
Changes in Physiological and Agronomical Parameters of Barley (Hordeum vulgare) Exposed to Cerium and Titanium Dioxide Nanoparticles

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Experimental Section
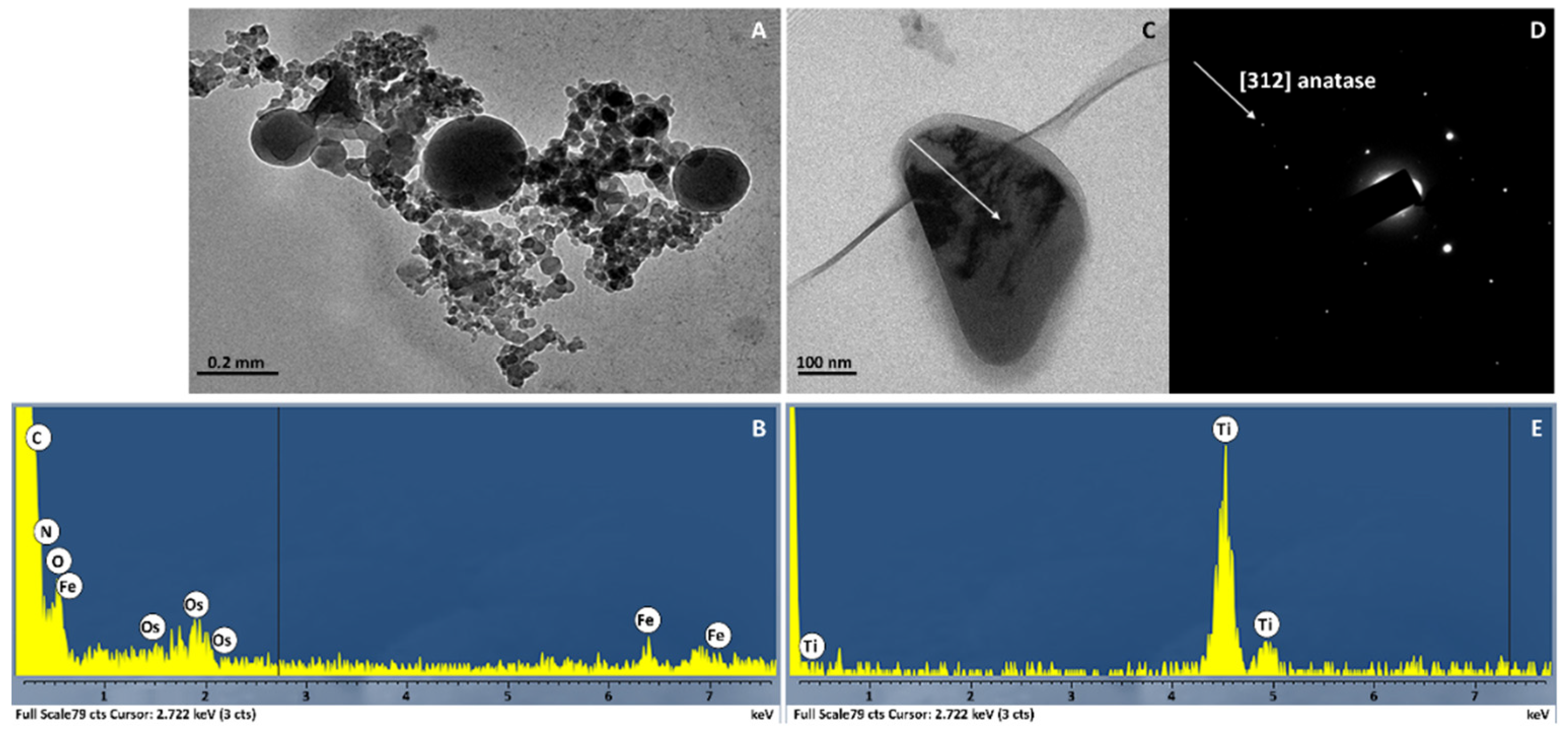
2.1. Characterization of nCeO2 and nTiO2
2.2. Addition of Nanoparticles to Soil
2.3. Plant Growth and Harvest
2.4. Gas Exchange Parameters
2.5. TEM Observations
2.6. Spectroscopy Analysis
2.7. TEM X-ray Microanalysis
2.8. Data Analysis
3. Results
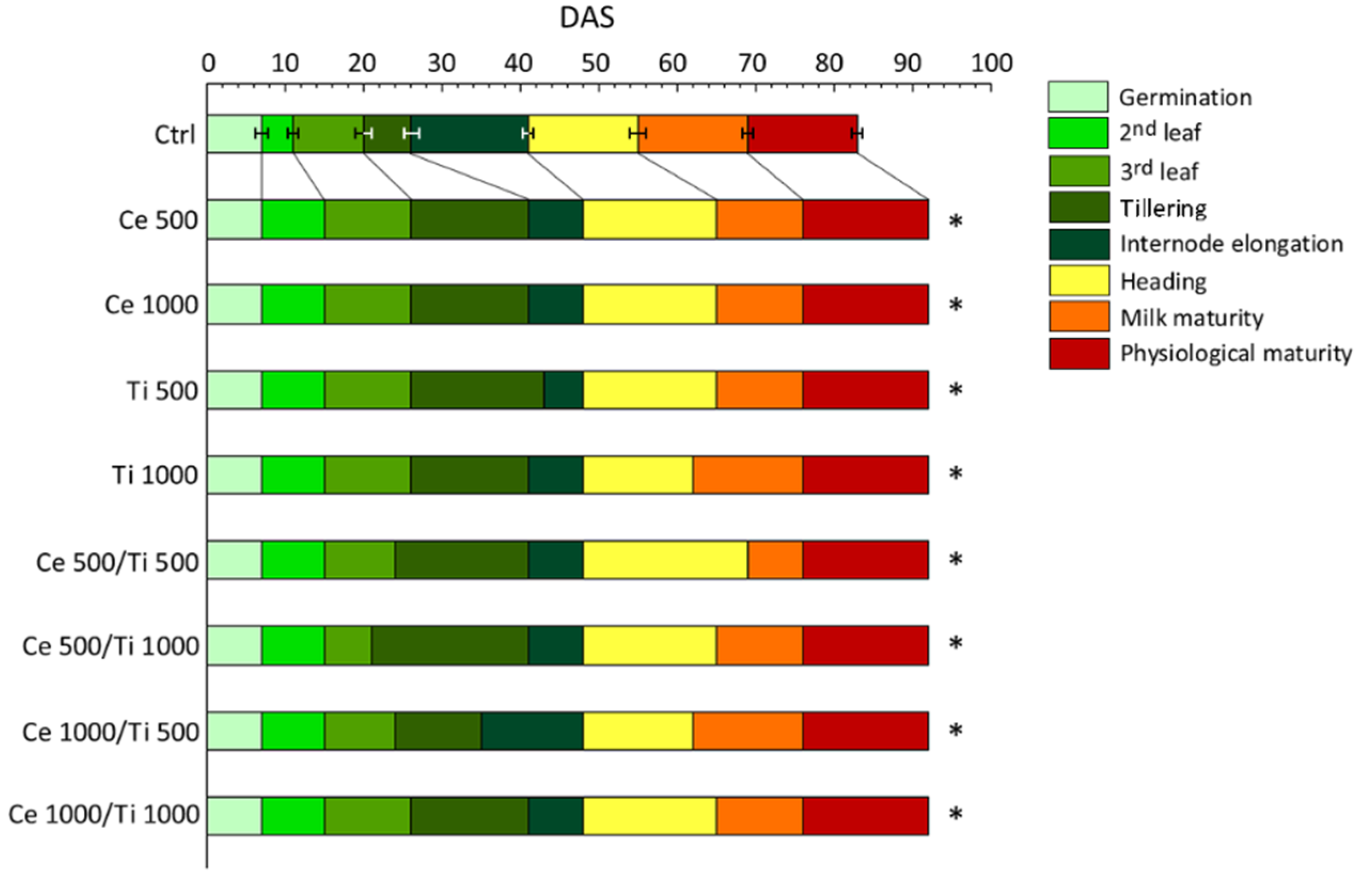
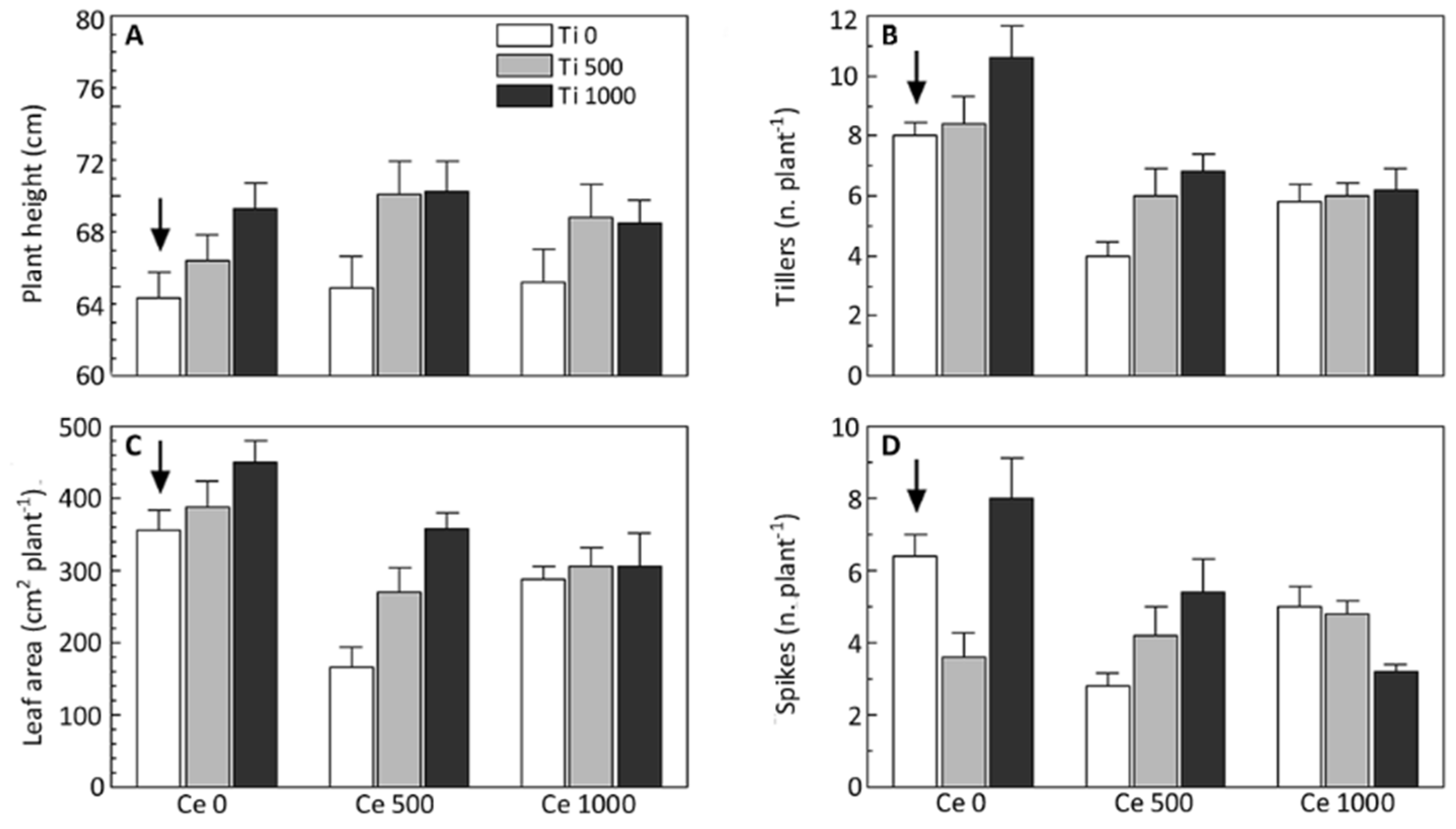
3.1. Phenology and Growth of Barley
3.2. Gas Exchanges
3.3. Plant Uptake and Accumulation of Cerium and Titanium
