
Reduction of Endogenous Melatonin Accelerates Cognitive Decline in Mice in a Simulated Occupational Formaldehyde Exposure Environment


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
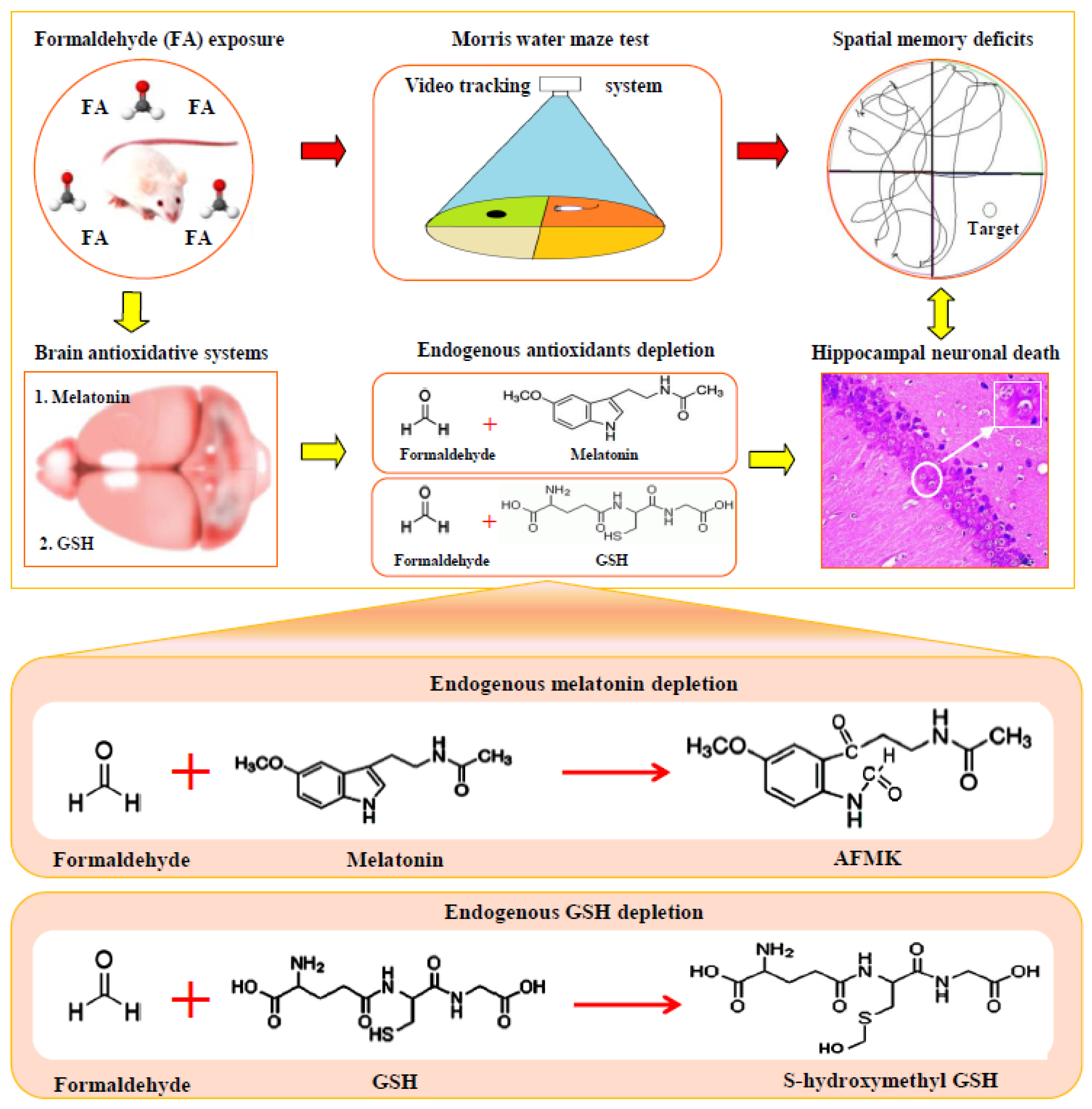
:1. Introduction
2. Methods
2.1. Animals
2.2. Reagents and Kits
2.3. Experimental Protocol
2.4. Morris Water Maze Test
2.5. Preparation of Brain Tissue Homogenates and Histological Analysis
2.6. Determination of FA by UV-HPLC
2.7. Detection of MT Levels Using ELISA Kit
2.8. Intracerebroventricular Injection of Reagents in Mice
2.9. Reactive Oxygen Species (ROS) Assay
2.10. l-glutathione (GSH) Assay
2.11. Cell Culture and Cell Viability Assay
2.12. Chemical Reaction between FA and MT
2.13. Statistical Analysis
3. Results
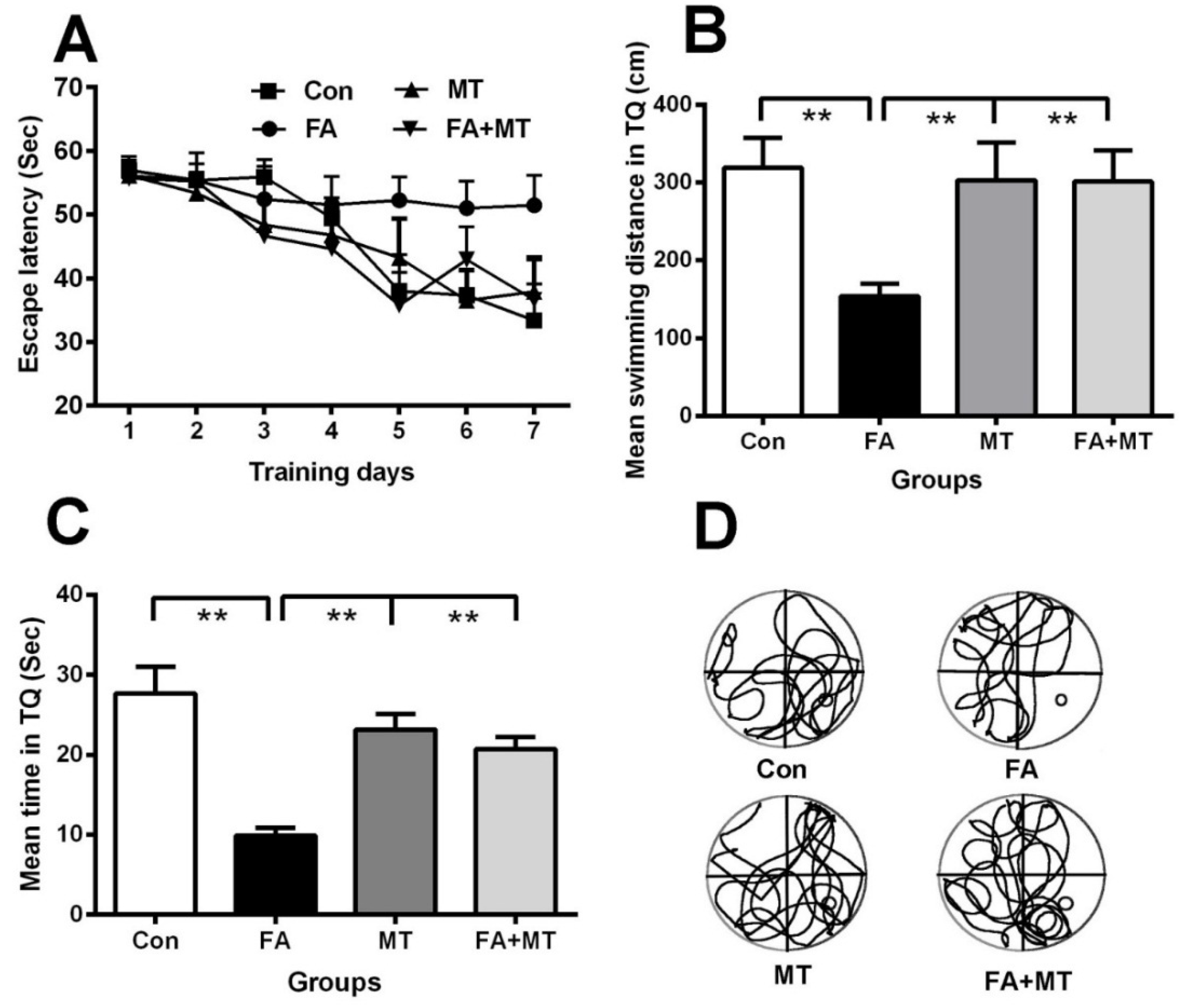
3.1. Treatment with MT Reverses Gaseous FA Exposure-Induced Memory Deficits
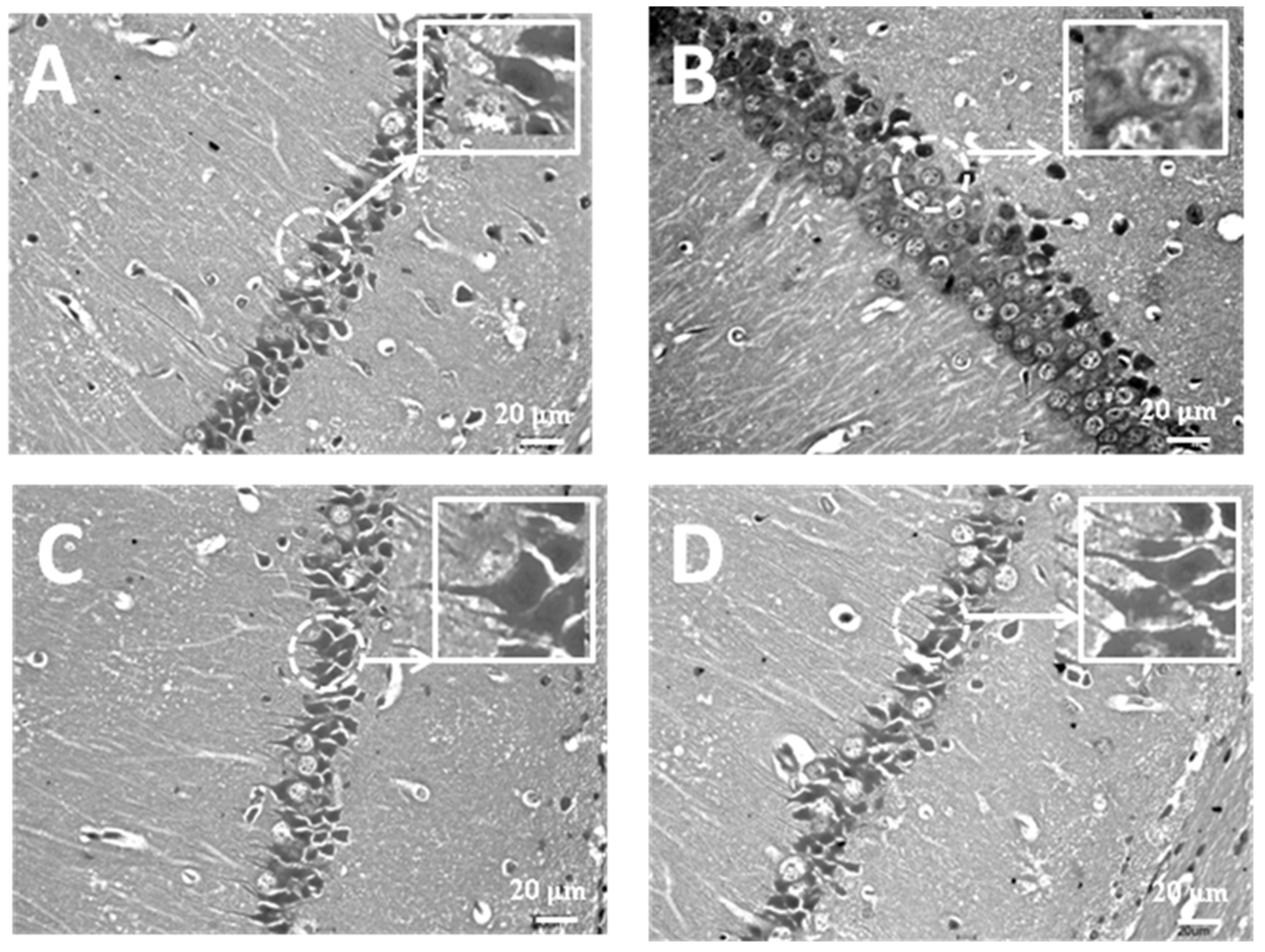
3.2. Treatment with MT Attenuates Hippocampal Neuronal Damage Induced by Exposure to FA
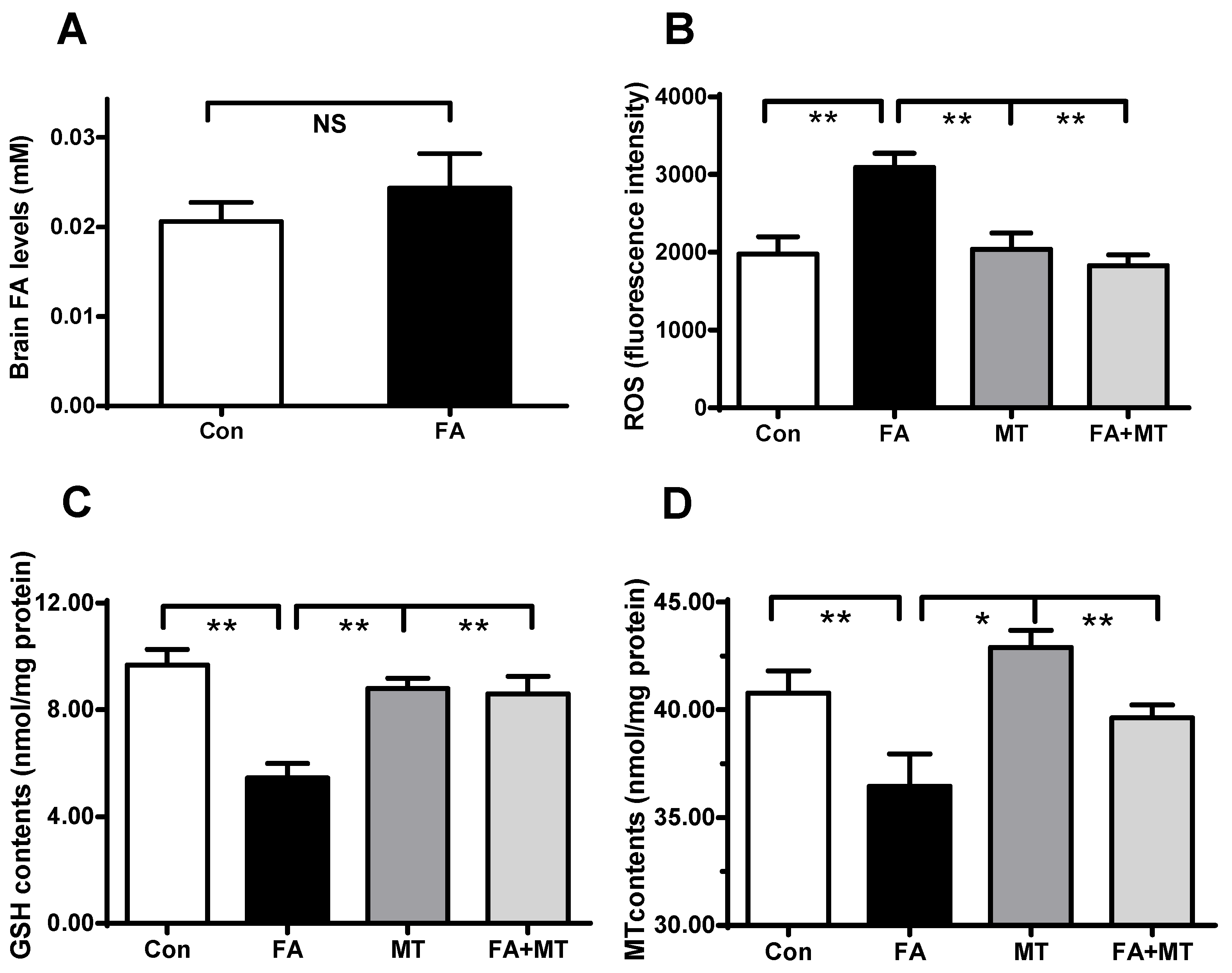
3.3. Application of MT Restores Brain MT Concentrations in Gaseous FA-Exposed Mice
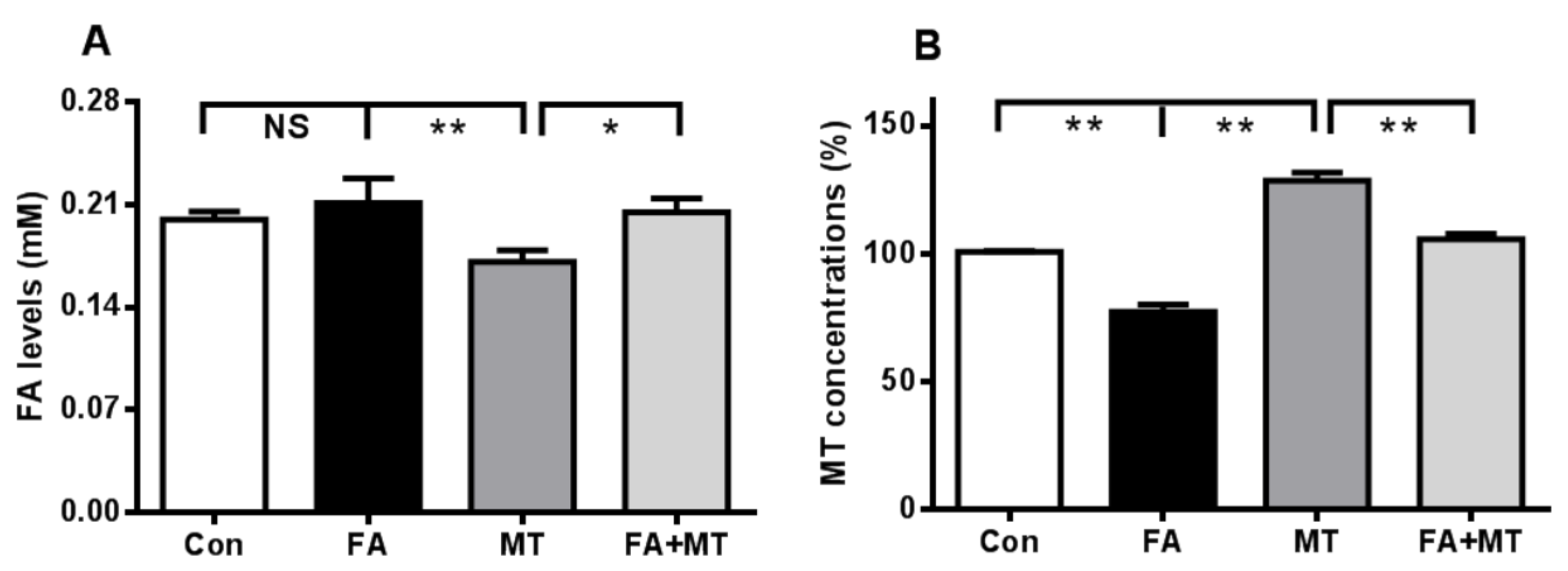
3.4. An i.c.v. Injection of Liquid FA Induces Endogenous MT Reduction in the Brain
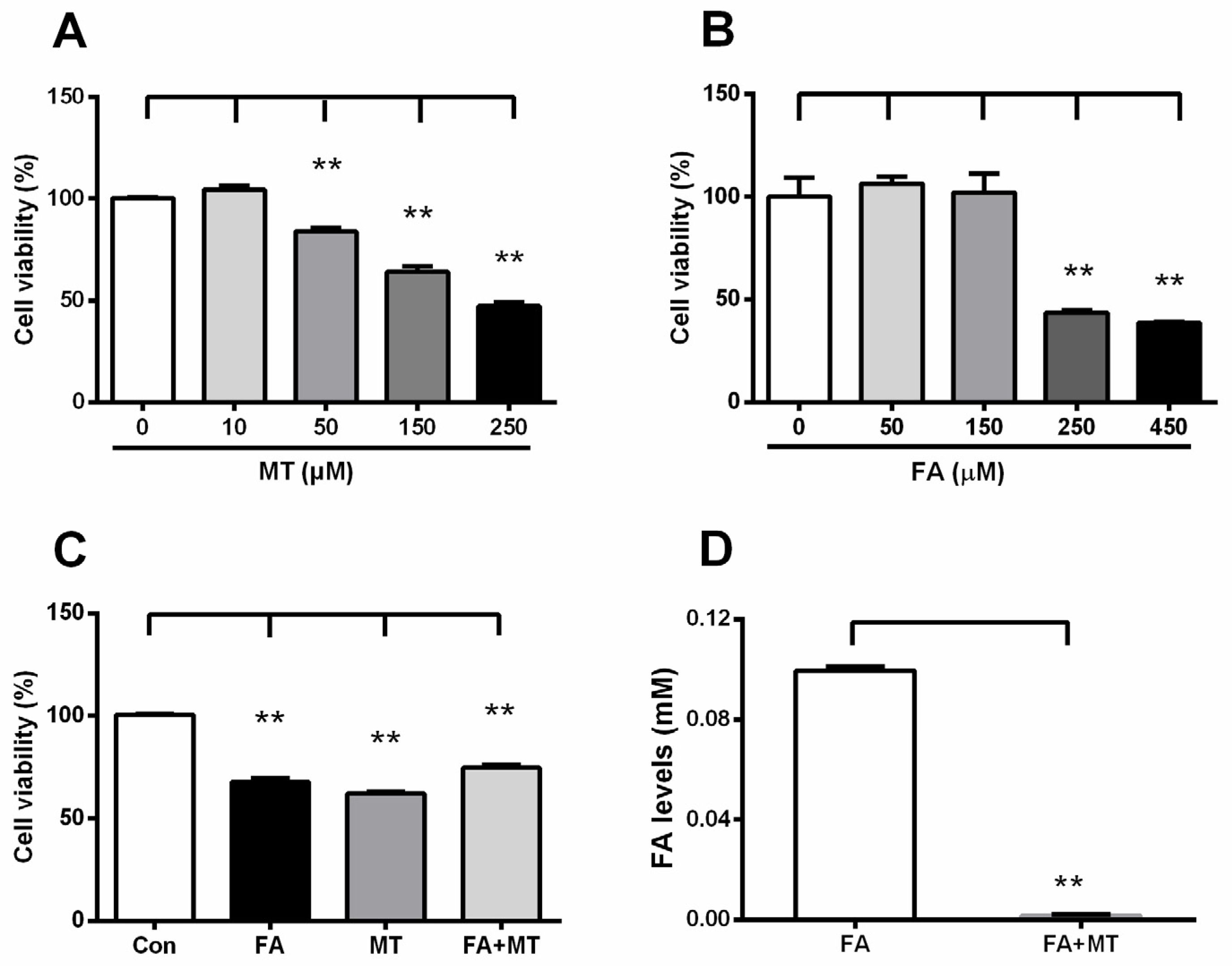
3.5. Liquid FA Directly Inactivates MT in Cultured Cells and in Vitro
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Arts, J.H.; Muijser, H.; Kuper, C.F.; Woutersen, R.A. Setting an indoor air exposure limit for formaldehyde: Factors of concern. Regul. Toxicol. Pharmacol. 2008, 52, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Songur, A.; Ozen, O.A.; Sarsilmaz, M. The toxic effects of formaldehyde on the nervous system. Rev. Environ. Contam Toxicol. 2010, 203, 105–118. [Google Scholar] [PubMed]
- Bohm, M.; Salem, M.Z.; Srba, J. Formaldehyde emission monitoring from a variety of solid wood, plywood, blockboard and flooring products manufactured for building and furnishing materials. J. Hazard. Mater. 2012, 221–222, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Bai, Y.; Duong, A.; Smith, M.T.; Li, L.; Zhang, L. Formaldehyde in China: Production, consumption, exposure levels, and health effects. Environ. Int. 2009, 35, 1210–1224. [Google Scholar] [CrossRef] [PubMed]
