3.1. Mutant p53 R248Q is Elevated in HCC70 Cells Similarly to Other Variants of mtp53
We compared the mtp53 protein expression levels in a number of established human breast cancer cell lines (see
Table 1 for details). The p53 expression panel included a comparison of wt as well as R248Q, R248W, R273H, null, R280K and C194T mutants. Stable mtp53 protein expression was detected by western blot analysis of whole cell extracts from the human breast cancer cell lines MDA-MB-231, T47D, MDA-MB-468, HCC2157, and HCC70 (
Figure 1, lane 1 and 3–6 respectively). In the HCC1806 cell line mtp53 has a 2bp insertion, causing no p53 protein to be detected (
Figure 1, lane 7). In the MCF-7 cell line the p53 expressed is wild-type and therefore was present at a low level that was not detectable at the exposure shown (
Figure 1, lane 2). In a collaborative study we previously showed that R273H and R280K confer gain-of-function characteristics to two TNBCs breast cancer cell lines, MDA-MD-468 and MDA-MB-231 respectively [
13] We were interested to determine if the AA-derived breast cancer cell lines HCC70 with R248Q and HCC1806 with no p53 expression showed increased deformability when compared to the wild-type p53 expression cell line MCF-7.
Table 1.
Panel of breast cancer cell lines. Types of human breast cancer cell lines used for comparison of p53 protein expression levels. TP53 mutation status and cell line characteristics are listed.
Table 1.
Panel of breast cancer cell lines. Types of human breast cancer cell lines used for comparison of p53 protein expression levels. TP53 mutation status and cell line characteristics are listed.
| Cell Line | P53 Status | Charcteristivs |
|---|
| HCC70 | R248Q | AA, TNBC |
| HCC2175 | R248W | AA, TNBC |
| MDA-MB-46 | 273 | AA, TNBC |
| HCC1809 | Null (2bp insertion) | AA, TNBC |
| MD-MB-231 | R280K | Non-AA, TNBC |
| T47D | C194T | Non-AA |
| MCF7 | wt | Non-AA |
Figure 1.
mtp53 is highly expressed in breast cancer cell lines. Expression of p53 protein in a panel of breast cancer cell lines: MDA-MB-231, MCF7, T47D, MDA-MB-468, HCC2157, HCC70 and HCC1806 and (lanes 1–7, respectively). 50 μg of protein extracted from different cell lines using RIPA buffer, were separated by gradient 4%–12% SDS/PAGE and analyzed by western blot using antibody to p53. Actin was used as a loading control. A representative blot is shown.
Figure 1.
mtp53 is highly expressed in breast cancer cell lines. Expression of p53 protein in a panel of breast cancer cell lines: MDA-MB-231, MCF7, T47D, MDA-MB-468, HCC2157, HCC70 and HCC1806 and (lanes 1–7, respectively). 50 μg of protein extracted from different cell lines using RIPA buffer, were separated by gradient 4%–12% SDS/PAGE and analyzed by western blot using antibody to p53. Actin was used as a loading control. A representative blot is shown.
3.3. Knockdown of mtp53 R248Q in HCC70 Cells
In order to further examine the associated actions of the mtp53 R248Q in the HCC70 cells we used our established protocol to genetically engineer mir30-based shRNA knockdown derivatives of HCC70 (from now on called HCC70.shp53) [
21]. We used shRNA mediated knockdown to test if the expression of mtp53 R248Q in the HCC70 cell line was directly responsible for the increased deformability. First we analyzed the efficiency of the knockdown in the HCC70.shp53 pool and next in three clonal cell lines (
Figure 3a,b). The expression of mtp53 was significantly decreased by the addition of doxycycline in the HCC70.shp53 pool, as well as clones 5A1, 5D4 and 5D2 (
Figure 3a, compare lanes 4 to 5, 6 to 7, and 10 to 11). No change in mtp53 level was detected following doxycycline addition to the HCC70.STGM vector control cell line (
Figure 3a, compare lane 2 to 3). The clone 5A1 showed an excellent knockdown of mtp53 R248Q. This knockdown was validated further by immunofluorescence imaging which showed that following doxycycline treatment the HCC.shp53 clone 5A1 had increased expression of GFP and reduced mtp53 staining (
Figure 3b). HCC70 STGM control cells showed no doxycycline-mediated decrease in mtp53 staining (
Figure 3b).