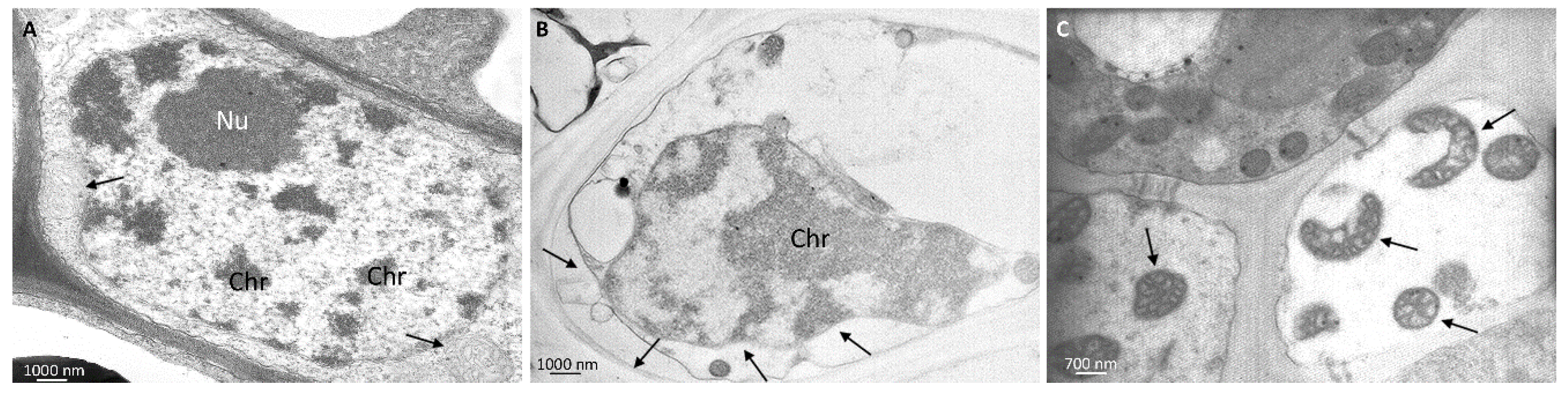
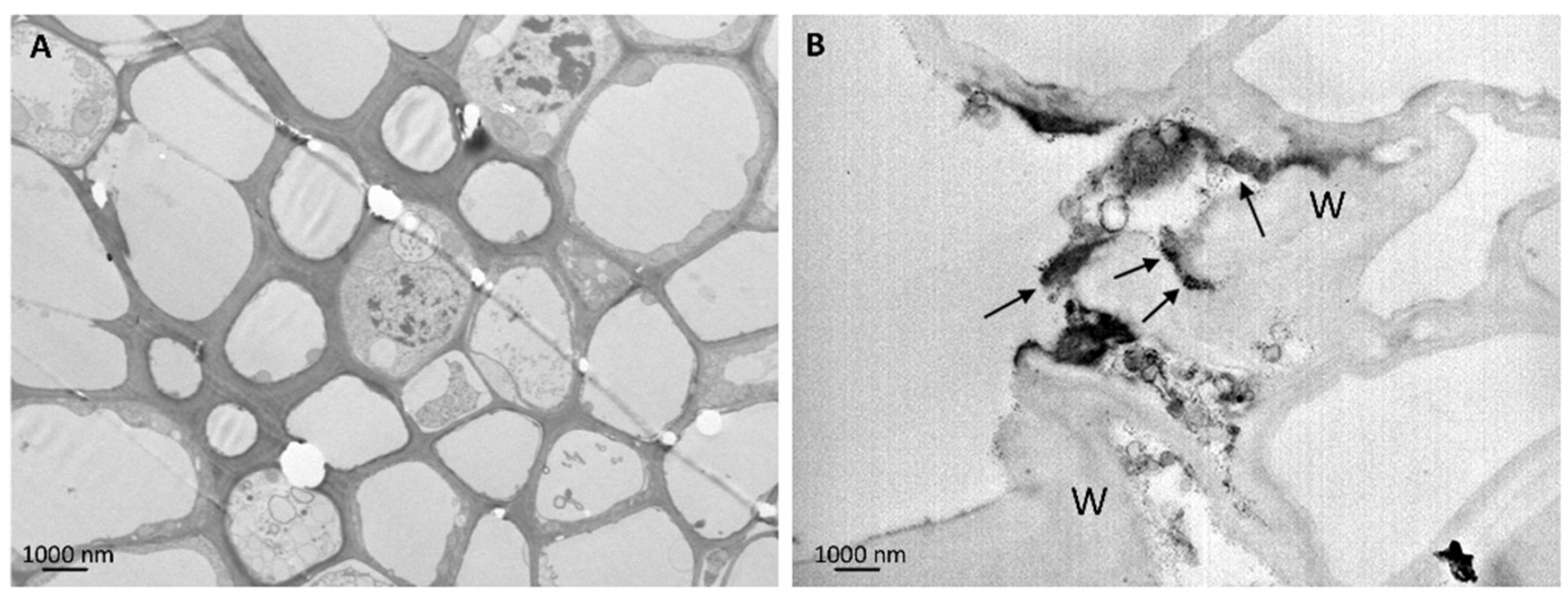
3.4. Ultrastructural Analyses
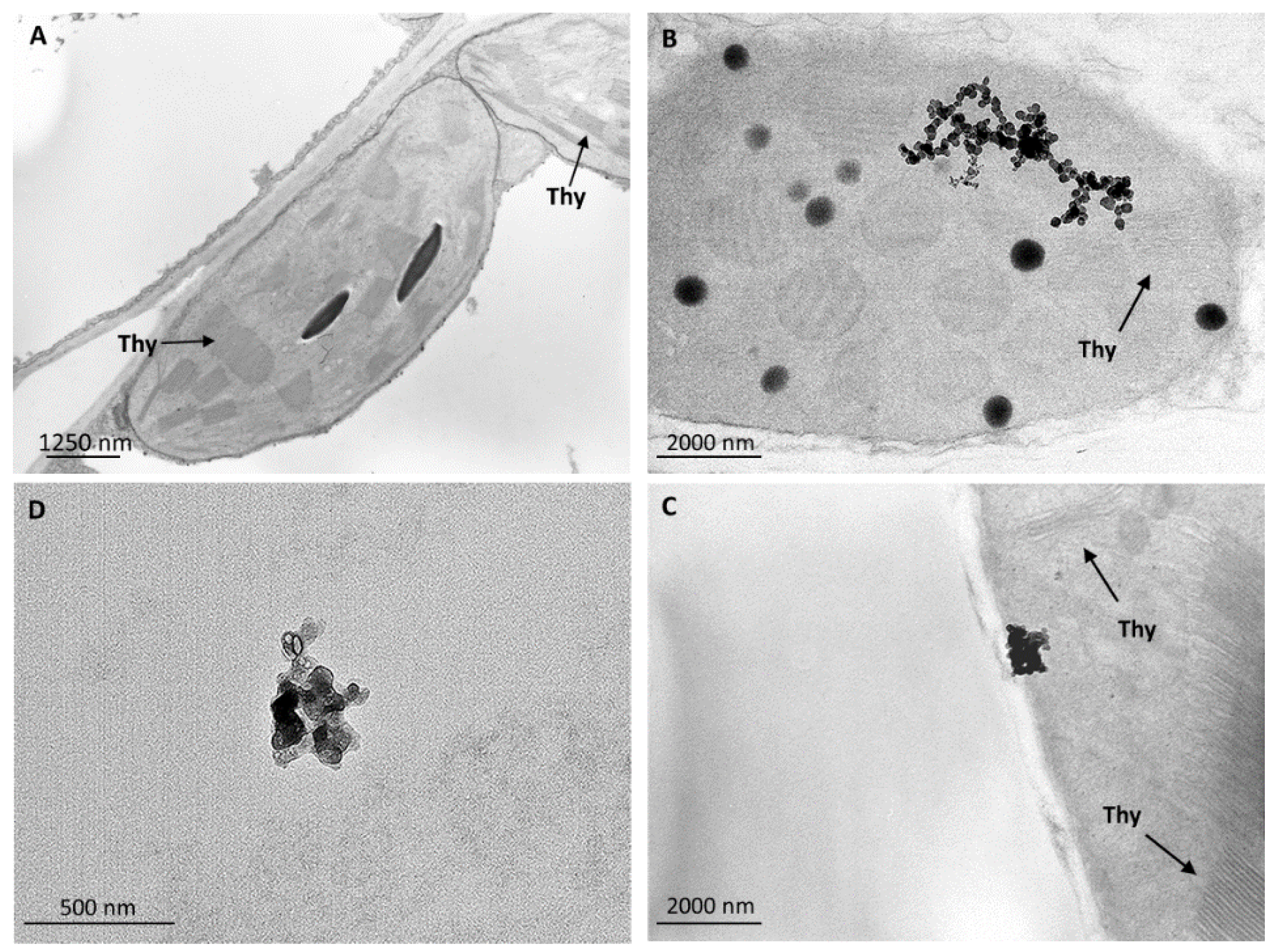
3.5. Nanostructures in Leaf Tissues
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tourinho, P.S.; Van Gestel, C.A.M.; Lofts, S.; Svedsen, C.; Soares, A.M.V.M.; Loureiro, S. Metal-based nanoparticles in soil: Fate, behavior, and effects on soil invertebrates. Environ. Toxicol. Chem. 2012, 31, 1679–1692. [Google Scholar] [CrossRef] [PubMed]
- Batley, G.; Kirby, J.K.; McClaughlin, M.J. Fate and risks of nanomaterials in aquatic and terrestrial environments. Acc. Chem. Res. 