- Kilburn, K.H.; Warshaw, R.; Thornton, J.C. Formaldehyde impairs memory, equilibrium, and dexterity in histology technicians: Effects which persist for days after exposure. Arch. Environ. Health 1987, 42, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Kilburn, K.H.; Warshaw, R.H. Neurobehavioral effects of formaldehyde and solvents on histology technicians: Repeated testing across time. Environ. Res. 1992, 58, 134–146. [Google Scholar] [CrossRef]
- Usanmaz, S.E.; Akarsu, E.S.; Vural, N. Neurotoxic effects of acute and subacute formaldehyde exposures in mice. Environ. Toxicol. Pharmacol. 2002, 11, 93–100. [Google Scholar] [CrossRef]
- Liu, Y.; Ye, Z.; Luo, H.; Sun, M.; Li, M.; Fan, D.; Chui, D. Inhalative formaldehyde exposure enhances aggressive behavior and disturbs monoamines in frontal cortex synaptosome of male rats. Neurosci. Lett. 2009, 464, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Li, C.M.; Qiao, Y.; Yan, Y.; Yang, X. Effect of inhaled formaldehyde on learning and memory of mice. Indoor Air 2008, 18, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Conaway, C.C.; Whysner, J.; Verna, L.K.; Williams, G.M. Formaldehyde mechanistic data and risk assessment: Endogenous protection from DNA adduct formation. Pharmacol. Ther. 1996, 71, 29–55. [Google Scholar] [CrossRef]
- Schroeter, J.D.; Campbell, J.; Kimbell, J.S.; Conolly, R.B.; Clewell, H.J.; Andersen, M.E. Effects of endogenous formaldehyde in nasal tissues on inhaled formaldehyde dosimetry predictions in the rat, monkey, and human nasal passages. Toxicol. Sci. 2014, 138, 412–424. [Google Scholar] [CrossRef] [PubMed]
- Heck, H.D.; White, E.L.; Casanova-Schmitz, M. Determination of formaldehyde in biological tissues by gas chromatography/mass spectrometry. Biomed. Mass Spectrom 1982, 9, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R.; Pandi-Perumal, S.R.; Cardinali, D.P. Melatonin. Int. J. Biochem. Cell. Biol. 2006, 38, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, D.; Bjoro, T.; Gautvik, K.; Haug, E. Melatonin reduces the production and secretion of prolactin and growth hormone from rat pituitary cells in culture. Acta Physiol. Scand. 