
Effects of Asian Dust Particles on the Early-Stage Antigen-Induced Immune Response of Asthma in NC/Nga Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of AD Particles
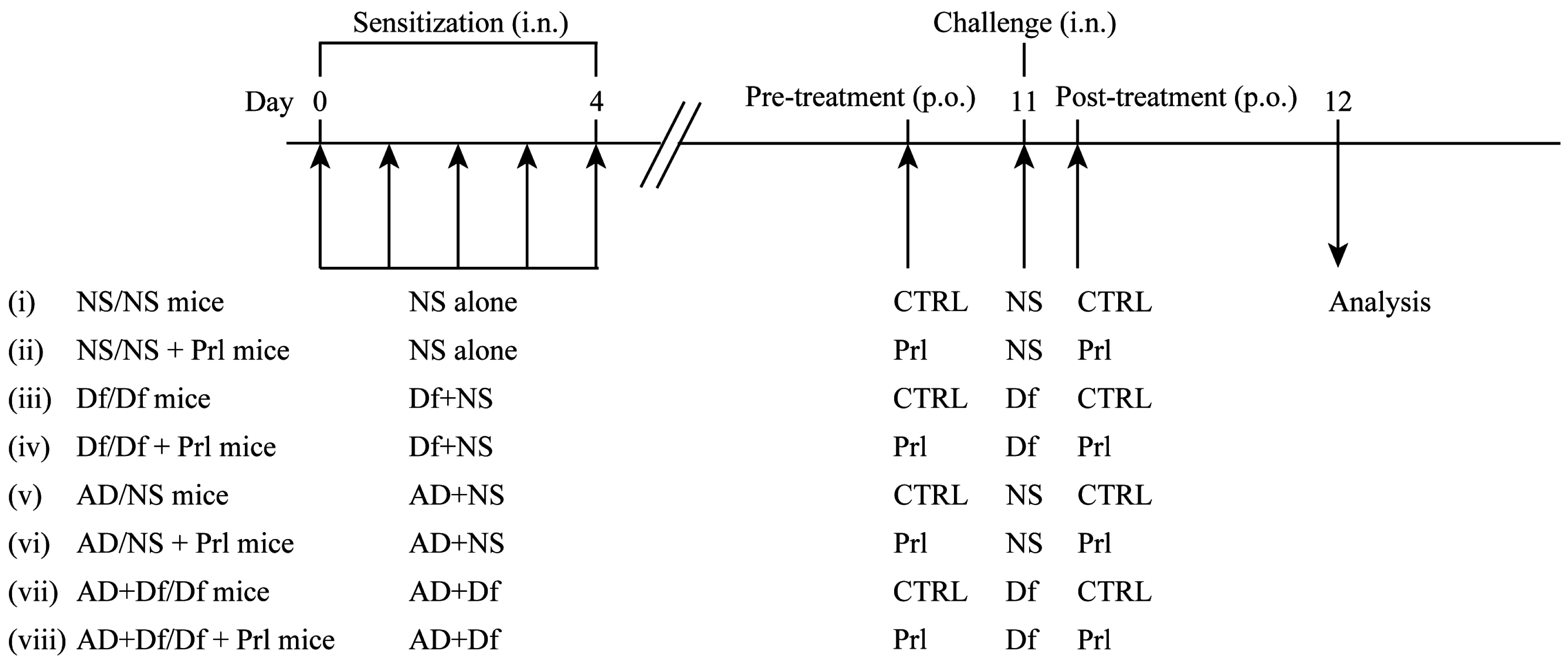
2.3. Experimental Protocol
2.4. BALF Procedure
2.5. Histological Examination
2.6. Quantitative Determination of Cytokine, Chemokine, and LT Levels
2.7. Statistical Analysis
3. Results
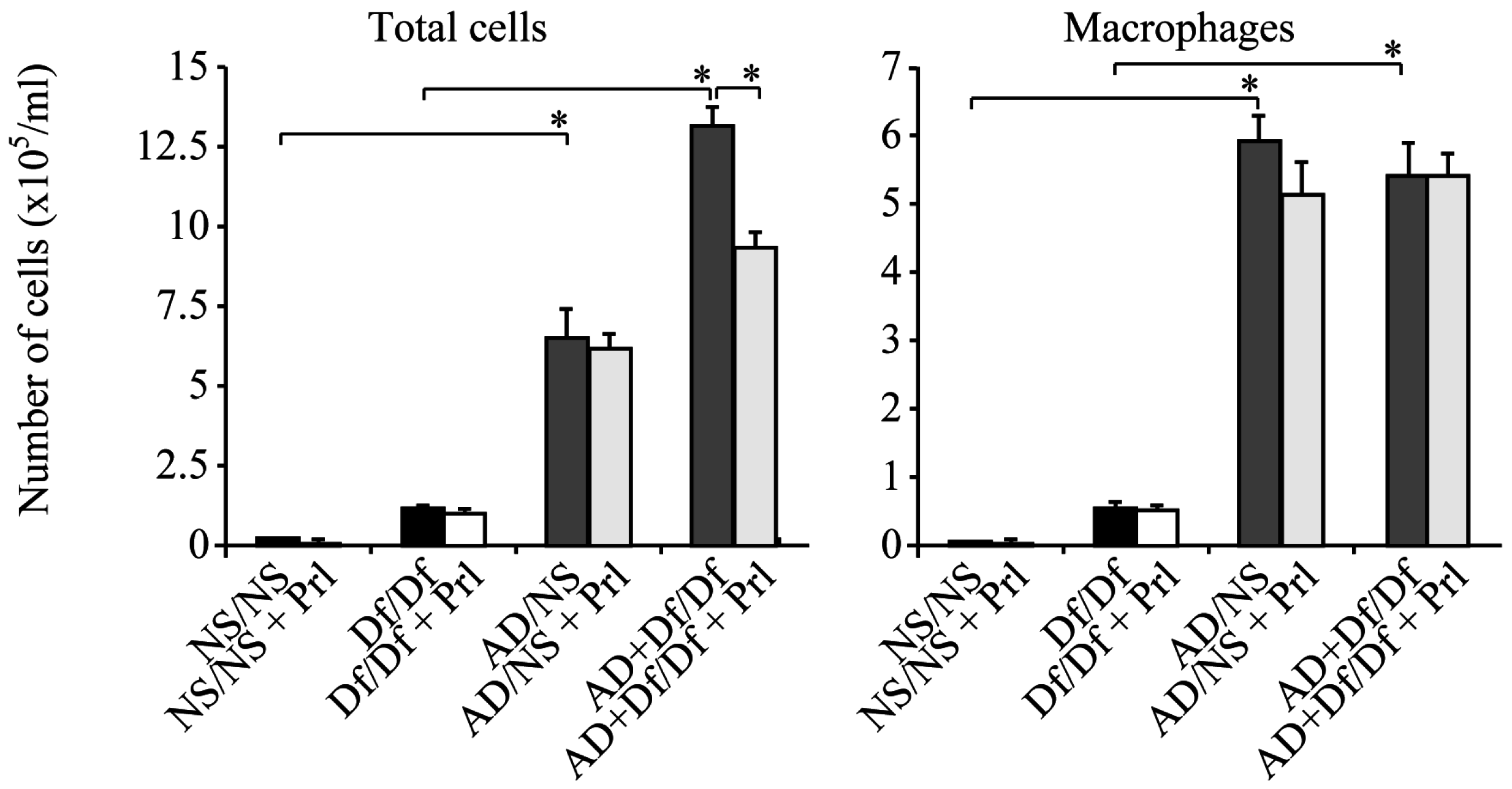
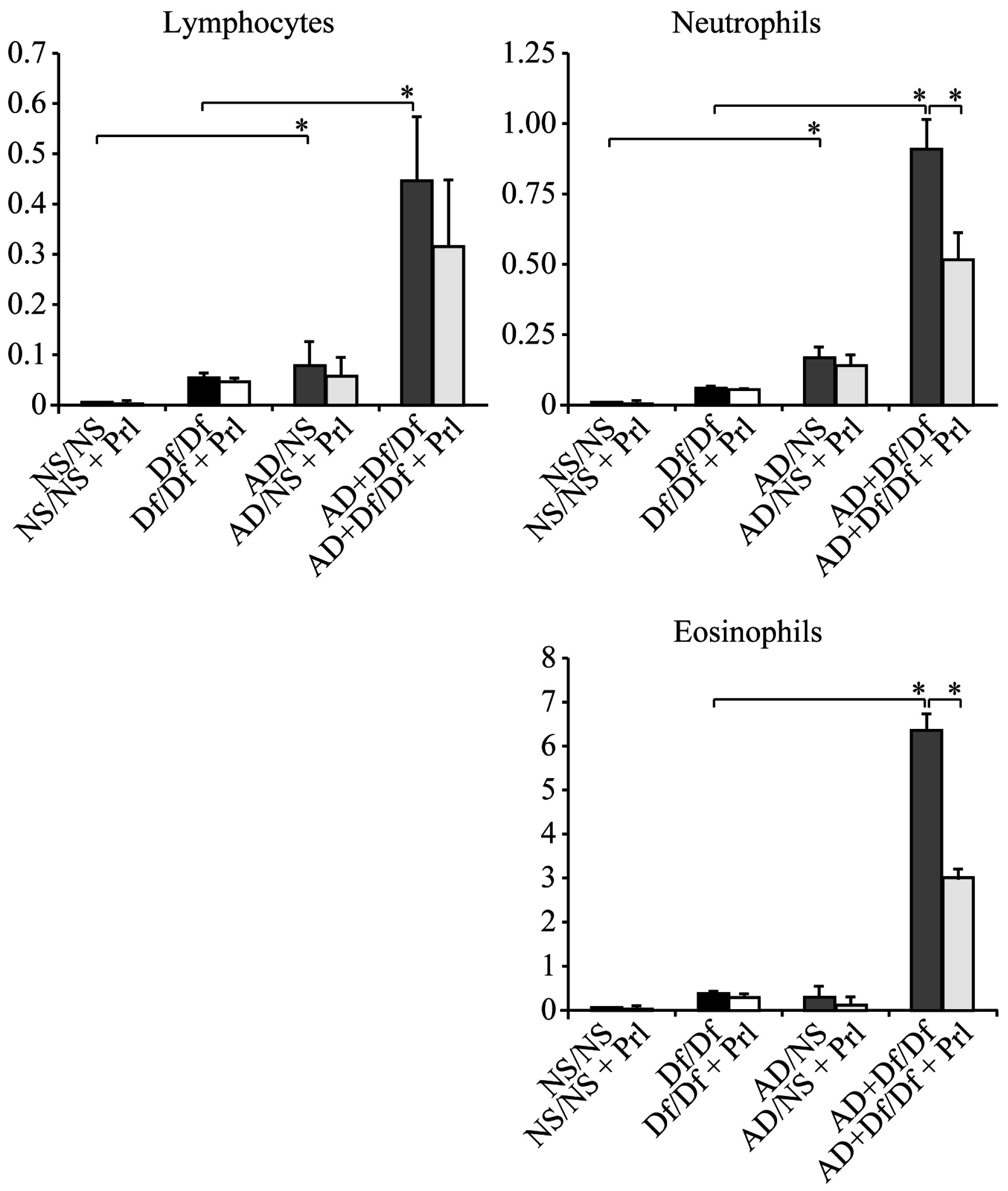
3.1. Cell Counts in BALF
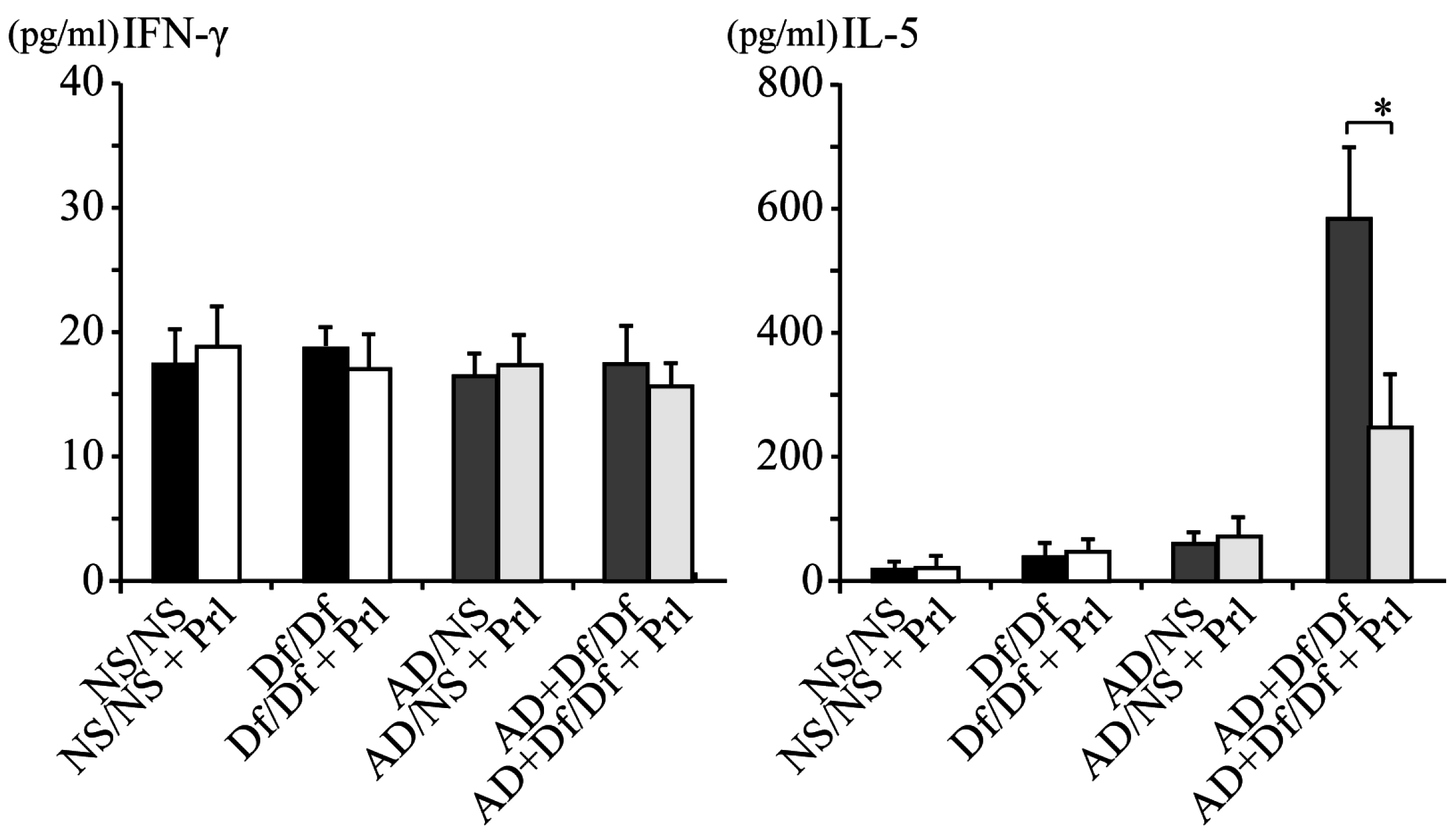
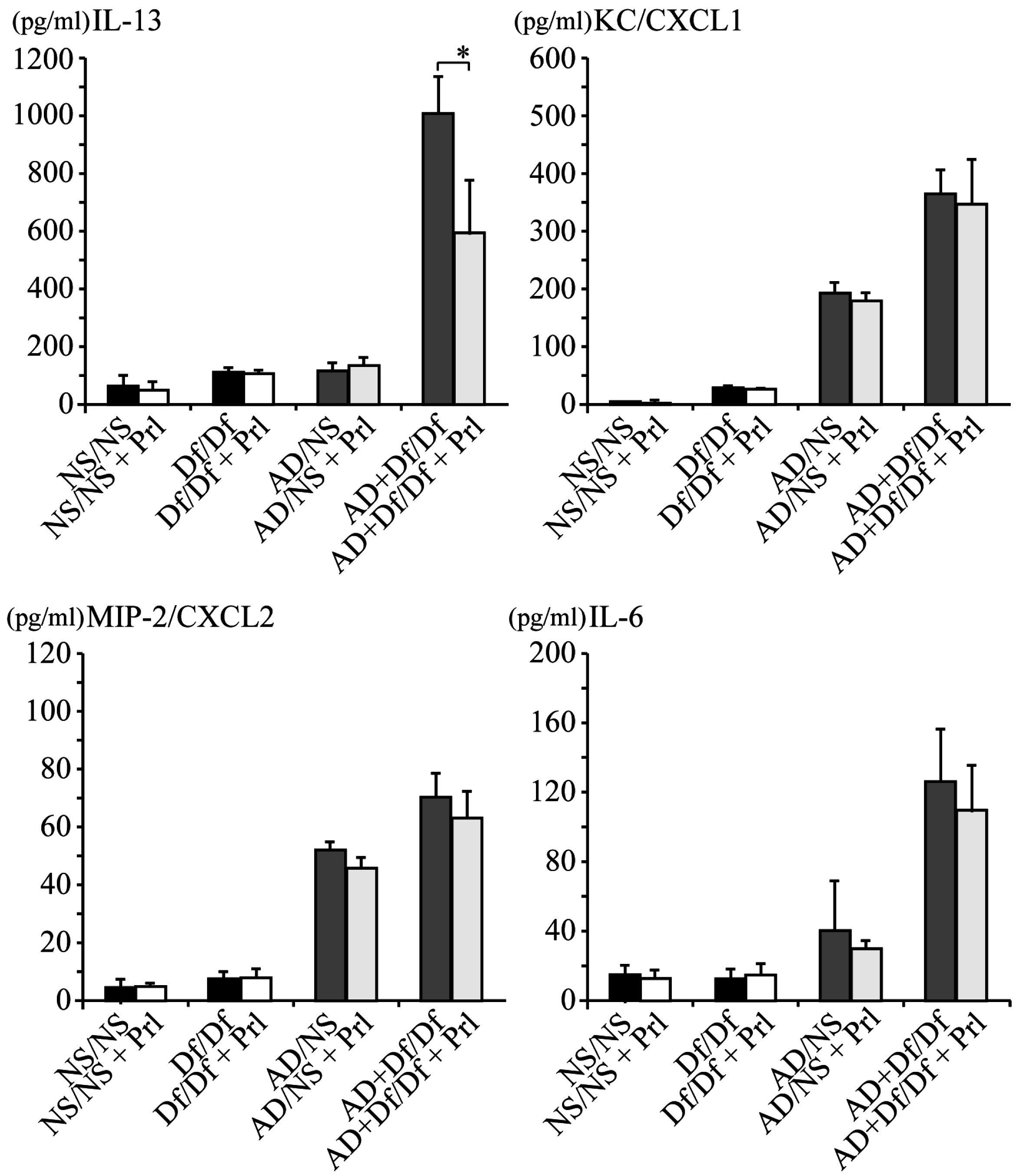
3.2. Cytokine Profile of BALF
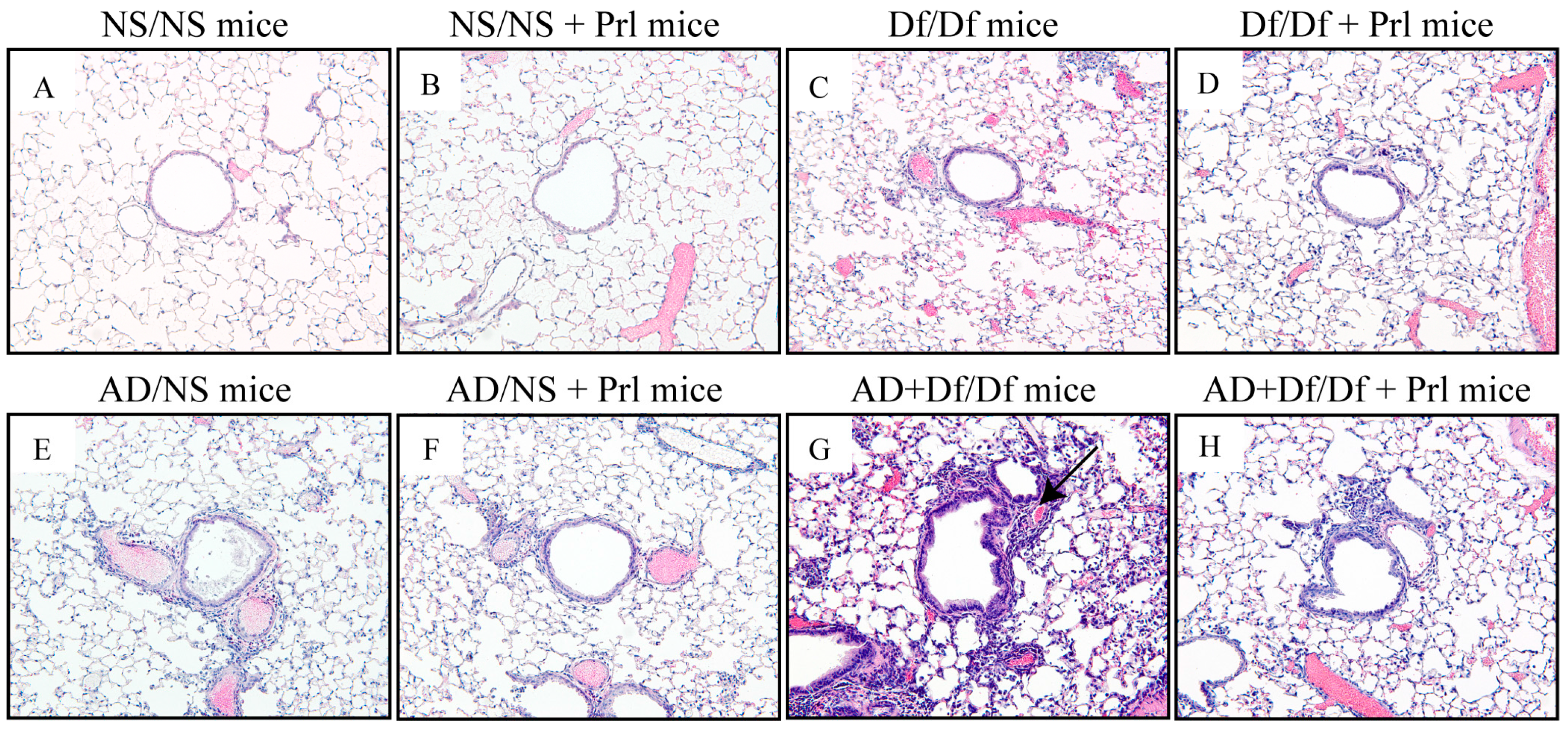
3.3. Histopathological Changes in the Lung
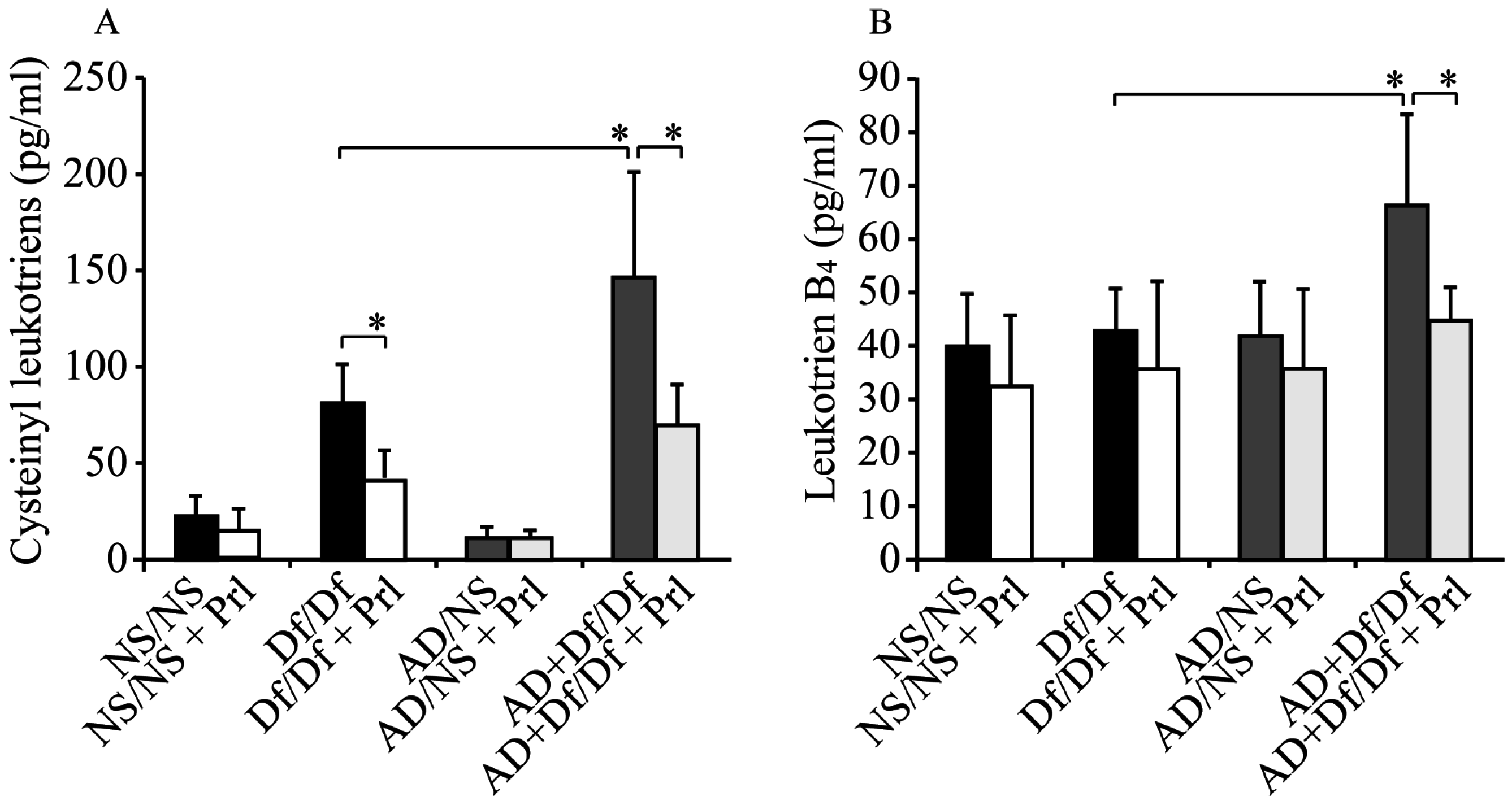
3.4. Measurements of CysLTs and LTB4 in BALF
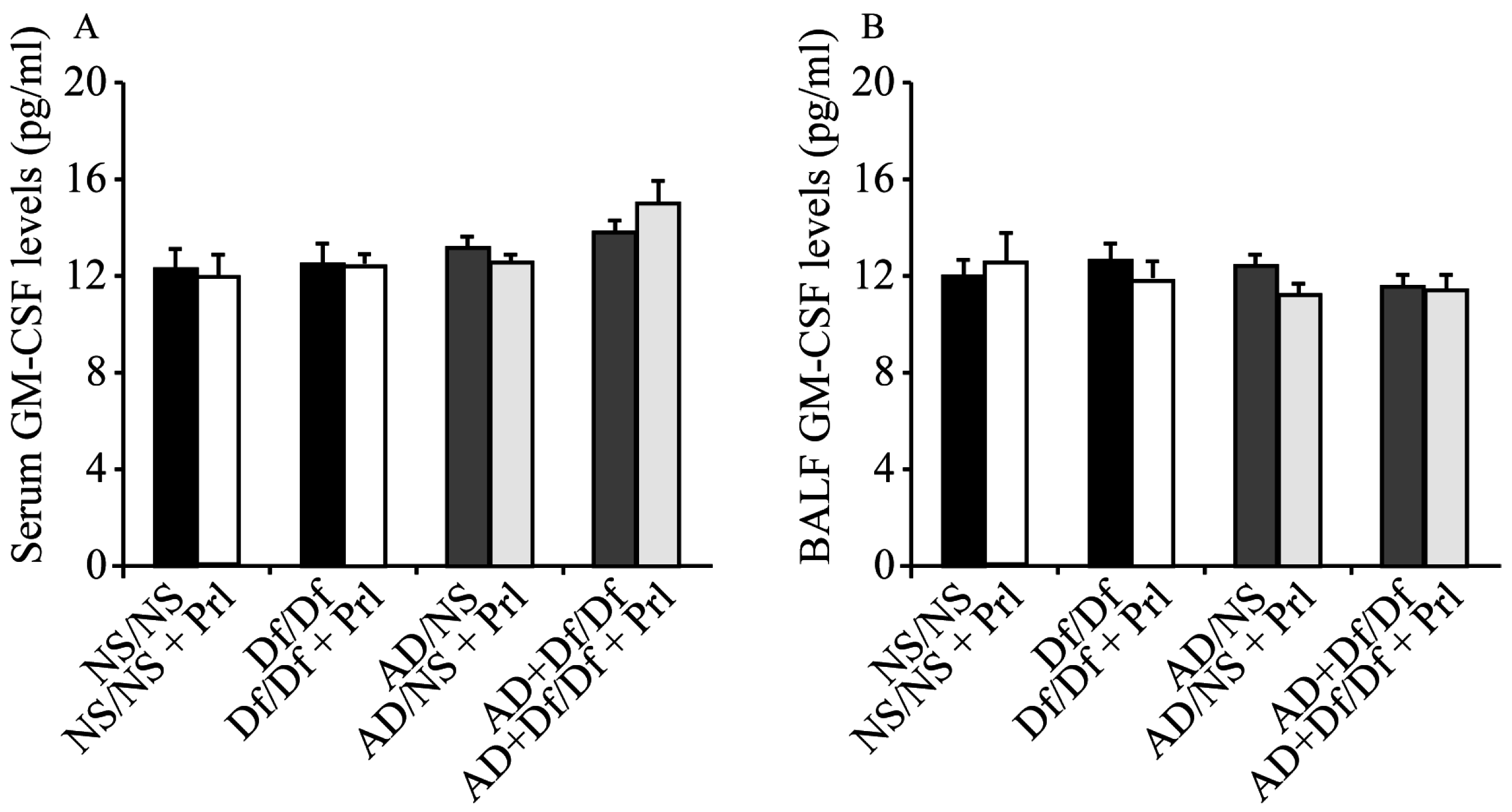
3.5. Measurement of GM-CSF in Serum and BALF
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AD | Asian dust |
| BALF | bronchoalveolar lavage fluid |
| CMC | carboxymethylcellulose sodium |
| CysLT1 | type 1 cysteinyl leukotriene |
| Df | Dermatophagoides farina |
| EIA | enzyme immunoassay |
| GM-CSF | Granulocyte-macrophage colony stimulating factor |
| H&E | hematoxylin and eosin |
| IFN | interferon |
| IL | interleukin |
| LT | leukotriene |
| LTB4 | Dihydroxy-LT |
| NS | normal saline |
| PM | particulate matter |
| PRL | Pranlukast |
| SD | standard deviation |
References
- Dockery, D.W.; Pope, C.A.; Xu, X.; Spengler, J.D.; Ware, J.H.; Fay, M.E.; Ferris, B.G., Jr.; Speizer, F.E. An association between air pollution and mortality in six U.S. Cities. N. Engl. J. Med. 1993, 329, 1753–1759. [Google Scholar] [CrossRef] [PubMed]
- Ware, J.H.; Thibodeau, L.A.; Speizer, F.E.; Colome, S.; Ferris, B.G., Jr. Assessment of the health effects of atmospheric sulfur oxides and particulate matter: Evidence from observational studies. Environ. Health Perspect. 1981, 41, 255–276. [Google Scholar] [CrossRef] [PubMed]
