceae) in a temperate estuary with description of two new species: Pseudo-nitzschia plurisecta sp. nov. and Pseudo-nitzschia abrensis sp. nov. J. Phycol. 2013, 49, 1192–1206. [Google Scholar]
- LIFEHAB: Life histories of microalgal species causing harmful blooms. Report of a European workshop organised jointly by the Fifth Framework Programme Energy, Environment and Sustainable Development of the European Commission, the Institut de Ciències del Mar, CMIMA-CSIC (Barcelona) and the Calvià Town Council (Majorca, Balearic Islands); Garcés, E.; Zingone, A.; Montresor, M.; Reguera, B.; Dale, B. (Eds.) Calvià, Majorca, Spain, 2001.
- Lefebvre, K.A.; Robertson, A. Domoic acid and human exposure risks: A review. Toxicon 2010, 56, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Morel, F.M.M.; Rueter, J.G.; Anderson, D.M.; Guillard, R.R.L. Aquil: A chemically defined phytoplankton culture medium for trace metal studies. J. Phycol. 1979, 15, 135–141. [Google Scholar] [CrossRef]
- Price, N.M.; Harrison, G.I.; Hering, J.G.; Hudson, R.J.; Nirel, P.M.V.; Palenik, B.; Morel, F.M.M. Preparation and chemistry of the artificial algal culture medium Aquil. Biological Oceanog. 1989, 6, 443–461. [Google Scholar]
- L’indice Diatomico di Eutrofizzazione/Polluzione (EPI-D) nel Monitoraggio delle Acque Correnti. Linee Guida. Available online: http://www.cisba.eu/attachments/article/75/IndiceEPI_D.pdf (assessed on 16 April 2015).
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. In Süβwasserflora von Mitteleuropa; Gustav Fischer-Verlag: Stuttgart, Germany, 1988. [Google Scholar]
- Nguyen, T.N.M.; Berzano, M.; Gualerzi, C.O.; Spurio, R. Development of molecular tools for the detection of freshwater diatoms. J. Microbiol. Meth. 2011, 84, 33–40. [Google Scholar] [CrossRef]
- Dittami, S.M.; Edvardsen, B. GPR-Analyzer: A simple tool for quantitative analysis of hierarchical multispecies microarrays. Environ. Sci. Pollut. Res. Int. 2013, 20, 6808–6815. [Google Scholar] [CrossRef] [PubMed]
- UNI EN. Water quality. In Guidance Standard for the Routine Sampling and Pre-Treatment of Benthic Diatom Samples from Rivers; European committee for standardization: Brussels, Belgium, 2005. [Google Scholar]
- Van Dam, H.; Mertens, A.; Sinkeldam, J. A coded checklist and ecological indicator values of freshwater diatoms from the Netherlands. Nether. J. Aquat. Ecol. 1994, 28, 117–133. [Google Scholar]
- Armbrust, E.V.; Berges, J.A.; Bowler, C.; Green, B.R.; Martinez, D.; Putnam, N.H.; Zhou, S.G.; Allen, A.E.; Apt, K.E.; Bechner, M.; et al. The genome of the diatom Thalassiosira pseudonana: Ecology, evolution, and metabolism. Science 2004, 306, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Bowler, C.; Bowle, C.; Allen, A.E.; Badger, J.H.; Grimwood, J.; Jabbari, K.; Kuo, A.; Maheswari, U.; Martens, C.; Maumus, F.; Otillar, R.P.; et al. The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 2008, 456, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Cimarelli, L.; Singh, K.S.; Mai, N.T.N.; Dhar, B.D.; Brandi, A.; Brandi, L.; Spurio, R. Molecular tools for the selective detection of nine diatom species biomarkers of various water quality levels. Int. J. Environ. Res. Public Health. (under review).