1987, 131, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Menendez-Pelaez, A.; Poeggeler, B.; Reiter, R.J.; Barlow-Walden, L.; Pablos, M.I.; Tan, D.X. Nuclear localization of melatonin in different mammalian tissues: Immunocytochemical and radioimmunoassay evidence. J. Cell. Biochem. 1993, 53, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Altun, A.; Ugur-Altun, B. Melatonin: Therapeutic and clinical utilization. Int. J. Clin. Pract. 2007, 61, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Rodenbeck, A.; Huether, G.; Ruther, E.; Hajak, G. Altered circadian melatonin secretion patterns in relation to sleep in patients with chronic sleep-wake rhythm disorders. J. Pineal Res. 1998, 25, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Ali, T.; Kim, M.O. Melatonin ameliorates amyloid beta-induced memory deficits, tau hyperphosphorylation and neurodegeneration via PI3/Akt/GSk3beta pathway in the mouse hippocampus. J. Pineal Res. 2015, 59, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Maurizi, C.P. The therapeutic potential for tryptophan and melatonin: Possible roles in depression, sleep, Alzheimer’s disease and abnormal aging. Med. Hypotheses 1990, 31, 233–242. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Vigo, D.E.; Olivar, N.; Vidal, M.F.; Furio, A.M.; Brusco, L.I. Therapeutic application of melatonin in mild cognitive impairment. Am.J. Neurodegener. Dis. 2012, 1, 280–291. [Google Scholar] [PubMed]
- Li, Y.; Song, Z.; Ding, Y.; Xin, Y.; Wu, T.; Su, T.; He, R.; Tai, F.; Lian, Z. Effects of formaldehyde exposure on anxiety-like and depression-like behavior, cognition, central levels of glucocorticoid receptor and tyrosine hydroxylase inmice. Chemosphere 2016, 144, 2004–2012. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.; Jiang, C.; Wan, Y.; Lv, J.; Jia, J.; Wang, X.; Yang, X.; Tong, Z. Aging-associated formaldehyde-induced norepinephrine deficiency contributes to age-related memory decline. Aging Cell. 2015, 14, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Han, C.; Luo, W.; Li, H.; Luo, H.; Qiang, M.; Su, T.; Wu, B.; Liu, Y.; Yang, X.; et al. Aging-associated excess formaldehyde leads to spatial memory deficits. Sci. Rep. 2013, 3, 1807. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Corral, S.A.; Acuna-Castroviejo, D.; Coto-Montes, A.; Boga, J.A.; Manchester, L.C.; Fuentes-Broto, L.; Korkmaz, A.; Ma, S.; Tan, D.X.; Reiter, R.J. Alzheimer’s disease: Pathological mechanisms and the beneficial role of melatonin. J. Pineal Res. 2012, 52, 167–202. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.Y.; Zhou, J.; Van Heerikhuize, J.; Hofman, M.A.; Swaab, D.F. Decreased melatonin levels in postmortem cerebrospinal fluid in relation to aging, Alzheimer’s disease, and apolipoprotein E-epsilon4/4 genotype. J. Clin. Endocrinol. Metab. 1999, 84, 323–327. [Google Scholar] [PubMed]
- Sirin, B.K.; Doguc, D.; Vural, H.; Ibrahim, E.; Ikbal, I.; Recep, S.; Namik, D. Plasma 8-isoPGF2alpha and serum melatonin levels in patients with minimal cognitive impairment and Alzheimer disease. Turk. J. Med. Sci. 2015, 45, 1073–1077. [Google Scholar] [CrossRef] [PubMed]
- Zararsiz, I.; Kus, I.; Ogeturk, M.; Akpolat, N.; Kose, E.; Meydan, S.; Sarsilmaz, M. Melatonin prevents formaldehyde-induced neurotoxicity in prefrontal cortex of rats: An immunohistochemical and biochemical study. Cell. Biochem. Funct. 2007, 25, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Ozen, O.A.; Kus, M.A.; Kus, I.; Alkoc, O.A.; Songur, A. Protective effects of melatonin against formaldehyde-induced oxidative damage and apoptosis in rat testes: An immunohistochemical and biochemical study. Syst. Biol. Reprod. Med. 2008, 54, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Pacchierotti, C.; Bossini, L.; Pieraccini, F.; Castrogiovanni, P. Melatonin in psychiatric disorders: A review on the melatonin involvement in psychiatry. Front. Neuroendocrinol. 2001, 22, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Ito, J.; Saijo, H.; Tanaka, H.; Tasaki, T.; Chou, K. Melatonin treatment for sleep-wake disorder—An experience for a severely mental retarded patient with blindness. Brain Dev. 1995, 27, 401–403. [Google Scholar]