2013, 46, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Piccinno, F.; Gottschalk, F.; Seeger, S.; Nowack, B. Industrial production quantities and uses of ten engineered nanomaterials in Europe and the world. J. Nanopart. Res. 2012. [Google Scholar] [CrossRef]
- Gogos, A.; Knauer, K.; Bucheli, T.D. Nanomaterials in plant protection and fertilization: Current state, foreseen applications, and research priorities. J. Agric. Food Chem. 2012, 60, 9781–9792. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.H.; Cohen, Y. Multimedia environmental distribution of engineered nanomaterials. Environ. Sci. Technol. 2014, 48, 3281–3292. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; He, X.; Zhang, H.; Ma, Y.; Zhang, P.; Ding, Y.; Zhao, Y. Uptake and distribution of ceria nanoparticles in cucumber plants. Metallomics 2011, 3, 816–822. [Google Scholar] [CrossRef] [PubMed]
- OECD. List of Manufactured Nanomaterials and List of Endpoints for Phase One of the Sponsorship Programme of Testing on Manufactured Nanomaterials: Revision. OECD Environment, Health and Safety Publications Series on the Safety of Manufactured Nanomaterials, No. 27, Paris, France. 2010. Available online: http://www.oecd.org/officialdocuments/publicdisplaydocumentpdf/?cote=env/jm/mono(2010)46&doclanguage=en (accessed on 30 December 2015).