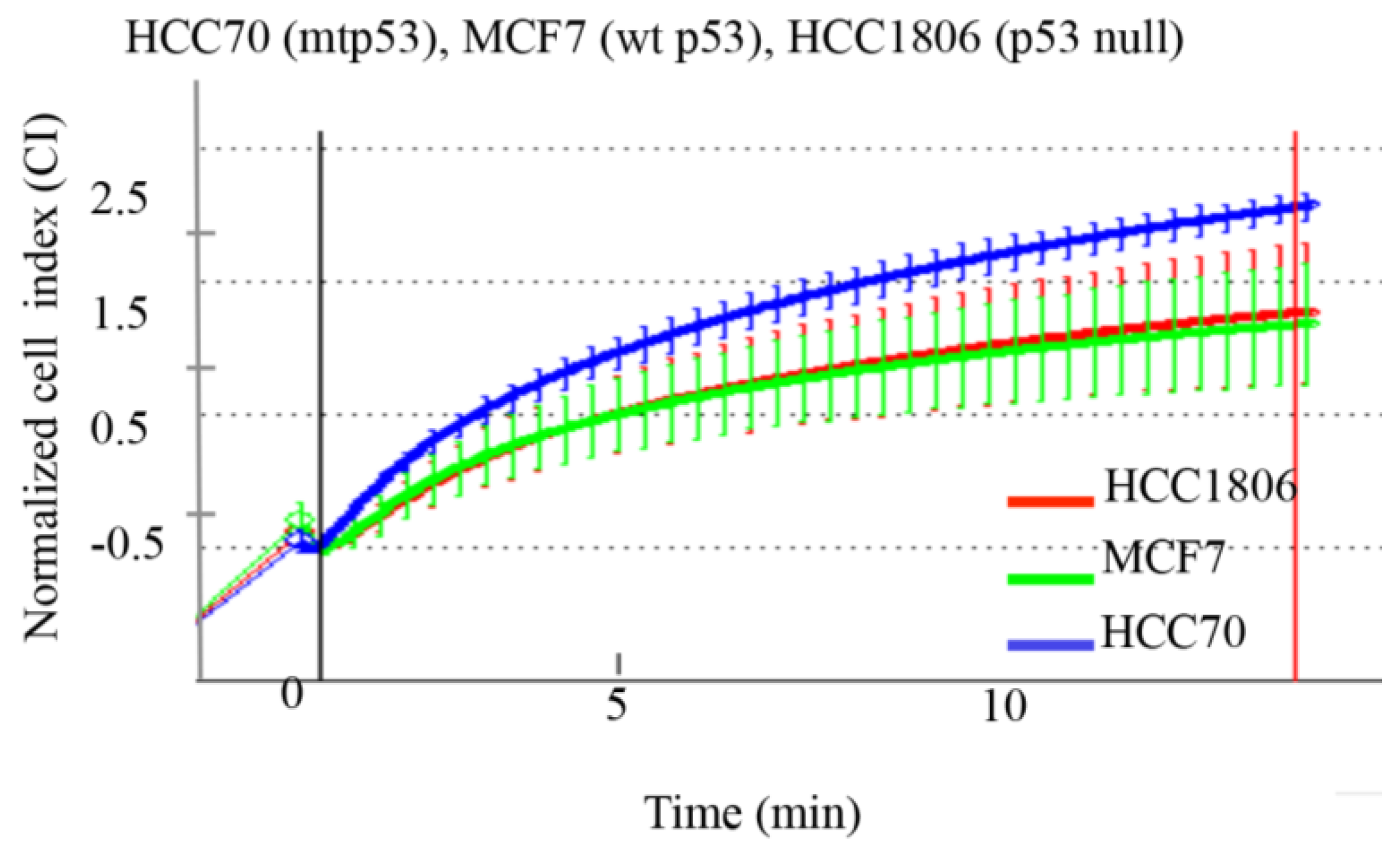
Figure 2.
Expression of mtp53 in the HCC70 cell line is associated with increase cell deformability compared to cell lines with wt or no p53. Average variations of the impedance (CI) as a function of time and resulting swelling are shown. Mtp53-expressing cell lines (HCC70, red), wt p53-expressing cell line (MCF7, green) and no p53-expressing cell line (HCC1806, blue) were measured under hypotonic stress using the xCELLigence system. Cell volume (as Normalized Cell Index) was recorded every 2 s following induction of swelling, as described in materials and methods. The HCC70 cell line (red) showed significantly higher deformability, compared to MCF7 and HCC1806 cell lines (green and blue, respectively). Average values and standard deviation at each time points are shown (n = 4). The first vertical line indicates the addition of water. The second vertical line (shown in red) was used to generate slopes of the curves (data not shown).
Figure 2.
Expression of mtp53 in the HCC70 cell line is associated with increase cell deformability compared to cell lines with wt or no p53. Average variations of the impedance (CI) as a function of time and resulting swelling are shown. Mtp53-expressing cell lines (HCC70, red), wt p53-expressing cell line (MCF7, green) and no p53-expressing cell line (HCC1806, blue) were measured under hypotonic stress using the xCELLigence system. Cell volume (as Normalized Cell Index) was recorded every 2 s following induction of swelling, as described in materials and methods. The HCC70 cell line (red) showed significantly higher deformability, compared to MCF7 and HCC1806 cell lines (green and blue, respectively). Average values and standard deviation at each time points are shown (n = 4). The first vertical line indicates the addition of water. The second vertical line (shown in red) was used to generate slopes of the curves (data not shown).
Figure 3.
Inducible depletion of mtp53 proteins R248Q in breast cancer cell line HCC70. (a) The mtp53 expression in whole-cell extracts from parental HCC70 cell line (lane 1) and cell lines expressing vector control (control, lane 2,3) or mtp53-targeted shRNA (lanes 4–11), including HCC70. shp53 italic or not—Must be consistent pool (lane 4,5) and clonal cell lines (HCC70.shp53 5A1, lanes 6,7; HCC70.shp53 5D4, lanes 8,9 and HCC70. shp53 5D2, lanes 10,11) is shown. Cells were grown in the presence or absence of 8 μg/mL doxycycline (Dox as indicated) for 6 days. Whole-cell lysates were prepared as described in the Materials and Methods section, and 50 μg of protein was separated by gradient 4%–12% SDS/PAGE and analyzed by western blot using antibody to p53. GAPDH was used as a loading control. Expression of mtp53 in parental and control cell line are also shown (lanes 1 and 2 respectively); (b) confocal microscopy images of mtp53 protein were obtained by using anti-p53 antibody. DAPI staining was used to determine the nucleus, and GFP was an indicator of doxycycline-mediated induction. Nuclear immunofluorescence representing mtp53 in the HCC70.shp53 5A1 cell line reduced following doxycycline (top panel) and did not change in the control cell line (bottom panel).