- Olcese, J.M.; Cao, C.; Mori, T.; Mamcarz, M.B.; Maxwell, A.; Runfeldt, M.J.; Wang, L.; Zhang, C.; Lin, X.; Zhang, G.; et al. Protection against cognitive deficits and markers of neurodegeneration by long-term oral administration of melatonin in a transgenic model of Alzheimer disease. J. Pineal Res. 2009, 47, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Zhang, J.; Luo, W.; Wang, W.; Li, F.; Li, H.; Luo, H.; Lu, J.; Zhou, J.; Wan, Y.; et al. Urine formaldehyde level is inversely correlated to mini mental state examination scores in senile dementia. Neurobiol. Aging 2011, 32, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Sun, C.F.; Zhang, J.; Gao, Y.B.; Wang, L.F.; Zuo, H.Y.; Wang, S.M.; Zhou, H.M.; Xu, X.P.; Dong, J.; et al. Microwave exposure impairs synaptic plasticity in the rat hippocampus and PC12 cells through over-activation of the NMDA receptor signaling pathway. Biomed. Environ. Sci. 2015, 28, 13–24. [Google Scholar] [PubMed]
- Cordis, G.A.; Bagchi, D.; Maulik, N.; Das, D.K. High-performance liquid chromatographic method for the simultaneous detection of malonaldehyde, acetaldehyde, formaldehyde, acetone and propionaldehyde to monitor the oxidative stress in heart. J. Chromatogr. A 1994, 661, 181–191. [Google Scholar] [CrossRef]
- Feng, Z.; Qin, C.; Chang, Y.; Zhang, J.T. Early melatonin supplementation alleviates oxidative stress in a transgenic mouse model of Alzheimer’s disease. Free Radic Biol. Med. 2006, 40, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Han, C.; Luo, W.; Wang, X.; Li, H.; Luo, H.; Zhou, J.; Qi, J.; He, R. Accumulated hippocampal formaldehyde induces age-dependent memory decline. Age (Dordr) 2013, 35, 583–596. [Google Scholar] [CrossRef] [PubMed]
- Umur, E.E.; Oktenli, C.; Celik, S.; Tangi, F.; Sayan, O.; Sanisoglu, Y.S.; Ipcioglu, O.; Terekeci, H.M.; Top, C.; Nalbant, S.; et al. Increased iron and oxidative stress are separately related to cognitive decline in elderly. Geriatr. Gerontol. Int. 2011, 11, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Ferreiro, E.; Baldeiras, I.; Ferreira, I.L.; Costa, R.O.; Rego, A.C.; Pereira, C.F.; Oliveira, C.R. Mitochondrial- and endoplasmic reticulum-associated oxidative stress in Alzheimer’s disease: From pathogenesis to biomarkers. Int. J. Cell. Biol. 2012, 2012, 735206. [Google Scholar] [CrossRef] [PubMed]