- Rico, C.M.; Majumdar, S.; Duarte-Gardea, M.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Interaction of nanoparticles with edible plants and their possible implications in the food chain. J. Agric. Food Chem. 2011, 59, 3485–3498. [Google Scholar] [CrossRef] [PubMed]
- Miralles, P.; Church, T.L.; Harris, A.T. Toxicity, uptake, and translocation of engineered nanomaterials in vascular plants. Environ. Sci. Technol. 2012, 46, 9224–9239. [Google Scholar] [CrossRef] [PubMed]
- Gardea-Torresdey, J.L.; Rico, C.M.; White, J.C. Trophic transfer, transformation, and impact of engineered nanomaterials in terrestrial environments. Environ. Sci. Technol. 2014, 48, 2526–2540. [Google Scholar] [CrossRef] [PubMed]
- Capaldi Arruda, S.C.; Silva, A.L.; Galazzi, R.M.; Azevedo, R.A.; Arruda, M.A. Nanoparticles applied to plant science: A review. Talanta 2015, 131, 693–705. [Google Scholar] [CrossRef] [PubMed]
- López-Moreno, M.L.; de La Rosa, G.; Hernández-Viezcas, J.A.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. X-ray Absorption Spectroscopy (XAS) corroboration of the uptake and storage of CeO2 nanoparticles and assessment of their differential toxicity in four edible plant species. J. Agric. Food Chem. 2010, 58, 3689–3693. [Google Scholar] [CrossRef] [PubMed]
- Larue, C.; Castillo-Michel, H.; Sobanska, S.; Trcera, N.; Sorieul, S.; Cécillon, L.; Ouerdane, L.; Legros, S.; Sarret, S. Fate of pristine TiO2 nanoparticles and aged paint-containing TiO2 nanoparticles in lettuce crop after foliar exposure. J. Hazard. Mater. 2014, 273, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Bradford, S.A.; Simunek, J.; Heggen, M.; Vereeken, H.; Klumpp, E. Retention and remobilization of stabilized silver nanoparticles in an undisturbed loamy sand soil. Environ. Sci. Technol. 2013, 47, 12229–12237. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Kuang, L.; He, X.; Bai, W.; Ding, Y.; Zhang, Z.; Zhao, Y.; Chai, Z. Effects of rare earth oxide nanoparticles on root elongation of plants. Chemosphere 2010, 78, 273–279. [Google Scholar] [CrossRef] [PubMed]
- López-Moreno, M.L.; de la Rosa, G.; Hernández-Viezcas, J.A.; Castillo-Michel, H.; Botez, C.E.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Evidence of the differential biotransformation and genotoxicity of ZnO and CeO2 nanoparticles on soybean (Glycine max) plants. Environ. Sci. Technol. 2010, 44, 7315–7320. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, P.; Zhang, Z.; He, X.; Zhang, J.; Ding, Y.; Zhang, J.; Zheng, L.; Guo, Z.; Zhang, L.; et al. Where does the transformation of precipitated ceria nanoparticles in hydroponic plants take place? Environ. Sci. Technol. 2015, 49, 10667–10674. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, S.; Peralta-Videa, J.R.; Bandyopadhyay, S.; Castillo-Michel, H.; Hernandez-Viezcas, J.-A.; Sahi, S.; Gardea-Torresdey, J.L. Exposure of cerium oxide nanoparticles to kidney bean shows disturbance in the plant defense mechanisms. J. Hazard. Mater. 2014, 278, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Rico, C.M.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Differential effects of cerium oxide nanoparticles on rice, wheat, and barley roots: A Fourier Transform Infrared (FT-IR) microspectroscopy study. Appl. Spectrosc. 2015, 69, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Rico, C.M.; Barrios, A.C.; Tan, W.; Rubenecia, R.; Lee, S.C.; Varela-Ramirez, A.; Peralta-Videa, J.-R.; Gardea Torresdey, J.L. Physiological and biochemical response of soil-grown barley (Hordeum vulgare L.) to cerium oxide nanoparticles. Environ. Sci. Pollut. Res. 2015, 22, 10551–10558. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.; Zhou, J.; Liu, C.; Yang, F.; Wu, C.; Zheng, L.; Yang, P. Effect of nano-TiO2 on photochemical reaction of chloroplasts of spinach. Biol. Trace Elem. Res. 2005, 105, 269–279. [Google Scholar] [CrossRef]
- Gao, F.; Hong, F.; Liu, C.; Zheng, L.; Su, M.; Wu, X.; Yang, F.; Wu, C.; Yang, P. Mechanism of nano-anatase TiO2 on promoting photosynthetic carbon reaction of spinach. Biol. Trace Elem. Res. 2006, 111, 239–253. [Google Scholar] [CrossRef]
- Yang, F.; Liu, C.; Gao, F.; Su, M.; Wu, X.; Zheng, L.; Hong, F.; Yang, P. The improvement of spinach growth by nano-anatase TiO2 treatment is related to nitrogen photoreduction. Biol. Trace Elem. Res. 2007, 119, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Linglan, M.; Chao, L.; Chunxiang, Q.; Sitao, Y.; Jie, L.; Fengqing, G.; Fashui, H. Rubisco activase mRNA expression in spinach: Modulation by nanoanatase treatment. Biol. Trace Elem. Res. 2008, 122, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Liu, Y.; Li, T. Nano-TiO2 improve the photosynthesis of tomato leaves under mild heat stress. Biol. Trace Elem. Res. 2013, 156, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Asli, S.; Neumann, M. Colloidal suspensions of clay or titanium dioxide nanoparticles can inhibit leaf growth and transpiration via physical effects on root water transport. Plant Cell Environ. 2009, 32, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Ruffini Castiglione, M.; Giorgetti, L.; Geri, C.; Cremonini, R. The effects of nano-TiO2 on seed germination, development and mitosis of root tip cells of Vicia narbonensis L. and Zea mays L. J. Nanopart. Res. 2011, 1, 2443–2449. [Google Scholar] [CrossRef]
- Song, U.; Shin, M.; Lee, G.; Roh, J.; Kim, Y.; Lee, E.J. Functional analysis of TiO2 nanoparticle toxicity in three plant species. Biol. Trace Elem. Res. 2013, 155, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Frazier, T.P.; Burklew, C.E.; Zhang, B. Titanium dioxide nanoparticles affect the growth and microRNA expression of tobacco (Nicotiana tabacum). Funct. Integr. Genom. 2014, 14, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Pakrashi, S.; Jain, N.; Dalai, S.; Jayakumar, J.; Chandrasekaran, P.T.; Raichur, A.M.; Chandrasekaran, N.; Mukherjee, A. In vivo genotoxicity assessment of titanium dioxide nanoparticles by Allium cepa root tip assay at high exposure concentrations. PLoS ONE 2013. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.K.; McFerran, S.; Lazareva, A.; Suh, S. Global life cycle releases of engineered nanomaterials. J. Nanopart. Res. 2013, 15, 1692. [Google Scholar] [CrossRef]
- Liu, R.; Lal, R. Potentials of engineered nanoparticles as fertilizers for increasing agronomic productions. Sci. Total Environ. 2015, 514, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Kumar, A. Human exposure of engineered nanoparticles from plants irrigated with contaminated waters: Mixture toxicity issues and challenges ahead. Adv. Sci. Lett. 2014, 20, 1204–1207. [Google Scholar] [CrossRef]
- Servin, A.D.; White, J.C. Nanotechnology in agriculture: Next steps for understanding engineered nanoparticle exposure and risk. NanoImpact 2016. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAOSTAT. Available online: http://faostat3.fao.org/download/Q/QC/E (accessed on 20 February 2016).
