
Structure, Spatial and Temporal Distribution of the Culex pipiens Complex in Shanghai, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Locations and Characteristics of Study Areas
2.2. Study Design
2.3. Identification and Classification of Captured Mosquitoes
2.4. Statistical Analysis
3. Results
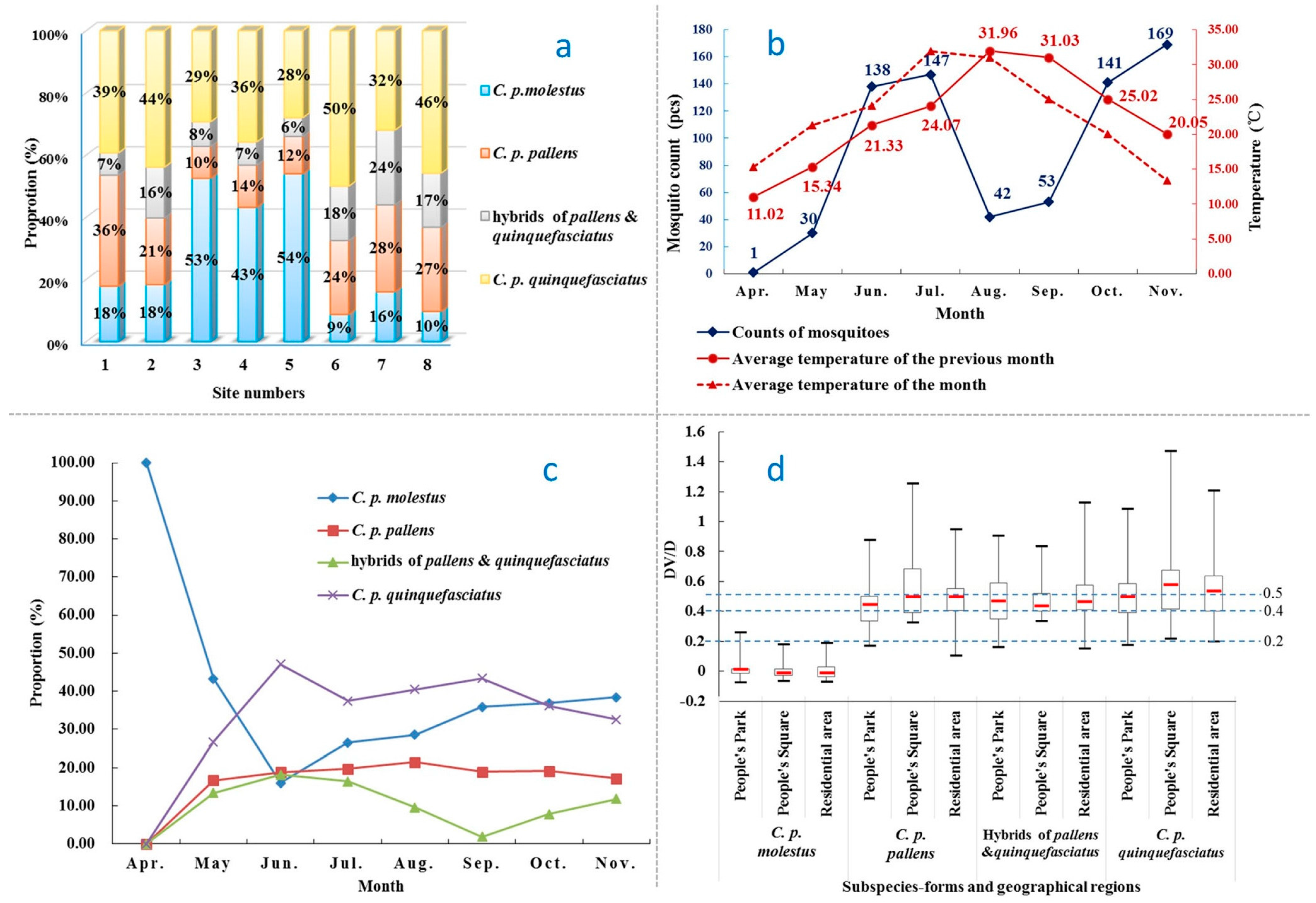
3.1. Population Structure
3.2. Seasonal Fluctuation
3.3. DV/D Ratio
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vinogradova, E.B. Culex pipiens Pipiens Mosquitoes: Taxonomy, Distribution, Ecology, Physiology, Genetics, Applied Importance and Control; Pensoft: Sofia, Bulgaria, 2000; pp. 1–25. [Google Scholar]
- Harbach, R.E. Culex pipiens: Species versus species complex taxonomic history and perspective. J. Am. Mosq. Control Assoc. 2012, 28, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Sirivanakam, S. Medical entomology studies-III. A revision of the subgenus Culex in the Oriental Region (Diptera: Culicidae). Contrib. Am. Entomol. Inst. (Ann Arbor) 1976, 12, 1–272. [Google Scholar]
- Zhao, T.; Lu, B. Biosystematics of Culex pipiens complex in China. Insect Sci. 1995, 2, 1–8. [Google Scholar] [CrossRef]
- Lu, B. Fauna Sinica. Insect Vol. 8, Diptera: Culicidae 1; Science Press: Beijing, China, 1997; pp. 5–410. [Google Scholar]
- Zhao, T.; Lu, B. A new record of Culex pipiens molestus in China and Studies of autogeny and systematics. Chin. J. Vector Biol. Control 1993, 4, 241–243. [Google Scholar]
- Song, S.; Zhao, T.; Dong, Y.; Lu, B. Hybridization and Wolbachia infection in the Culex pipiens complex in China. Acta Entomol. Sin. 2001, 45, 706–710. [Google Scholar]
- Xing, D.; Zhao, T. A new record of Culex pipiens mosquito molestus in Inner Mongolia and its infection with Wolbachia. Acta Parasitol. Med. Entomol. Sin. 2013, 20, 251–254. [Google Scholar]
- Wu, T.P.; Hu, Q.; Zhao, T.; Tian, J.H.; Xue, R.D. Morphological studies on Culex molestus of the Culex pipiens complex (Diptera: Culicidae) in underground parking lots in Wuhan, central China. Fla. Entomol. 2014, 97, 1191–1198. [Google Scholar] [CrossRef]