- Bruce, K.L.; Leterme, S.C.; Ellis, A.V.; Lenehan, C.E. Approaches for the detection of harmful algal blooms using oligonucleotide interactions. Anal. Bioanal. Chem. 2015, 407, 95–116. [Google Scholar] [CrossRef] [PubMed]
- Barra, L.; Ruggiero, M.V.; Chen, J.; Kooistra, W.H. Specificity of LSU rRNA-targeted oligonucleotide probes for Pseudo-nitzschia species tested through dot-blot hybridisation. Environ. Sci. Pollut. Res. 2014, 21, 548–557. [Google Scholar] [CrossRef]
- Hosoi-Tanabe, S.; Sako, Y. Species-specific detection and quantification of toxic marine dinoflagellates Alexandrium tamarense and A. catenella by Real-Time PCR assay. Mar. Biotech. 2005, 7, 506–514. [Google Scholar] [CrossRef]
- Landis, F.C.; Gargas, A. Using ITS2 secondary structure to create species-specific oligonucleotide probes for fungi. Mycologia 2007, 99, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Blomster, J.; Hoey, E.M.; Maggs, C.A.; Stanhope, M.J. Species-specific oligonucleotide probes for macroalgae: molecular discrimination of two marine fouling species of Enteromorpha (Ulvophyceae). Molec. Ecol. 2000, 9, 177–186. [Google Scholar] [CrossRef]
- Bates, S.S.; Freitas, A.S.W.; Milley, J.E. Controls on DA production by the diatom Nitzschia pungens f. multiseries in culture: Nutrients and irradiance. Can. J. Fish. Aquat. Sci. 1991, 48, 1136–1144. [Google Scholar] [CrossRef]
- Pan, Y.; Rao, D.V.S.; Mann, K.H.; Brown, R.G.; Pocklington, R. Effects of silicate limitation on production of DA, a neurotoxin, by the diatom Nitzschia pungens f. multiseries (Hasle) I. Batch culture studies. Mar. Ecol.-Progr. Ser. 1996, 131, 225–233. [Google Scholar] [CrossRef]
- Lelong, A.; Hégaret, H.; Soudant, P. Link between domoic acid production and cell physiology after exchange of bacterial communities between toxic Pseudo-nitzschia multiseries and non-toxic Pseudo-nitzschia delicatissima. Mar. Drugs 2014, 12, 3587–3607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Takata, Y.; Kodama, M. Direct contact between Pseudo-nitzschia multiseries and bacteria is necessary for the diatom to produce a high level of domoic acid. Fisher. Sci. 2009, 75, 771–776. [Google Scholar] [CrossRef]
- Smith, M.W.; Maier, M.A.; Suciu, D.; Peterson, T.D.; Bradstreet, T.; Nakayama, J.; Simon, H.M. High resolution microarray assay for rapid taxonomic assessment of Pseudo-nitzschia spp. (Bacillariophyceae) in the field. Harmful Algae 2012, 19, 169–180. [Google Scholar] [CrossRef]
- Barra, L.; Ruggiero, M.V.; Sarno, D.; Montresor, M.; Kooistra, W.H.C.F. Strengths and weaknesses of microarray approaches to detect Pseudo-nitzschia species in the field. Environ. Sci. Pollut. Res. 2013, 20, 6705–6718. [Google Scholar] [CrossRef]
- Sessitsch, A.; Hackl, E.; Wenzl, P.; Kilian, A.; Kostic, T.; Stralis-Pavese, N.; Tankouo Sandjong, B.; Bodrossy, L. Diagnostic microbial microarrays in soil Ecology. New Phytol. 2006, 171, 719–736. [Google Scholar] [CrossRef] [PubMed]
- Kermarrec, L.; Franc, A.; Rimet, F.; Chaumeil, P.; Frigerio, J.M.; Humbert, J.F.; Bouchez, A. A next-generation sequencing approach to river biomonitoring using benthic diatoms. Freshwater Sci. 2014, 33, 349–364. [Google Scholar] [CrossRef]
- Buhmann, M.T.; Poulsen, N.; Klemm, J.; Kennedy, M.R.; Sherrill, C.D.; Kroger, N. A tyrosine-rich cell surface protein in the diatom Amphora coffeaeformis identified through transcriptome analysis and genetic transformation. PLoS One 2014, 9, 1–11. [Google Scholar] [CrossRef]
- Siaut, M.; Heijde, M.; Mangogna, M.; Montsant, A.; Coesel, S.; Allen, A.; Manfredonia, A.; Falciatore, A.; Bowler, C. Molecular toolbox for studying diatom biology in Phaeodactylum tricornutum. Gene 2007, 406, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Evans, K.M.; Wortley, A.H.; Mann, D.G. An assessment of potential diatom “barcode” genes (cox1, rbcL, 18S and ITS rDNA) and their effectiveness in determining relationships in Sellaphora (Bacillariophyta). Protist 2007, 158, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Moniz, M.B.J.; Kaczmarska, I. Barcoding of diatoms: Nuclear encoded ITS revisited. Protist 2010, 161, 7–34. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhar, B.C.; Cimarelli, L.; Singh, K.S.; Brandi, L.; Brandi, A.; Puccinelli, C.; Marcheggiani, S.; Spurio, R. Molecular Detection of a Potentially Toxic Diatom Species. Int. J. Environ. Res. Public Health 2015, 12, 4921-4941. https://doi.org/10.3390/ijerph120504921
Dhar BC, Cimarelli L, Singh KS, Brandi L, Brandi A, Puccinelli C, Marcheggiani S, Spurio R. Molecular Detection of a Potentially Toxic Diatom Species. International Journal of Environmental Research and Public Health. 2015; 12(5):4921-4941. https://doi.org/10.3390/ijerph120504921
Chicago/Turabian StyleDhar, Bidhan Chandra, Lucia Cimarelli, Kumar Saurabh Singh, Letizia Brandi, Anna Brandi, Camilla Puccinelli, Stefania Marcheggiani, and Roberto Spurio. 2015. "Molecular Detection of a Potentially Toxic Diatom Species" International Journal of Environmental Research and Public Health 12, no. 5: 4921-4941. https://doi.org/10.3390/ijerph120504921
APA StyleDhar, B. C., Cimarelli, L., Singh, K. S., Brandi, L., Brandi, A., Puccinelli, C., Marcheggiani, S., & Spurio, R. (2015). Molecular Detection of a Potentially Toxic Diatom Species. International Journal of Environmental Research and Public Health, 12(5), 4921-4941. https://doi.org/10.3390/ijerph120504921