- Priester, J.H.; Ge, Y.; Mielke, R.E.; Horst, A.M.; Moritz, S.C.; Espinosa, K.; Gelb, J.; Walker, S.L.; Nisbet, R.M.; An, Y.J.; et al. Soybean susceptibility to manufactured nanomaterials with evidence for food quality and soil fertility interruption. Proc. Natl. Acad. Sci. USA 2012. [Google Scholar] [CrossRef] [PubMed]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A Decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Carlino, E. TEM for Characterization of semiconductor nanomaterials. In Transmission Electron Microscopy Characterization of Nanomaterials; Kumar, C.S.S.R., Ed.; Springer: Berlin, Germany, 2014; pp. 89–138. [Google Scholar]
- Colman, B.P.; Arnaout, C.L.; Anciaux, S.; Gunsch, C.K.; Hochella, M.F., Jr.; Kim, B.; Lowry, G.V.; McGill, B.M.; Reinsch, B.C.; Richardson, C.J.; et al. Low concentrations of silver nanoparticles in biosolids cause adverse ecosystem responses under realistic field scenario. PLoS ONE 2013. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-J.; Kwaka, J.I.; Lee, W.-M.; Holden, P.A.; Ana, Y.-J. Zinc oxide nanoparticles delay soybean development: A standard soil microcosm study. Ecotoxicol. Environ. Saf. 2014, 100, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Dofing, S.M. Phenological development-yield relationships in spring barley in a subarctic environment. Can. J. Plant Sci. 1995, 75, 93–97. [Google Scholar] [CrossRef]
- Yang, F.; Hong, F.; You, W.; Liu, C.; Gao, F.; Wu, C.; Yang, P. Influences of nano-anatase TiO2 on the nitrogen metabolism of growing spinach. Biol. Trace Elem. Res. 2006, 110, 179–190. [Google Scholar] [CrossRef]
- Morteza, E.; Moaveni, P.; Farahanim, H.A.; Kiyani, M. Study of photosynthetic pigments changes of maize (Zea mays L.) under nano TiO2 spraying at various growth stages. SpringerPlus 2013. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Sun, Y.; Hernandez-Viezcas, J.A.; Servin, A.D.; Hong, J.; Niu, G.; Peralta-Videa, J.R.; Duarte-Gardea, M.; Gardea-Torresdey, J.L. Influence of CeO2 and ZnO Nanoparticles on cucumber physiological markers and bioaccumulation of Ce and Zn: A life cycle study. J. Agric. Food Chem. 2013, 61, 11945–11951. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, G.; Hund-Rinke, K.; Kuhlbusch, T.; van den Brink, N.; Nickel, C. Fate and bioavailability of engineered nanoparticles in Soils: A review, critical reviews in environmental science and technology. 2014, 44, 2720–2764. [Google Scholar]
- Schlich, K.; Hund-Rinke, K. Influence of soil properties on the effect of silver nanomaterials on microbial activity in five soils. Environ. Pollut. 2015, 196, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, F.; Tanner, S.; Schulin, R.; Rotzetter, A.; Stark, W.; von Quadt, A.; Nowack, B. Dissolved cerium contributes to uptake of Ce in the presence of differently sized CeO2-nanoparticles by three crop plants. Metallomics 2015, 7, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Rico, C.M.; Lee, S.C.; Rubenecia, R.; Mukherjee, A.; Hong, J.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Cerium oxide nanoparticles impact yield and modify nutritional parameters in wheat (Triticum aestivum L.). J. Agric. Food. Chem. 2014, 62, 9669–9675. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Ma, Y.H.; Zhang, Z.Y.; He, X.; Li, Y.Y.; Zhang, J.; Zheng, L.R.; Zhao, Y.L. Species-specific toxicity of ceria nanoparticles to Lactuca plants. Nanotoxicology 2013, 10, 1–8. [Google Scholar]
- Foltête, A.-S.; Masfaraud, J.-F.; Bigorgne, E.; Nahmani, J.; Chaurand, P.; Botta, C.; Labille, J.; Rose, J.; Férard, J.-F.; Cotelle, S. Environmental impact of sunscreen nanomaterials: Ecotoxicity and genotoxicity of altered TiO2 nanocomposites on Vicia Faba. Environ. Pollut. 2001, 159, 2515–2522. [Google Scholar] [CrossRef] [PubMed]
- Kurepa, J.; Paunesku, T.; Vogt, S.; Arora, H.; Rabatic, B.M.; Lu, J.; Wanzer, M.B.; Woloschak, G.E.; Smalle, J.A. Uptake and distribution of ultrasmall anatase TiO2 alizarin red S nanoconjugates in Arabidopsis thaliana. Nano Lett. 2010, 10, 2296–2302. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Bandyopadhyay, M.; Mukherjee, A. Genotoxicity of titanium dioxide (TiO2) nanoparticles at two trophic levels: Plant and human lymphocytes. Chemosphere 2010, 81, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Sohaebuddin, S.K.; Thevenot, P.T.; Baker, D.; Eaton, J.W.; Tang, L. Nanomaterial cytotoxicity is composition, size, and cell type dependent. Part. Fibre Toxicol. 2010, 7, 22–39. [Google Scholar] [CrossRef] [PubMed]