- Ji, S.; Zhao, T.; Leng, P.; Huang, H.; Mu, D.; Wang, J. A new record of Culex pipiens molestus in Shanghai and studies of autogeny and resistance to four insecticides. Acta Parasitol. Med. Entomol. Sin. 2010, 17, 170–173. [Google Scholar]
- Reisen, W.K.; Milby, M.M.; Presser, S.B.; Hardy, J.L. Ecology of mosquitoes and St. Louis encephalitis virus in the Los Angeles Basin of California, 1987–1990. J. Med. Entomol. 1992, 29, 582–598. [Google Scholar] [PubMed]
- Bogh, C.; Pedersen, E.M.; Mukoko, D.A.; Ouma, J.H. Permethrin-impregnated bednet effects on resting and feeding behaviour of lymphatic filariasis vector mosquitoes in Kenya. Med. Vet. Entomol. 1998, 12, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; Sardelis, M.R.; O’Guinn, M.L.; Dohm, D.J. Potential vectors of West Nile virus in North America. Curr. Top. Microbiol. Immunol. 2002, 267, 241–252. [Google Scholar] [PubMed]
- Kimura, M.; Darbro, J.M.; Harrington, L.C. Avian malaria parasites share congeneric mosquito vectors. J. Parasitol. 2010, 96, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Hayes, E.B.; Komar, N.; Nasci, R.S.; Montgomery, S.P.; O’Leary, D.R.; Campbell, G.L. Epidemiology and transmission dynamics of West Nile virus disease. Emerg. Infect. Dis. 2005, 11, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Hubalek, Z. Mosquito-borne viruses in Europe. Parasitol. Res. 2008, 103, S29–S43. [Google Scholar] [CrossRef] [PubMed]
- Zeller, H.; Lenglet, A.; Van Bortel, W. West Nile virus: The need to strengthen preparedness in Europe. Eurosurveillance 2010, 15, 1–2. [Google Scholar]
- Andreadis, T.G. The contribution of Culex pipiens complex mosquitoes to transmission and persistence of West Nile virus in North America. J. Am. Mosq. Control Assoc. 2012, 28, S137–S151. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, M.; Czajka, C.; Borstler, J.; Melaun, C.; Jost, H.; von Thien, H.; Badusche, M.; Becker, N.; Schmidt-Chanasit, J.; Kruger, A.; et al. First nationwide surveillance of Culex pipiens complex and Culex torrentium mosquitoes demonstrated the presence of Culex pipiens biotype pipiens/molestus hybrids in Germany. PLoS ONE 2013, 8, e71832. [Google Scholar] [CrossRef] [PubMed]