Figure 3.
Inducible depletion of mtp53 proteins R248Q in breast cancer cell line HCC70. (a) The mtp53 expression in whole-cell extracts from parental HCC70 cell line (lane 1) and cell lines expressing vector control (control, lane 2,3) or mtp53-targeted shRNA (lanes 4–11), including HCC70. shp53 italic or not—Must be consistent pool (lane 4,5) and clonal cell lines (HCC70.shp53 5A1, lanes 6,7; HCC70.shp53 5D4, lanes 8,9 and HCC70. shp53 5D2, lanes 10,11) is shown. Cells were grown in the presence or absence of 8 μg/mL doxycycline (Dox as indicated) for 6 days. Whole-cell lysates were prepared as described in the Materials and Methods section, and 50 μg of protein was separated by gradient 4%–12% SDS/PAGE and analyzed by western blot using antibody to p53. GAPDH was used as a loading control. Expression of mtp53 in parental and control cell line are also shown (lanes 1 and 2 respectively); (b) confocal microscopy images of mtp53 protein were obtained by using anti-p53 antibody. DAPI staining was used to determine the nucleus, and GFP was an indicator of doxycycline-mediated induction. Nuclear immunofluorescence representing mtp53 in the HCC70.shp53 5A1 cell line reduced following doxycycline (top panel) and did not change in the control cell line (bottom panel).
![Ijerph 13 00022 g003]()
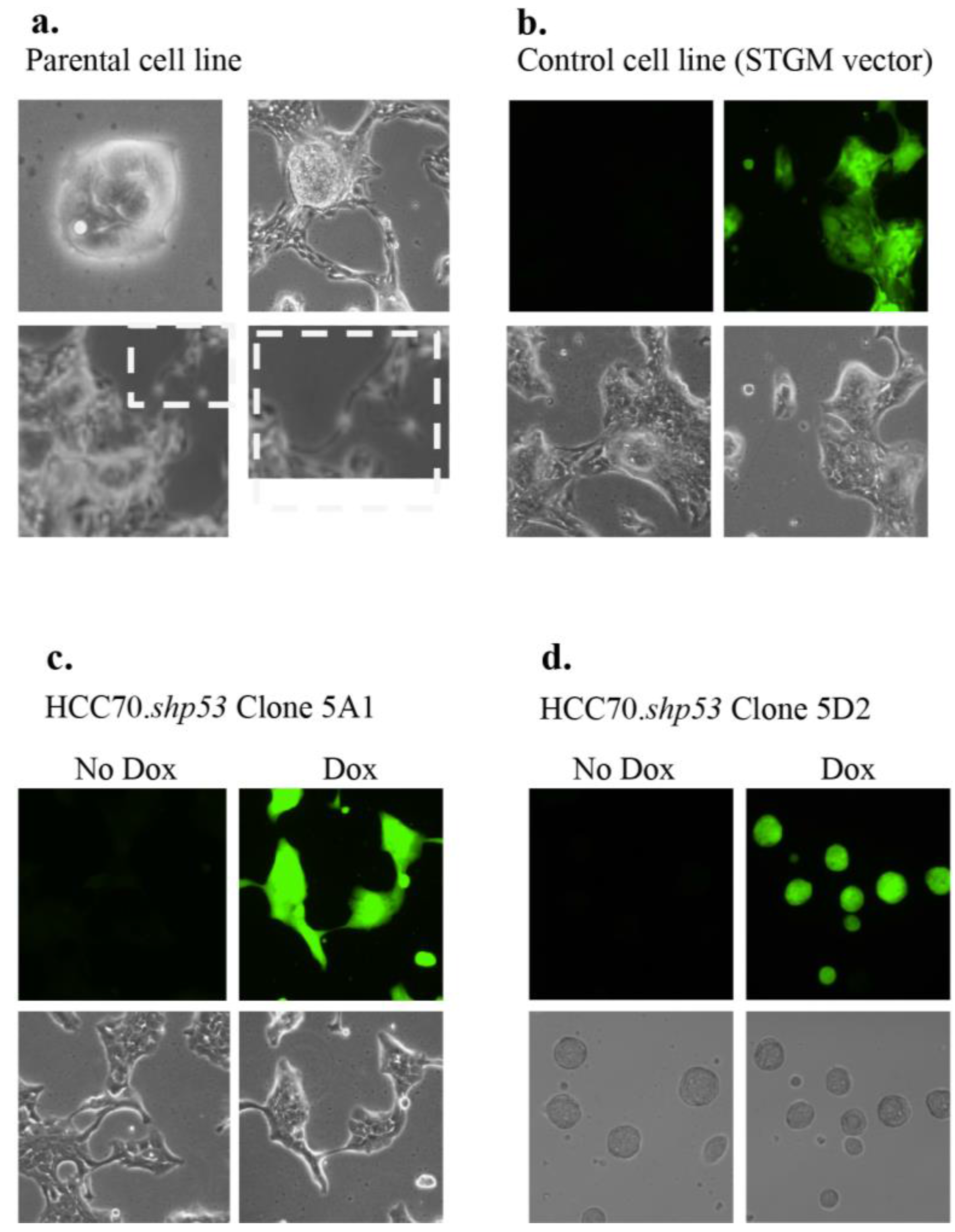
The HCC70 cell line morphology was difficult to characterize and depended on the density of the cells (
Figure 4a). When sparsely plated, the HCC70 parental cell line exhibited cells densely packed together with no distinguishable borders between them. Single cells shed off of the colonies and once a higher density was achieved the outer layer of cells in each colony branched out to contact the neighboring colony, while the central part of the colony continued to form a densely packed group of cells (
Figure 4a). The cell morphology in cells infected with STGM control vector was the same as the parental cell line with or without Dox (
Figure 4b). During establishing of clonal cell lines from HCC70. shp53
. pool culture we isolated two morphologically distinct clones (5A1 and 5D2), which collectively represented the mixed morphology of the parental cell line (
Figure 4c,d). The knockdown of mtp53 did not influence the cell morphology.