- Panariti, A.; Miserocchi, G.; Rivolta, I. The effect of nanoparticle uptake on cellular behavior: disrupting or enabling functions? Nanotechnol. Sci. Appl. 2012, 5, 87–100. [Google Scholar] [PubMed]
- Lam, E.; Kato, N.; Lawton, M. Programmed cell death, mitochondria and the plant hypersensitive response. Nature 2012, 411, 848–853. [Google Scholar] [CrossRef] [PubMed]
- White, E. Life, death, and the pursuit of apoptosis. Genes Dev. 1996, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kratsch, H.A.; Wise, R.R. The ultrastructure of chilling stress. Plant Cell Environ. 2000, 23, 337–350. [Google Scholar] [CrossRef]
- Zhao, H.; Zhou, Q.; Zhou, M.; Li, C.; Gong, X.; Liu, C.; Qu, C.; Wang, L.; Si, W.; Hong, F. Magnesium deficiency results in damage of nitrogen and carbon cross-talk of maize and improvement by cerium addition. Biol. Trace Elem. Res. 2012, 148, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Viezcas, J.A.; Castillo-Michel, H.; Cooke Andrews, J.; Cotte, M.; Rico, C.; Peralta-Videa, J.R.; Ge, Y.; Priester, J.H.; Holden, P.A.; Gardea-Torresdey, J.L. In situ synchrotron X-ray fluorescence mapping and speciation of CeO2 and ZnO nanoparticles in soil cultivated soybean (Glycine max). ACS Nano 2013, 7, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Ma, Y.; Zhang, Z.; He, X.; Zhang, J.; Guo, Z.; Tai, R.; Zhao, Y.; Chai, Z. Biotransformation of ceria nanoparticles in cucumber plants. ACS Nano 2012, 6, 9943–9950. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, P.; Zhang, Z.; He, X.; Li, Y.; Zhang, J.; Zheng, L.; Chu, S.; Yang, K.; Zhao, Y.; et al. Origin of the different phytotoxicity and biotransformation of cerium and lanthanum oxide nanoparticle in cucumber. Nanotoxicology 2015, 30, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Horie, M.; Nishio, K.; Fujita, K.V.; Endoh, S.; Miyauchi, A.; Saito, Y.; Iwahashi, H.; Yamamoto, K.; Murayama, H.; Nakano, H.; et al. Protein adsorption of ultrafine metal oxide and its influence on cytotoxicity toward cultured cells. Chem. Res. Toxicol. 2009, 22, 543l–5453. [Google Scholar] [CrossRef] [PubMed]
- Servin, A.D.; Castillo-Michel, H.; Hernandez-Viezcas, J.A.; Corral Diaz, B.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Synchrotron micro-XRF and micro-XANES confirmation of the uptake and translocation of TiO2 nanoparticles in cucumber (Cucumis sativus) plants. Environ. Sci. Technol. 2012, 46, 7637–7643. [Google Scholar] [CrossRef] [PubMed]
- Servin, A.D.; Morales, M.I.; Castillo-Michel, H.; Hernandez-Viezcas, J.A.; Munoz, B.; Zhao, L.; Nunez, J.E.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Synchrotron verification of TiO2 accumulation in cucumber fruit: A possible pathway of TiO2 nanoparticle transfer from soil into the food chain. Environ. Sci. Technol. 2013, 47, 11592–11598. [Google Scholar] [CrossRef] [PubMed]
- Balbi, T.; Smerilli, A.; Fabbri, R.; Ciacci, C.; Montagna, M.; Grasselli, E.; Brunelli, A.; Pojana, G.; Marcomini, A.; Gallo, G.; et al. Co-exposure to n-TiO2 and Cd2+ results in interactive effects on biomarker responses but not in increased toxicity in the marine bivalve M. galloprovincialis. Sci. Total Environ. 2014, 493, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Ribas Ferreira, J.L.; Lonné, M.N.; Franc, T.A.; Maximilla, N.M.; Lugokenski, T.H.; Costa, P.G.; Fillmann, G.; Antunes Soares, F.A.; de la Torre, F.R.; Monserrat, J.M. Co-exposure of the organic nanomaterial fullerene C60 with benzo[a]pyrene in Danio rerio (zebrafish) hepatocytes: Evidence of toxicological interactions. Aquat. Toxicol. 2014, 147, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, P.; Anandan, A.; Fernandes, T.F.; Ayoko, G.A.; Biskos, G. Engineered nanomaterials: Knowledge gaps in fate, exposure, toxicity, and future directions. J. Nanomater. 2014. [Google Scholar] [CrossRef]
- Lu, C.M.; Zhang, C.Y.; Wen, J.Q.; Wu, G.R.; Tao, M.X. Research of the effect of nanometer materials on germination and growth enhancement of Glycine max and its mechanism. Soybean Sci. 2002, 21, 168–172. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Booting | Heading | Milk Maturity | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Amax | gs | Tr | Amax | gs | Tr | Amax | gs | Tr | |