- Barr, A.R. The distribution of Culex p. pipiens and C. p. quinquefasciatus in North America. Am. J. Trop. Med. Hyg. 1957, 6, 153–165. [Google Scholar]
- Van Dine, D.L. A sugarcane leafhopper in Hawaii. Hawaii Agric. Exp. Stn. Bull. 1904, 5, 29. [Google Scholar]
- Spielman, A. Structure and seasonality of nearctic Culex pipiens populations. Ann. N. Y. Acad. Sci. 2001, 951, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Byrne, K.; Nichols, R.A. Culex pipiens in London underground tunnels: Differentiation between surface and subterranean populations. Heredity 1999, 82, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Bahnck, C.M.; Fonseca, D.M. Rapid assay to identify the two genetic forms of Culex (Culex) pipiens L. (Diptera: Culicidae) and hybrid populations. Am. J. Trop. Med. Hyg. 2006, 75, 251–255. [Google Scholar] [PubMed]
- Sundararaman, S. Biometrical studies on intergradation in the genitaiia of certain populations of Culex pipiens and Culex quinquefasciatus in the United States. Am. J. Hyg. 1949, 60, 307–314. [Google Scholar]
- Jakob, W.L.; Francy, D.B. Observations on the DV/D ratio of male genitalia of Culex pipiens complex mosquitoes in the United States. Mosq. Syst. 1984, 16, 282–288. [Google Scholar]
- Iltis, G.W. Biosystematics of the Culex pipiens Complex in Northern California. Ph.D. Thesis, University of California, Davis, CA, USA, 1966. [Google Scholar]
- Jakob, W.L.; Daggers, S.A.; Francy, D.B.; Mullenix, J.; Moseley, K. The Culex pipiens complex in Memphis, Tennessee. Mosq. Syst. 1979, 11, 179–186. [Google Scholar]
- Chen, H.; Lu, B. Study on the Culex pipiens complex in China. J. Guiyang Med. Univ. 1983, 8, 1–13. [Google Scholar]
- Ishii, T. On the Culex pipiens group in Japan. III. A historical review of its research. IV. Review of the adult character (3). J. Sci. (Coll. Gen. Educ. Univ. Tokushima) 1980, 13, 26–62. [Google Scholar]
- Ishii, T. Integrated study onthe Culex pipiens complex: Species diversion in the Culex pipiens complex. Akaieka Newsl. 1991, 14, 5–40. [Google Scholar]