- Goudie, A.S. Desert dust and human health disorders. Environ. Int. 2014, 63, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.; Ramanathan, V.; Li, F.; Kim, D. Dust plumes over the Pacific, Indian, and Atlantic Oceans: Climatology and radiative impact. J. Geophys. Res.-Atmos. 2007. [Google Scholar] [CrossRef]
- Tanaka, T.Y.; Chiba, M. A numerical study of the contributions of dust source regions to the global dust budget. Glob. Planet Chang. 2006, 52, 88–104. [Google Scholar] [CrossRef]
- Sugimoto, N.; Lee, C.H. Characteristics of dust aerosols inferred from lidar depolarization measurements at two wavelengths. Appl. Opt. 2006, 45, 7468–7474. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, N.; Matsui, I.; Shimizu, A.; Nishizawa, T.; Hara, Y.; Xie, C.; Uno, I.; Yumimoto, K.; Wang, Z.; Yoon, S. Lidar network observations of tropospheric aerosols. Proc. SPIE 2008. [Google Scholar] [CrossRef]
- Hashizume, M.; Ueda, K.; Nishiwaki, Y.; Michikawa, T.; Onozuka, D. Health effects of asian dust events: A review of the literature. Nihon Eiseigaku Zasshi 2010, 65, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Igishi, T.; Burioka, N.; Yamasaki, A.; Kurai, J.; Takeuchi, H.; Sako, T.; Yoshida, A.; Yoneda, K.; Fukuoka, Y.; et al. Pollen augments the influence of desert dust on symptoms of adult asthma patients. Allergol. Int. 2011, 60, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Yamasaki, A.; Burioka, N.; Kurai, J.; Yoneda, K.; Yoshida, A.; Igishi, T.; Fukuoka, Y.; Nakamoto, M.; Takeuchi, H.; et al. Correlation between asian dust storms and worsening asthma in western Japan. Allergol. Int. 2011, 60, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Kurai, J.; Tomita, K.; Sano, H.; Abe, S.; Saito, R.; Minato, S.; Igishi, T.; Burioka, N.; Sako, T.; et al. Effects on asthma and induction of interleukin-8 caused by Asian dust particles collected in western Japan. J. Asthma 2014, 6, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Noma, H.; Kurai, J.; Sano, H.; Saito, R.; Abe, S.; Kimura, Y.; Aiba, S.; Oshimura, M.; Yamasaki, A.; et al. Decreased pulmonary function in school children in western Japan after exposures to Asian desert dusts and its association with interleukin-8. BioMed Res. Int. 2015, 2015, 583293. [Google Scholar] [CrossRef] [PubMed]
- Kanatani, K.T.; Ito, I.; Al-Delaimy, W.K.; Adachi, Y.; Mathews, W.C.; Ramsdell, J.W. Desert dust exposure is associated with increased risk of asthma hospitalization in children. Am. J. Respir. Crit. Care Med. 2010, 182, 1475–1481. [Google Scholar] [CrossRef] [PubMed]
- Ueda, K.; Nitta, H.; Odajima, H. The effects of weather, air pollutants, and Asian dust on hospitalization for asthma in Fukuoka. Environ. Health Prev. Med. 2010, 15, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Yoo, Y.; Choung, J.T.; Yu, J.; Kim do, K.; Koh, Y.Y. Acute effects of asian dust events on respiratory symptoms and peak expiratory flow in children with mild asthma. J. Korean Med. Sci. 2008, 23, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Gehring, U.; Wijga, A.H.; Hoek, G.; Bellander, T.; Berdel, D.; Bruske, I.; Fuertes, E.; Gruzieva, O.; Heinrich, J.; Hoffmann, B.; et al. Exposure to air pollution and development of asthma and rhinoconjunctivitis throughout childhood and adolescence: A population-based birth cohort study. Lancet Respir. Med. 2015, 3, 933–942. [Google Scholar] [CrossRef]