| nCeO2 | 0.0003 *** | 0.0335 | 0.0047 ** | ns | ns | ns | ns | ns | ns |
| nTiO2 | 0.0003 *** | 0.0105 ** | 0.0105 ** | ns | ns | ns | ns | ns | ns |
| nCeO2 × nTiO2 | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Ctrl | 20.4 ± 1.8 b | 0.278 ± 0.05 ab | 3.06 ± 0.64 b | 19.4 ± 1.8 a | 0.350 ± 0.06 a | 3.24 ± 0.29 a | 15.3 ± 1.6 a | 0.298 ± 0.05 a | 2.95 ± 0.30 a |
| Ce 500 | 25.7 ± 1.0 a | 0.390 ± 00.5 a | 5.36 ± 0.71 a | 21.2 ± 1.3 a | 0.254 ± 0.02 a | 4.33 ± 0.58 a | 14.4 ± 1.6 a | 0.282 ± 0.04 a | 2.53 ± 0.24 a |
| Ce 1000 | 19.4 ± 0.9 b | 0.249 ± 0.03 b | 3.23 ± 0.51 b | 19.4 ± 1.7 a | 0.220 ± 0.02 a | 4.12 ± 0.33 a | 17.8 ± 1.1 a | 0.349 ± 0.05 a | 3.15 ± 0.32 a |
| Ctrl | 17.5 ± 1.8 b | 0.205 ± 0.06 b | 2.49 ± 0.26 b | 21.6 ± 1.4 a | 0.339 ± 0.04 a | 3.79 ± 0.36 a | 16.4 ± 1.3 a | 0.229 ± 0.03 a | 2.29 ± 0.21 a |
| Ti 500 | 22.8 ± 1.5 ab | 0.287 ± 00.3 ab | 3.73 ± 0.34 ab | 18.5 ± 1.6 a | 0.229 ± 0.03 a | 4.04 ± 0.43 a | 15.7 ± 1.4 a | 0.327 ± 0.05 a | 3.05 ± 0.29 a |
| Ti 1000 | 23.9 ± 1.0 a | 0.387 ± 0.05 a | 3.87 ± 0.45 a | 18.6 ± 1.9 a | 0.230 ± 0.03 a | 3.64 ± 0.62 a | 16.1 ± 1.8 a | 0.357 ± 0.05 a | 3.16 ± 0.30 a |
| Treatment | Ce Roots | Ce Stems | Ce Leaves | Ce Kernels |
|---|---|---|---|---|
| (mg·kg−1) | (mg·kg−1) | (mg·kg−1) | (μg·kg−1) | |
| Ctrl | 3.30 ± 0.63 d | 0.64 ± 0.12 c | 0.73 ± 0.12 c | 0.50 ± 0.19 a |
| Ce 500 | 45.3 ± 11.6 cd | 1.38 ± 0.21 abc | 1.62 ± 0.11 bc | 0.87 ± 0.57 a |
| Ce 1000 | 96.9 ± 1.42 bc | 1.75 ± 0.29 ab | 3.03 ± 0.46 a | 0.69 ± 0.36 a |
| Ti 500 | 14.0 ± 0.97 d | 0.86 ± 0.29 bc | 0.78 ± 0.11 c | 0.98 ± 0.59 a |
| Ti 1000 | 19.1 ± 1.28 d | 0.81 ± 0.21 bc | 0.84 ± 0.09 c | 1.22 ± 0.73 a |
| Ce 500/Ti 500 | 58 ± 9.61 cd | 1.39 ± 0.24 abc | 1.42 ± 0.15 bc | 0.34 ± 0.12 a |
| Ce 500/Ti 1000 | 87 ± 5.60 bc | 1.69 ± 0.23 ab | 1.50 ± 0.08 bc | 1.13 ± 0.52 a |
| Ce 1000/Ti 500 | 149 ± 19.4 ab | 1.98 ± 0.19 a | 1.76 ± 0.14 b | 0.75 ± 0.41 a |
| Ce 1000/Ti 1000 | 164 ± 32 a | 2.01 ± 0.17 a | 1.79 ± 0.17 b | 0.03 ± 0.01 a |
| Treatment | Ti Roots | Ti Stems | Ti Leaves | Ti Kernels |
|---|---|---|---|---|
| (mg·kg−1) | (mg·kg−1) | (mg·kg−1) | (μg·kg−1) | |
| Ctrl | 77 ± 3.19 a | 0.26 ± 0.043 a | 1.03 ± 0.06 a | 1.39 ± 1.39 a |
| Ce 500 | 66.5 ± 5.15 a | 0.19 ± 0.02 a | 1.27 ± 0.32 a | 0.48 ± 0.48 a |
| Ce 1000 | 63.9 ± 2.63 a | 0.31 ± 0.02 a | 1.31 ± 0.28 a | 0.26 ± 0.18 a |
| Ti 500 | 66.7 ± 7.49 a | 0.28 ± 0.03 a | 1.39 ± 0.35 a | 0.71 ± 0.61 a |
| Ti 1000 | 81.7 ± 4.96 a | 0.39 ± 0.06 a | 0.96 ± 0.09 a | <d.l. |
| Ce 500/Ti 500 | 63.9 ± 4.56 a | 0.20 ± 0.09 a | 1.33 ± 0.31 a | 3.62 ± 2.60 a |
| Ce 500/Ti 1000 | 59.4 ± 7 a | 0.22 ± 0.16 a | 1.15 ± 0.27 a | 8.14 ± 4.99 a |
| Ce 1000/Ti 500 | 69.1 ± 7.92 a | 0.22 ± 0.07 a | 1.07 ± 0.22 a | 1.34 ± 1.3 a |
| Ce 1000/Ti 1000 | 68.4 ± 5.41 a | 0.19 ± 0.04 a | 0.89 ± 0.11 a | < d.l. |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchiol, L.; Mattiello, A.; Pošćić, F.; Fellet, G.; Zavalloni, C.; Carlino, E.; Musetti, R. Changes in Physiological and Agronomical Parameters of Barley (Hordeum vulgare) Exposed to Cerium and Titanium Dioxide Nanoparticles. Int. J. Environ. Res. Public Health 2016, 13, 332. https://doi.org/10.3390/ijerph13030332
Marchiol L, Mattiello A, Pošćić F, Fellet G, Zavalloni C, Carlino E, Musetti R. Changes in Physiological and Agronomical Parameters of Barley (Hordeum vulgare) Exposed to Cerium and Titanium Dioxide Nanoparticles. International Journal of Environmental Research and Public Health. 2016; 13(3):332. https://doi.org/10.3390/ijerph13030332
Chicago/Turabian StyleMarchiol, Luca, Alessandro Mattiello, Filip Pošćić, Guido Fellet, Costanza Zavalloni, Elvio Carlino, and Rita Musetti. 2016. "Changes in Physiological and Agronomical Parameters of Barley (Hordeum vulgare) Exposed to Cerium and Titanium Dioxide Nanoparticles" International Journal of Environmental Research and Public Health 13, no. 3: 332. https://doi.org/10.3390/ijerph13030332
APA StyleMarchiol, L., Mattiello, A., Pošćić, F., Fellet, G., Zavalloni, C., Carlino, E., & Musetti, R. (2016). Changes in Physiological and Agronomical Parameters of Barley (Hordeum vulgare) Exposed to Cerium and Titanium Dioxide Nanoparticles. International Journal of Environmental Research and Public Health, 13(3), 332. https://doi.org/10.3390/ijerph13030332