Figure 4.
Morphology of HCC70.shp53 5A1 and 5D2 diverge from that of parental cell line. (a) Images of light microscopy of live HCC70 parental cell (boxed panel shows increased magnification on the right of the same region boxed on the left) and (b) STGM-control cell lines are shown. Fluorescent microscopy showing GFP indicates that Dox was added to the culture conditions. In the STGM-control there was no mtp53 knockdown. During cloning of the HCC70.shp53 pool we isolated clones that were representative of each type of morphology. (c) Specifically, HCC70.shp53 5A1 exhibited branched colony structures only (bottom left) and (d) HCC70.shp53 5D2—stem cell-like round colony morphology.
Figure 4.
Morphology of HCC70.shp53 5A1 and 5D2 diverge from that of parental cell line. (a) Images of light microscopy of live HCC70 parental cell (boxed panel shows increased magnification on the right of the same region boxed on the left) and (b) STGM-control cell lines are shown. Fluorescent microscopy showing GFP indicates that Dox was added to the culture conditions. In the STGM-control there was no mtp53 knockdown. During cloning of the HCC70.shp53 pool we isolated clones that were representative of each type of morphology. (c) Specifically, HCC70.shp53 5A1 exhibited branched colony structures only (bottom left) and (d) HCC70.shp53 5D2—stem cell-like round colony morphology.
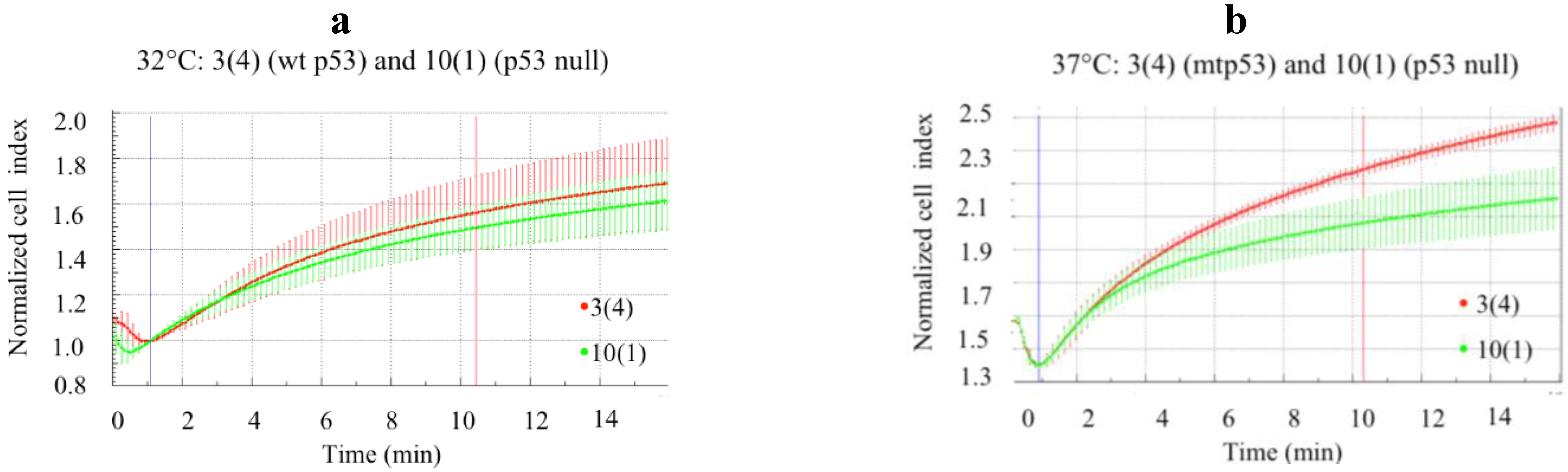
3.4. Deformability of the HCC70 Cell Line Depends on Expression of mtp53 R248Q and on the Morphology of the Cells
The clonal cell lines were assessed for impedance changes during hyposmotic pressure to see if HCC70.shp53 recapitulated the reduction of mtp53 resulting in increased stiffness [
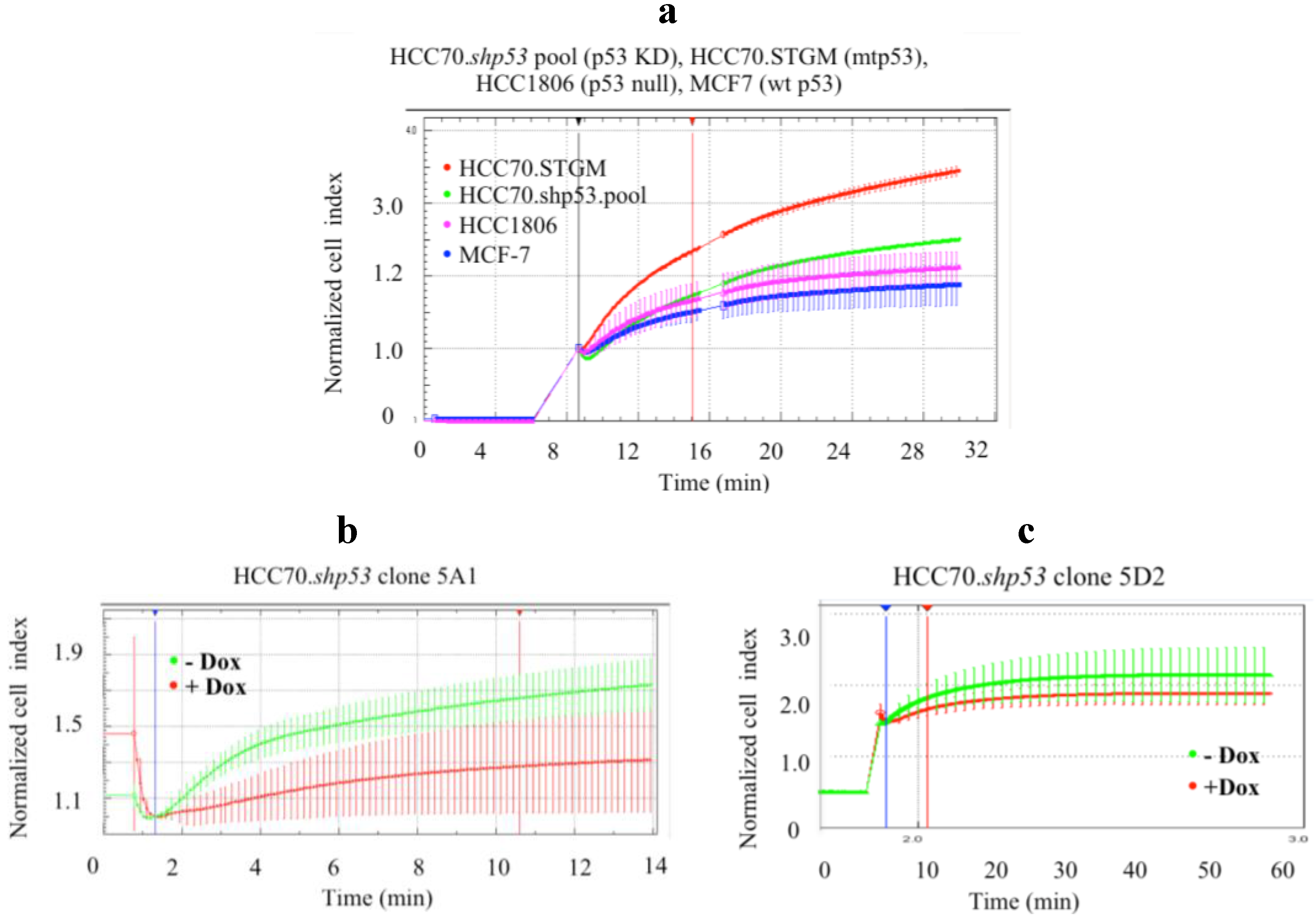
9]. In the pool population of HCC70.shp53 we observed decreased deformability when compared to the STGM control cell line (
Figure 5a). Prior to mtp53 knockdown the HCC70.shp53 cells were less deformable that either MCF-7 or HCC1806 (
Figure 5a). The knockdown of R248Q in HCC70.shp53 clone 5A1 decreased cell deformability (
Figure 5b). However the knockdown of R248Q mtp53 in clone 5D2 did not detectably influence cell deformability (
Figure 5c). Therefore clonal differences acting together with p53 knockdown caused variability in the outcomes.