- Gehring, U.; Wijga, A.H.; Brauer, M.; Fischer, P.; de Jongste, J.C.; Kerkhof, M.; Oldenwening, M.; Smit, H.A.; Brunekreef, B. Traffic-related air pollution and the development of asthma and allergies during the first 8 years of life. Am. J. Respir. Crit. Care Med. 2010, 181, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Salmon, J.A.; Higgs, G.A. Prostaglandins and leukotrienes as inflammatory mediators. Br. Med. Bull. 1987, 43, 285–296. [Google Scholar] [PubMed]
- Peters-Golden, M.; Henderson, W.R., Jr. Leukotrienes. N. Engl. J. Med. 2007, 357, 1841–1854. [Google Scholar] [CrossRef] [PubMed]
- Brink, C.; Dahlen, S.E.; Drazen, J.; Evans, J.F.; Hay, D.W.; Nicosia, S.; Serhan, C.N.; Shimizu, T.; Yokomizo, T. International Union of Pharmacology XXXVII. Nomenclature for leukotriene and lipoxin receptors. Pharmacol. Rev. 2003, 55, 195–227. [Google Scholar] [CrossRef] [PubMed]
- Hicks, A.; Monkarsh, S.P.; Hoffman, A.F.; Goodnow, R., Jr. Leukotriene b4 receptor antagonists as therapeutics for inflammatory disease: Preclinical and clinical developments. Expert Opin. Investig. Drugs 2007, 16, 1909–1920. [Google Scholar] [CrossRef] [PubMed]
- Miyahara, N.; Miyahara, S.; Takeda, K.; Gelfand, E.W. Role of the LTB4/BLT1 pathway in allergen-induced airway hyperresponsiveness and inflammation. Allergol. Int. 2006, 55, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Yokomizo, T. The role of leukotrienes in allergic diseases. Allergol. Int. 2015, 64, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Hilberg, T.; Deigner, H.P.; Moller, E.; Claus, R.A.; Ruryk, A.; Glaser, D.; Landre, J.; Brunkhorst, F.M.; Reinhart, K.; Gabriel, H.H.; et al. Transcription in response to physical stress—Clues to the molecular mechanisms of exercise-induced asthma. FASEB J. 2005, 19, 1492–1494. [Google Scholar] [CrossRef] [PubMed]