- Zhao, T.; Lu, B. Biosystematic studies on the Culex pipiens complex of China: Mathematical analysis of DV/D ratio of male genitalia. Acta Entomol. Sin. 1994, 37, 446–449. [Google Scholar]
- Rubio, A.; Bellocq, M.I.; Vezzani, D. Macro- and microenvironmental factors affecting tyre-breeding flies (Insecta: Diptera) in urbanised areas. Ecol. Entomol. 2013, 38, 303–314. [Google Scholar] [CrossRef]
- Hansen, A.J.; Knight, R.L.; Marlzuff, J.M.; Powell, S.; Brown, K.; Gude, P.H. Effects of exurban development on biodiversity: Patterns, mechanisms, and research needs. Ecol. Appl. 2005, 15, 1893–1905. [Google Scholar] [CrossRef]
- Paul, M.J.; Meyer, J.L. Streams in the urban landscape. Annu. Rev. Ecol. Syst. 2001, 32, 333–365. [Google Scholar] [CrossRef]
- Rey, J.R.; Nishimura, N.; Wagner, B.; Braks, M.A.; O’Connell, S.M.; Lounibos, L.P. Habitat segregation of mosquito arbovirus vectors in south Florida. J. Med. Entomol. 2006, 43, 1134–1141. [Google Scholar] [CrossRef] [PubMed]
- Wild, J.D.E. Photoperiodism in insects and mites. Annu. Rev. Enterp. 1962, 7, 1–26. [Google Scholar] [CrossRef]





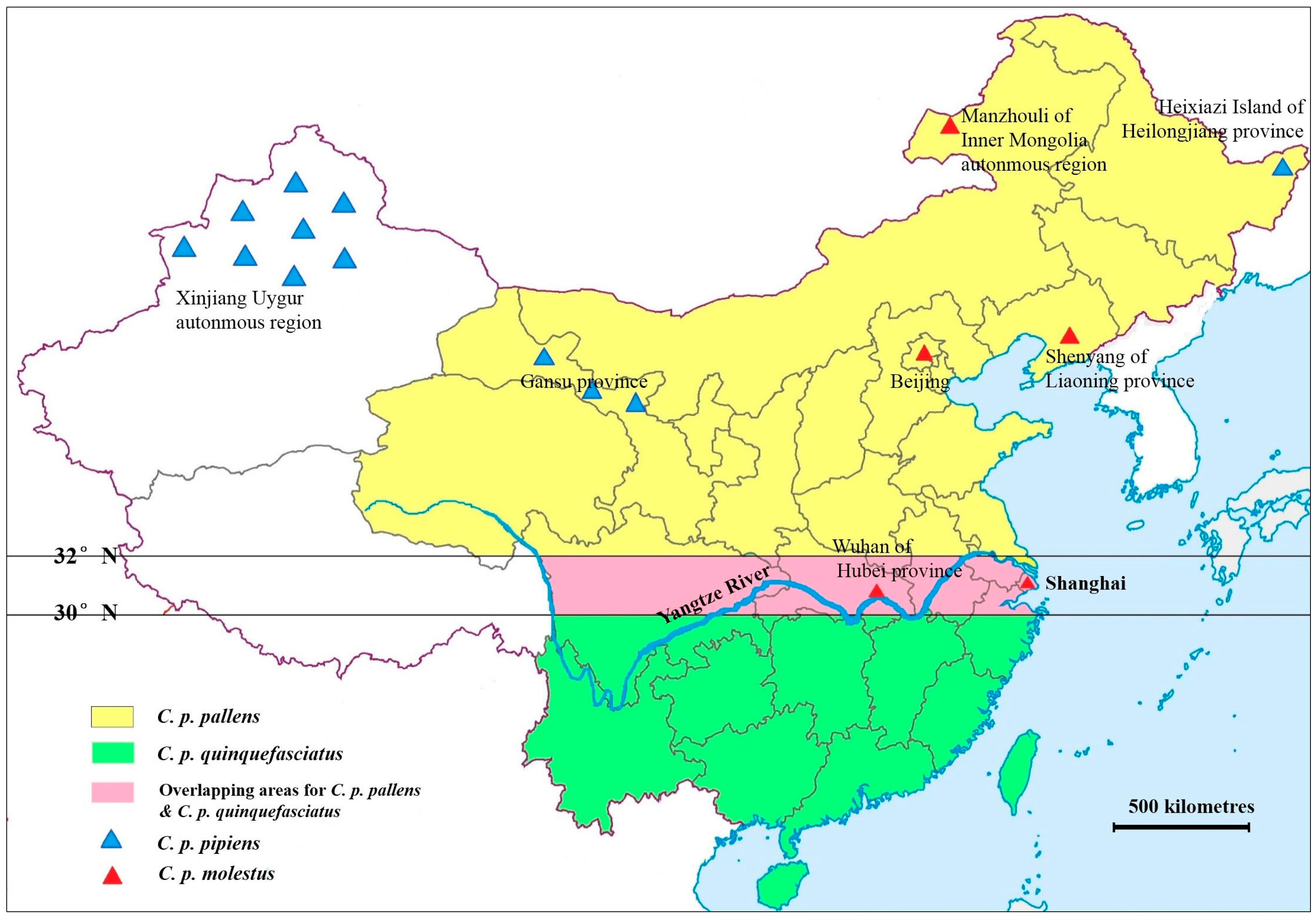{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Numbers | Area | Environmental Types | Coordinates |
|---|---|---|---|
| 1 | People’s Park | Green land (Park) | 31°13′59.72″ N 121°28′02.06″ E |
| 2 | People’s Park | Green land (Park) | 31°14′03.10″ N 121°28′06.26″ E |
| 3 | People’s Park | Green land (Park) | 31°14′02.48″ N 121°28′13.02″ E |
| 4 | People’s Square | Green land | 31°13′53.01″ N 121°28′19.12″ E |
| 5 | People’s Square | Green land | 31°13′47.13″ N 121°28′09.01″ E |
| 6 | Residential area | Old types “Shikumen” | 31°14′11.15″ N 121°27′48.74″ E |
| 7 | Residential area | High-rise residential | 31°14′11.18″ N 121°27′45.45″ E |
| 8 | Residential area | Old Villa | 31°14′15.83″ N 121°27′42.76″ E |
| Site Numbers | C. p. molestus | C. p. pallens | Hybrids of pallens & quinquefasciatus | C. p. quinquefasciatus | Sum |
|---|---|---|---|---|---|
| 1 | 5 (17.86) | 10 (35.71) | 2 (7.14) | 11 (39.29) | 28 |
| 2 | 18 (18.37) | 21 (21.43) | 16 (16.33) | 43 (43.88) | 98 |
| 3 | 113 (52.56) | 22 (10.23) | 17 (7.91) | 63 (29.30) | 215 |