Figure 5.
Cell deformability decreased upon mtp53 KD in pool population and in clonal cell lines with branched mesenchymal-like morphology. (a) Deformability (Normalized Cell Index) of HCC70 control cell line (red) or HCC70.shp53 pool (green), treated with dox, was compared to MCF7 (blue) and HCC1806 (fuchsia) cell lines, as described in the Materials and Methods section. Following mtp53 knockdown in HCC70.shp53 pool cell line change in CI during swelling was significantly reduced compared to the HCC70 control cell line. (b,c) Average variations in deformability of clonal cell lines HCC70.shp53 5A1 (b) and 5D2 (c) were compared prior and following mtp53 knockdown (green and red, respectively). Mtp53 knockdown in (b) HCC70.shp53 clone 5A1 significantly decreased the ability of cells to swell (c) while in the clone 5D2 very little impedance change was detected. Average values and standard deviation at each time points are shown (n = 4). The first vertical line indicates the addition of water. The second vertical line (shown in red) was used to generate slopes of the curves (data not shown).
Figure 5.
Cell deformability decreased upon mtp53 KD in pool population and in clonal cell lines with branched mesenchymal-like morphology. (a) Deformability (Normalized Cell Index) of HCC70 control cell line (red) or HCC70.shp53 pool (green), treated with dox, was compared to MCF7 (blue) and HCC1806 (fuchsia) cell lines, as described in the Materials and Methods section. Following mtp53 knockdown in HCC70.shp53 pool cell line change in CI during swelling was significantly reduced compared to the HCC70 control cell line. (b,c) Average variations in deformability of clonal cell lines HCC70.shp53 5A1 (b) and 5D2 (c) were compared prior and following mtp53 knockdown (green and red, respectively). Mtp53 knockdown in (b) HCC70.shp53 clone 5A1 significantly decreased the ability of cells to swell (c) while in the clone 5D2 very little impedance change was detected. Average values and standard deviation at each time points are shown (n = 4). The first vertical line indicates the addition of water. The second vertical line (shown in red) was used to generate slopes of the curves (data not shown).