- Moneim, A.E. Oxidant/Antioxidant imbalance and the risk of Alzheimer’s disease. Curr. Alzheimer Res. 2015, 12, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Nishio, K.; Yoshida, Y.; Niki, E. Cytotoxic effect of formaldehyde with free radicals via increment of cellular reactive oxygen species. Toxicology 2005, 210, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.Q.; Ren, Y.K.; Zhou, C.F.; Yang, C.T.; Gu, H.F.; He, J.Q.; Chen, R.Q.; Zhuang, Y.Y.; Fang, H.R.; Wang, C.Y. Hydrogen sulfide prevents formaldehyde-induced neurotoxicity to PC12 cells by attenuation of mitochondrial dysfunction and pro-apoptotic potential. Neurochem. Int. 2012, 61, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Ji, Z.; Wei, C.; McHale, C.M.; Ding, S.; Thomas, R.; Yang, X.; Zhang, L. Inhaled formaldehyde induces DNA-protein crosslinks and oxidative stress in bone marrow and other distant organs of exposed mice. Environ. Mol. Mutagen. 2013, 54, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Gulec, M.; Gurel, A.; Armutcu, F. Vitamin E protects against oxidative damage caused by formaldehyde in the liver and plasma of rats. Mol. Cell. Biochem. 2006, 290, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Romanazzi, V.; Pirro, V.; Bellisario, V.; Mengozzi, G.; Peluso, M.; Pazzi, M.; Bugiani, M.; Verlato, G.; Bono, R. 15-F(2)t isoprostane as biomarker of oxidative stress induced by tobacco smoke and occupational exposure to formaldehyde in workers of plastic laminates. Sci. Total Environ. 2013, 442, 20–25. [Google Scholar] [CrossRef]
- Chen, Q.; Yu, W.; Shi, J.; Shen, J.; Hu, Y.; Gong, J.; Li, J.; Li, N. The effect of extracorporeal membrane oxygenation therapy on systemic oxidative stress injury in a porcine model. Artif. Organs. 2014, 38, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Currais, A.; Maher, P. Functional consequences of age-dependent changes in glutathione status in the brain. Antioxid. Redox Signal. 2013, 19, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Teng, S.; Beard, K.; Pourahmad, J.; Moridani, M.; Easson, E.; Poon, R.; O’Brien, P.J. The formaldehyde metabolic detoxification enzyme systems and molecular cytotoxic mechanism in isolated rat hepatocytes. Chem. Biol. Interact. 2001, 130–132, 285–296. [Google Scholar] [CrossRef]