| 4 | 35 (43.21) | 11 (13.58) | 6 (7.41) | 29 (35.80) | 81 |
| 5 | 27 (54.00) | 6 (12.00) | 3 (6.00) | 14 (28.00) | 50 |
| 6 | 7 (8.75) | 19 (23.75) | 14 (17.50) | 40 (50.00) | 80 |
| 7 | 4 (16.00) | 7 (28.00) | 6 (24.00) | 8 (32.00) | 25 |
| 8 | 14 (9.72) | 39 (27.08) | 25 (17.36) | 66 (45.83) | 144 |
| People’s Park (1–3) | 136 (39.88) | 53 (15.54) | 35 (10.26) | 117 (34.31) | 341 |
| People’s Square (4, 5) | 62 (47.33) | 17 (12.98) | 9 (6.87) | 43 (32.82) | 131 |
| Residential (6–8) | 25 (10.04) | 65 (26.10) | 45 (18.07) | 114 (45.78) | 249 |
| Total | 223 (30.93) | 135 (18.72) | 89 (12.34) | 274 (38.00) | 721 |
| Forms | n | Median | Mean ± STD | Intervals |
|---|---|---|---|---|
| C. p. molestus | 223 | −0.012 a | 0.004 ± 0.048 | −0.079–0.258 |
| C. p. pallens | 135 | 0.479 b | 0.486 ± 0.168 | 0.100–1.250 |
| Hybrids of pallens and quinquefasciatus | 89 | 0.469 b,c | 0.490 ± 0.180 | 0.150–1.125 |
| C. p. quinquefasciatus | 274 | 0.500 c | 0.539 ± 0.195 | 0.171–1.467 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Q.; Xiong, C.; Su, F.; Cao, H.; Zhou, J.; Jiang, Q. Structure, Spatial and Temporal Distribution of the Culex pipiens Complex in Shanghai, China. Int. J. Environ. Res. Public Health 2016, 13, 1150. https://doi.org/10.3390/ijerph13111150
Gao Q, Xiong C, Su F, Cao H, Zhou J, Jiang Q. Structure, Spatial and Temporal Distribution of the Culex pipiens Complex in Shanghai, China. International Journal of Environmental Research and Public Health. 2016; 13(11):1150. https://doi.org/10.3390/ijerph13111150
Chicago/Turabian StyleGao, Qiang, Chenglong Xiong, Fei Su, Hui Cao, Jianjun Zhou, and Qingwu Jiang. 2016. "Structure, Spatial and Temporal Distribution of the Culex pipiens Complex in Shanghai, China" International Journal of Environmental Research and Public Health 13, no. 11: 1150. https://doi.org/10.3390/ijerph13111150
APA StyleGao, Q., Xiong, C., Su, F., Cao, H., Zhou, J., & Jiang, Q. (2016). Structure, Spatial and Temporal Distribution of the Culex pipiens Complex in Shanghai, China. International Journal of Environmental Research and Public Health, 13(11), 1150. https://doi.org/10.3390/ijerph13111150