![Ijerph 13 00022 g005]()
3.6. The Knockdown of mtp53 R248Q in HCC70 Changes Cell Morphology
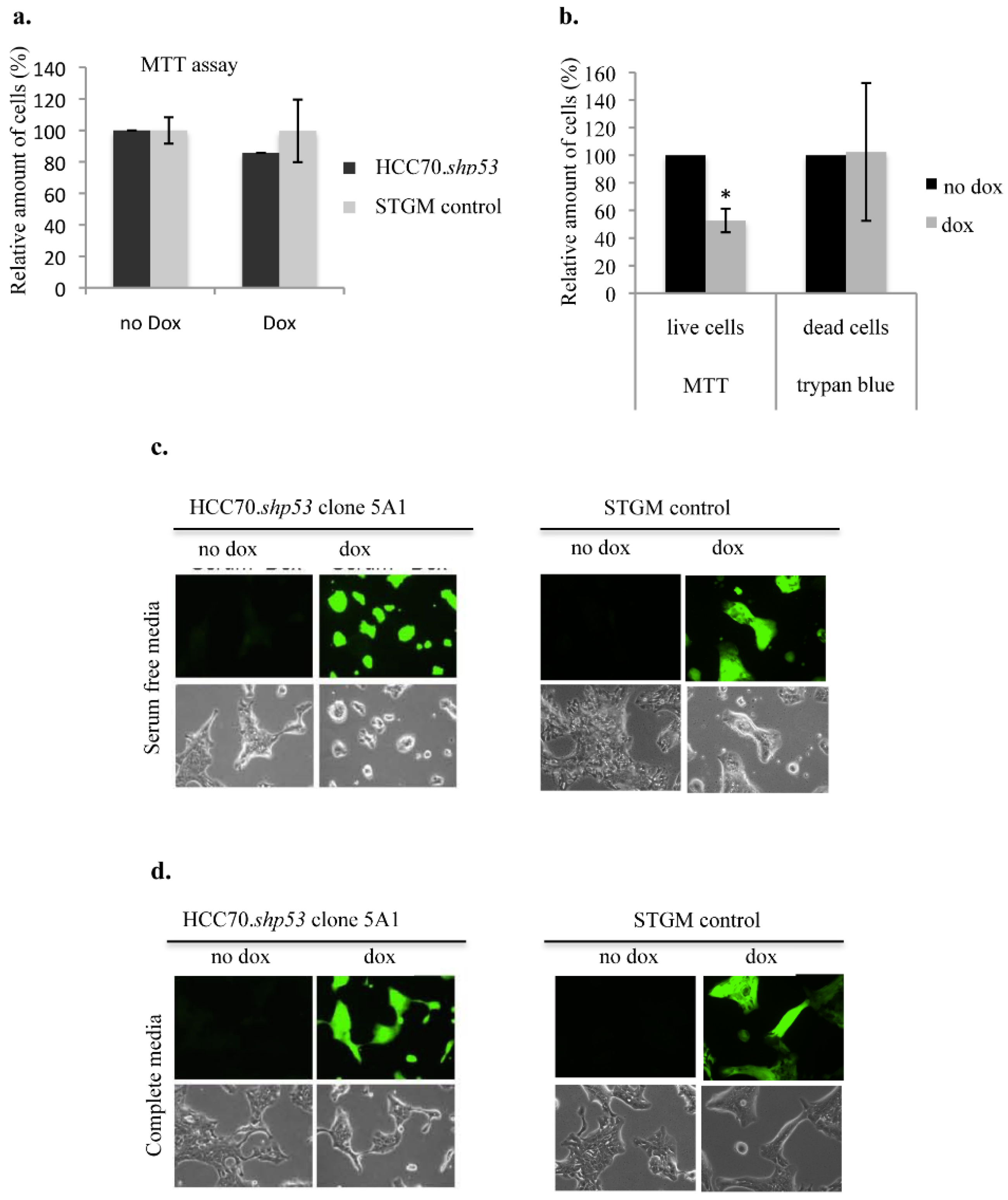
We examined the ability of mtp53 R248Q to confer a survival advantage on HCC70 under low serum stressful conditions (
Figure 7). We first tested if knockdown of mtp53 R248Q (by doxycycline treatment) under normal cell culture conditions (cell culture media conditioned with 10% FBS) reduced cell viability in the HCC70.shp53 pool cell line. Doxycycline (Dox) treatment did not reduce the metabolic activity of the mitochondria as shown by MTT assay (
Figure 7a). We then examined the influence of serum deprivation on clone 5A1 that showed a decrease in flexibility with mtp53 knockdown. Serum deprivation conditions for the HCC70.shp53 clone 5A1 with p53 knockdown decreased mitochondrial activity but did not influence the trypan blue staining (
Figure 7b). This suggested that mtp53 knockdown decreased the metabolic activity of the cells but did not increase cell death. We observed that the morphology of serum starved HCC70.shp53 with p53 knockdown became more round compared to the non-treated cells (
Figure 7c). There was no difference observed for the serum starved STGM control cell line treated with Dox (
Figure 7c). Finally under normal serum conditions the treatment of HCC70.shp53 cells with Dox did not influence the cell morphology (
Figure 7d). Therefore the lack of serum in the absence of mtp53 expression was a contributor for the loss of the mtp53-associated morphology phenotype and decreased the mitochondrial activity.