- Malireddy, S.; Lawson, C.; Steinhour, E.; Hart, J.; Kotha, S.R.; Patel, R.B.; Zhao, L.; Wilkins, J.R.; Marsh, C.B.; Magalang, U.J.; et al. Airborne agricultural particulate matter induces inflammatory cytokine secretion by respiratory epithelial cells: Mechanisms of regulation by eicosanoid lipid signal mediators. Indian J. Biochem. Biophys. 2013, 50, 387–401. [Google Scholar] [PubMed]
- Ichinose, T.; Hiyoshi, K.; Yoshida, S.; Takano, H.; Inoue, K.; Nishikawa, M.; Mori, I.; Kawazato, H.; Yasuda, A.; Shibamoto, T. Asian sand dust aggravates allergic rhinitis in guinea pigs induced by Japanese cedar pollen. Inhal. Toxicol. 2009, 21, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Rabinovitch, N.; Strand, M.; Gelfand, E.W. Particulate levels are associated with early asthma worsening in children with persistent disease. Am. J. Respir. Crit. Care Med. 2006, 173, 1098–1105. [Google Scholar] [CrossRef] [PubMed]
- Shibamori, M.; Ogino, K.; Kambayashi, Y.; Ishiyama, H. Intranasal mite allergen induces allergic asthma-like responses in NC/NGA mice. Life Sci. 2006, 78, 987–994. [Google Scholar] [CrossRef] [PubMed]
- Nakagome, K.; Matsushita, S.; Nagata, M. Neutrophilic inflammation in severe asthma. Int. Arch. Allergy Immunol. 2012, 158 (Suppl. 1), 96–102. [Google Scholar] [CrossRef] [PubMed]
- Holgate, S.T.; Sandstrom, T.; Frew, A.J.; Stenfors, N.; Nordenhall, C.; Salvi, S.; Blomberg, A.; Helleday, R.; Soderberg, M. Health effects of acute exposure to air pollution. Part I: Healthy and asthmatic subjects exposed to diesel exhaust. Res. Rep. Health Eff. Inst. 2003, 112, 1–30. [Google Scholar]
- Bellido-Casado, J.; Plaza, V.; Perpina, M.; Picado, C.; Bardagi, S.; Martinez-Bru, C.; Torrejon, M. Inflammatory response of rapid onset asthma exacerbation. Arch. Bronconeumol. 