- Hardeland, R. Antioxidative protection by melatonin: Multiplicity of mechanisms from radical detoxification to radical avoidance. Endocrine 2005, 27, 119–130. [Google Scholar] [CrossRef]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolin, I.; Herrera, F.; Martin, V.; Reiter, R.J. Regulation of antioxidant enzymes: A significant role for melatonin. J. Pineal Res. 2004, 36, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tulpule, K.; Dringen, R. Formaldehyde in brain: An overlooked player in neurodegeneration? J. Neurochem. 2013, 127, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Buzsaki, G.; Moser, E.I. Memory, navigation and theta rhythm in the hippocampal-entorhinal system. Nat. Neurosci. 2013, 16, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Reiter, R.J.; Qi, W.B.; Karbownik, M.; Calvo, J.R. Significance of melatonin in antioxidative defense system: Reactions and products. Biol. Signals Recept 2000, 9, 137–159. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Neurobiology, pathophysiology, and treatment of melatonin deficiency and dysfunction. Sci. World J. 2012, 2012, 640389. [Google Scholar] [CrossRef] [PubMed]
- Jean-Louis, G.; von Gizycki, H.; Zizi, F. Melatonin effects on sleep, mood, and cognition in elderly with mild cognitive impairment. J. Pineal Res. 1998, 25, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Haimov, I.; Laudon, M.; Zisapel, N.; Souroujon, M.; Nof, D.; Shlitner, A.; Herer, P.; Tzischinsky, O.; Lavie, P. Sleep disorders and melatonin rhythms in elderly people. BMJ 1994, 309, 167. [Google Scholar] [CrossRef] [PubMed]
- Karasek, M. Melatonin, human aging, and age-related diseases. Exp. Gerontol. 2004, 39, 1723–1729. [Google Scholar] [CrossRef] [PubMed]