Figure 7.
Loss of branched colony morphology following mtp53 knockdown during culture in serum free media. (a) MTT assay of the cell viability following dox treatment in HCC70.shp53 and STGM-control cell lines (black and grey bars, respectively). (b,c) Cell morphology was assessed in serum deprivation conditions following mtp53 KD. (b) MTT assay and trypan blue in HCC70.shp53 cells following serum starvation and with a comparison between the cells prior and following mtp53 knockdown (black and grey bars, receptively). (c) Images of light and fluorescent microscopy of HCC70.shp53 clone 5A1 and STGM control treated with or without dox and cultures in serum deprived conditions. (d) Images of light and fluorescent microscopy of HCC70.shp53 clone 5A1 and STGM control treated with or without dox and cultures in complete medium.
Figure 7.
Loss of branched colony morphology following mtp53 knockdown during culture in serum free media. (a) MTT assay of the cell viability following dox treatment in HCC70.shp53 and STGM-control cell lines (black and grey bars, respectively). (b,c) Cell morphology was assessed in serum deprivation conditions following mtp53 KD. (b) MTT assay and trypan blue in HCC70.shp53 cells following serum starvation and with a comparison between the cells prior and following mtp53 knockdown (black and grey bars, receptively). (c) Images of light and fluorescent microscopy of HCC70.shp53 clone 5A1 and STGM control treated with or without dox and cultures in serum deprived conditions. (d) Images of light and fluorescent microscopy of HCC70.shp53 clone 5A1 and STGM control treated with or without dox and cultures in complete medium.
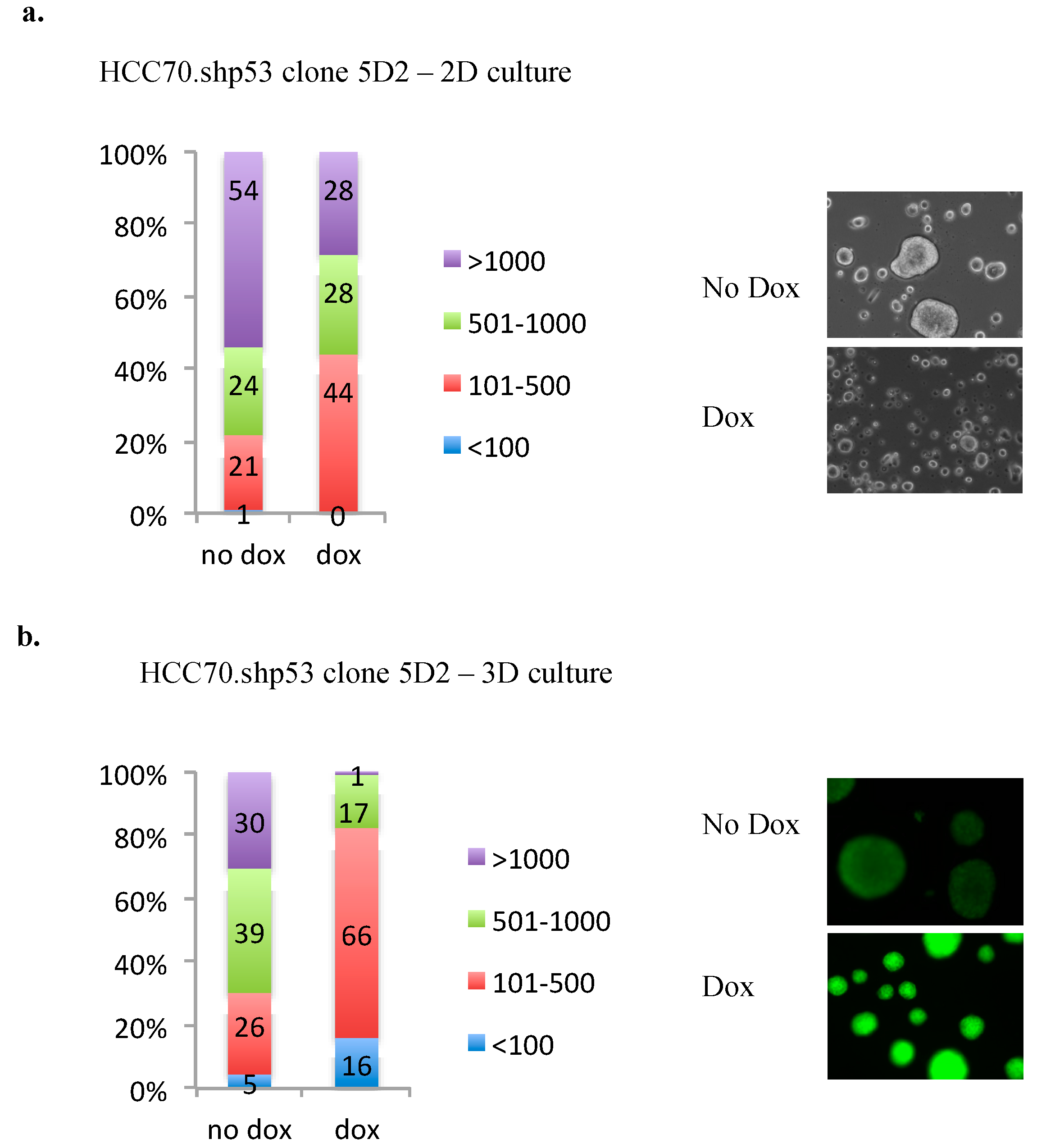
3.7. Knockdown of mtp53 in HCC70.shp53 5D2 Decrease Colony Size and PARP on the Chromatin
The HCC70.shp53 5A1 and 5D2 clones demonstrated some differences in flexibility and morphology. We were interested to determine if there were mtp53 associated phenotypes that could be detected in HCC70.shp53 5D2 that associated with mtp53 actions in a different clonal background. The 5D2 cells showed an interesting colony formation phenotype. We therefore asked if the 5D2 clone exhibited any of the other phenotypes previously associated with mtp53 R273H expression in MDA-MB-468 cells. MDA-468. shp53 cells show a mtp53-driven PARP association with the chromatin [
14] and increased colony size when grown in 3D culture [
13]. The knockdown of mtp53 in HCC70.shp53 5D2 reduced the colony size during cell culture in either 2-dimensions (2D) or 3-dimensions (3D) (
Figure 8a,b). These colony formation assays were only carried out with 5D2 and not 5A1, in order to determine if there was a mtp53 associated phenotype in 5D2 cells.