2010, 46, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, S.L.; Standiford, T.; Kasahara, K.; Strieter, R.M. Interleukin-8 (IL-8): The major neutrophil chemotactic factor in the lung. Exp. Lung Res. 1991, 17, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Sierra-Vargas, M.P.; Guzman-Grenfell, A.M.; Blanco-Jimenez, S.; Sepulveda-Sanchez, J.D.; Bernabe-Cabanillas, R.M.; Cardenas-Gonzalez, B.; Ceballos, G.; Hicks, J.J. Airborne particulate matter pm2.5 from mexico city affects the generation of reactive oxygen species by blood neutrophils from asthmatics: An in vitro approach. J. Occup. Med. Toxicol. 2009, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- McCreanor, J.; Cullinan, P.; Nieuwenhuijsen, M.J.; Stewart-Evans, J.; Malliarou, E.; Jarup, L.; Harrington, R.; Svartengren, M.; Han, I.K.; Ohman-Strickland, P.; et al. Respiratory effects of exposure to diesel traffic in persons with asthma. N. Engl. J. Med. 2007, 357, 2348–2358. [Google Scholar] [CrossRef] [PubMed]
- Busse, W.W. Leukotrienes and inflammation. Am. J. Respir. Crit. Care Med. 1998, 157, S210–S213. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.R.; Pistorese, B.P.; Chi, E.Y.; Goodman, R.B.; Matthay, M.A. Effects of leukotriene b4 in the human lung. Recruitment of neutrophils into the alveolar spaces without a change in protein permeability. J. Clin. Investig. 1989, 84, 1609–1619. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, I.; Kumar, B.; Schriever, C. Long-term montelukast therapy in moderate to severe COPD—A preliminary observation. Respir. Med. 2004, 98, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Riccioni, G.; Bucciarelli, T.; Mancini, B.; Di Ilio, C.; D’Orazio, N. Antileukotriene drugs: Clinical application, effectiveness and safety. Curr. Med. Chem. 2007, 14, 1966–1977. [Google Scholar] [CrossRef] [PubMed]