- Almughrabi, O.M.; Marzouk, K.M.; Hasanato, R.M.; Shafik, S.S. Melatonin levels in periodontal health and disease. J. Periodontal Res. 2013, 48, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Corrales, A.; Martinez, P.; Garcia, S.; Vidal, V.; Garcia, E.; Florez, J.; Sanchez-Barcelo, E.J.; Martinez-Cue, C.; Rueda, N. Long-term oral administration of melatonin improves spatial learning and memory and protects against cholinergic degeneration in middle-aged Ts65Dn mice, a model of Down syndrome. J. Pineal Res. 2013, 54, 346–358. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.H.; Swaab, D.F. The human pineal gland and melatonin in aging and Alzheimer’s disease. J. Pineal Res. 2005, 38, 145–152. [Google Scholar] [CrossRef] [PubMed]







© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mei, Y.; Duan, C.; Li, X.; Zhao, Y.; Cao, F.; Shang, S.; Ding, S.; Yue, X.; Gao, G.; Yang, H.; et al. Reduction of Endogenous Melatonin Accelerates Cognitive Decline in Mice in a Simulated Occupational Formaldehyde Exposure Environment. Int. J. Environ. Res. Public Health 2016, 13, 258. https://doi.org/10.3390/ijerph13030258
Mei Y, Duan C, Li X, Zhao Y, Cao F, Shang S, Ding S, Yue X, Gao G, Yang H, et al. Reduction of Endogenous Melatonin Accelerates Cognitive Decline in Mice in a Simulated Occupational Formaldehyde Exposure Environment. International Journal of Environmental Research and Public Health. 2016; 13(3):258. https://doi.org/10.3390/ijerph13030258
Chicago/Turabian StyleMei, Yufei, Chunli Duan, Xiaoxiao Li, Yun Zhao, Fenghua Cao, Shuai Shang, Shumao Ding, Xiangpei Yue, Ge Gao, Hui Yang, and et al. 2016. "Reduction of Endogenous Melatonin Accelerates Cognitive Decline in Mice in a Simulated Occupational Formaldehyde Exposure Environment" International Journal of Environmental Research and Public Health 13, no. 3: 258. https://doi.org/10.3390/ijerph13030258
APA StyleMei, Y., Duan, C., Li, X., Zhao, Y., Cao, F., Shang, S., Ding, S., Yue, X., Gao, G., Yang, H., Shen, L., Feng, X., Jia, J., Tong, Z., & Yang, X. (2016). Reduction of Endogenous Melatonin Accelerates Cognitive Decline in Mice in a Simulated Occupational Formaldehyde Exposure Environment. International Journal of Environmental Research and Public Health, 13(3), 258. https://doi.org/10.3390/ijerph13030258