Figure 8.
HCC70 colony size is reduced by the knockdown of R248Q mtp53. (a) Colony size of HCC70.shp53 clone 5D2 treated with or without Dox, was quantified using ImageJ software and presented as a distribution of size grouped randomly into <100 (blue), 101–500 (red), 501–1000 (green) and >1000 (purple). 100 colonies were used in quantification and % colonies in each size group are presented in a bar graph. Representative light and fluorescent microscopy images are shown. (b) Colony size of HCC70.shp53 clone 5D2 treated with or without Dox and cultured on matrigel for 3D colony formation, was quantified using ImageJ software and presented as a distribution of size grouped randomly into <100 (blue), 101–500 (red), 501–1000 (green) and >1000 (purple). 100 colonies were used in quantification and % colonies in each size group are presented in a bar graph.
Figure 8.
HCC70 colony size is reduced by the knockdown of R248Q mtp53. (a) Colony size of HCC70.shp53 clone 5D2 treated with or without Dox, was quantified using ImageJ software and presented as a distribution of size grouped randomly into <100 (blue), 101–500 (red), 501–1000 (green) and >1000 (purple). 100 colonies were used in quantification and % colonies in each size group are presented in a bar graph. Representative light and fluorescent microscopy images are shown. (b) Colony size of HCC70.shp53 clone 5D2 treated with or without Dox and cultured on matrigel for 3D colony formation, was quantified using ImageJ software and presented as a distribution of size grouped randomly into <100 (blue), 101–500 (red), 501–1000 (green) and >1000 (purple). 100 colonies were used in quantification and % colonies in each size group are presented in a bar graph.
In order to address a previously reported nuclear cytoplasmic associated mutant p53 gain-of-function property we addressed the level and localization of PARP [
14]. We previously observed many different bands and variable mobility of mutant p53 in the cytoplasmic and chromatin fractions when they are analyzed on a gradient gel; these bands suggest different p53 post-translational modifications [
14]. Furthermore, knockdown of R273H in MDA-MB-468 cells causes PARP to move from the nucleus to the cytoplasm [
14]. Depletion of R248Q in the 5A1 and 5D2 cells caused PARP1 to be reduced on the chromatin (
Figure 9, compare lane 5 to 6 and lane 7 and 8). Image J analysis demonstrated a greater reduction of chromatin associated PARP1 in 5A1 cells following mtp53 knockdown. Interestingly clone 5D2 showed a greater increase in cytoplasmic PARP1 when mtp53 was depleted, and this correlated with our past findings for R273H (compare lane 3 to 4). Therefore the mtp53 depletion phenotypes of increased cytoplasmic PARP and decreased flexibility are not directly related.
Figure 9.
Depletion of mtp53 modulates PARP expression. HCC70.
shp53 clones 5A1 and 5D2 were grown in the presence or absence of 6 μg/mL of doxycycline for 7 days and fractionation was carried out. Samples were resolved on a gradient 4%–12% SDS/PAGE which resulted in the observation of differentially migrating p53 in cytoplasmic and chromatin fractions [
14]. A total of 50 μg of protein from the cytoplasmic fraction and 10 μg of the chromatin fraction per lane were resolved. Protein levels of p53, PARP1, actin, and fibrillarin in the fractions were determined by western blot analysis and the results were quantified by ImageJ. Actin was used to normalize the cytoplasmic fractions and fibrillarin was used to normalize the chromatin fraction (relative expression values comparing with and without knockdown are shown under the PARP1 and mtp53 western blots).
Figure 9.
Depletion of mtp53 modulates PARP expression. HCC70.
shp53 clones 5A1 and 5D2 were grown in the presence or absence of 6 μg/mL of doxycycline for 7 days and fractionation was carried out. Samples were resolved on a gradient 4%–12% SDS/PAGE which resulted in the observation of differentially migrating p53 in cytoplasmic and chromatin fractions [
14]. A total of 50 μg of protein from the cytoplasmic fraction and 10 μg of the chromatin fraction per lane were resolved. Protein levels of p53, PARP1, actin, and fibrillarin in the fractions were determined by western blot analysis and the results were quantified by ImageJ. Actin was used to normalize the cytoplasmic fractions and fibrillarin was used to normalize the chromatin fraction (relative expression values comparing with and without knockdown are shown under the PARP1 and mtp53 western blots).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