- Celik, P.; Sakar, A.; Havlucu, Y.; Yuksel, H.; Turkdogan, P.; Yorgancioglu, A. Short-term effects of montelukast in stable patients with moderate to severe copd. Respir. Med. 2005, 99, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Gueli, N.; Verrusio, W.; Linguanti, A.; De Santis, W.; Canitano, N.; Ippoliti, F.; Marigliano, V.; Cacciafesta, M. Montelukast therapy and psychological distress in chronic obstructive pulmonary disease (COPD): A preliminary report. Arch. Gerontol. Geriatr. 2011, 52, e36–e39. [Google Scholar] [CrossRef] [PubMed]
- Shimbori, C.; Shiota, N.; Okunishi, H. Pranlukast, a cysteinyl leukotriene type 1 receptor antagonist, attenuates the progression but not the onset of silica-induced pulmonary fibrosis in mice. Int. Arch. Allergy Immunol. 2012, 158, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Ramires, R.; Caiaffa, M.F.; Tursi, A.; Haeggstrom, J.Z.; Macchia, L. Novel inhibitory effect on 5-lipoxygenase activity by the anti-asthma drug montelukast. Biochem. Biophys. Res. Commun. 2004, 324, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Levine, S.J. Bronchial epithelial cell-cytokine interactions in airway inflammation. J. Investig. Med. 1995, 43, 241–249. [Google Scholar] [PubMed]
- Owen, W.F., Jr.; Rothenberg, M.E.; Silberstein, D.S.; Gasson, J.C.; Stevens, R.L.; Austen, K.F.; Soberman, R.J. Regulation of human eosinophil viability, density, and function by granulocyte/macrophage colony-stimulating factor in the presence of 3T3 fibroblasts. J. Exp. Med. 1987, 166, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Resnick, M.B.; Weller, P.F. Mechanisms of eosinophil recruitment. Am. J. Respir. Cell Mol. Biol. 1993, 8, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Zaady, E.; Offer, Z.Y.; Shachak, M. The content and contributions of deposited aeolian organic matter in a dry land ecosystem of the Negev Desert, Israel. Atmos. Environ. 2001, 35, 769–776. [Google Scholar] [CrossRef]
- Hioki, T.; Nakanishi, S.; Mukai, H.; Murano, K.; Ohara, T.; Wakamatsu, S. Analysis of long-range transported and local air pollution with trace metal concentration ratio and lead isotope ratio in precipitation. J. Jpn. Soc. Atmos. Environ. 2008, 43, 100–111. [Google Scholar]
- Wuebbles, D.J.; Lei, H.; Lin, J. Intercontinental transport of aerosols and photochemical oxidants from Asia and its consequences. Environ. Pollut. 2007, 150, 65–84. [Google Scholar] [CrossRef] [PubMed]








© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurai, J.; Watanabe, M.; Sano, H.; Hantan, D.; Tohda, Y.; Shimizu, E. Effects of Asian Dust Particles on the Early-Stage Antigen-Induced Immune Response of Asthma in NC/Nga Mice. Int. J. Environ. Res. Public Health 2016, 13, 1144. https://doi.org/10.3390/ijerph13111144
Kurai J, Watanabe M, Sano H, Hantan D, Tohda Y, Shimizu E. Effects of Asian Dust Particles on the Early-Stage Antigen-Induced Immune Response of Asthma in NC/Nga Mice. International Journal of Environmental Research and Public Health. 2016; 13(11):1144. https://doi.org/10.3390/ijerph13111144
Chicago/Turabian StyleKurai, Jun, Masanari Watanabe, Hiroyuki Sano, Degejirihu Hantan, Yuji Tohda, and Eiji Shimizu. 2016. "Effects of Asian Dust Particles on the Early-Stage Antigen-Induced Immune Response of Asthma in NC/Nga Mice" International Journal of Environmental Research and Public Health 13, no. 11: 1144. https://doi.org/10.3390/ijerph13111144
APA StyleKurai, J., Watanabe, M., Sano, H., Hantan, D., Tohda, Y., & Shimizu, E. (2016). Effects of Asian Dust Particles on the Early-Stage Antigen-Induced Immune Response of Asthma in NC/Nga Mice. International Journal of Environmental Research and Public Health, 13(11), 1144. https://doi.org/10.3390/ijerph13111144