
Frontal Cortex Transcriptome Analysis of Mice Exposed to Electronic Cigarettes During Early Life Stages

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care and Exposures
2.2. E-Cigarette Exposures
2.3. Sample Preparation
2.4. RNA-Seq, Ingenuity Pathway Analyses and Statistical Analyses
2.5. Real-Time Quantitative Polymerase Chain Reaction (qPCR)
2.6. Statistical Analysis for Obstetric Outcomes and qPCR
3. Results
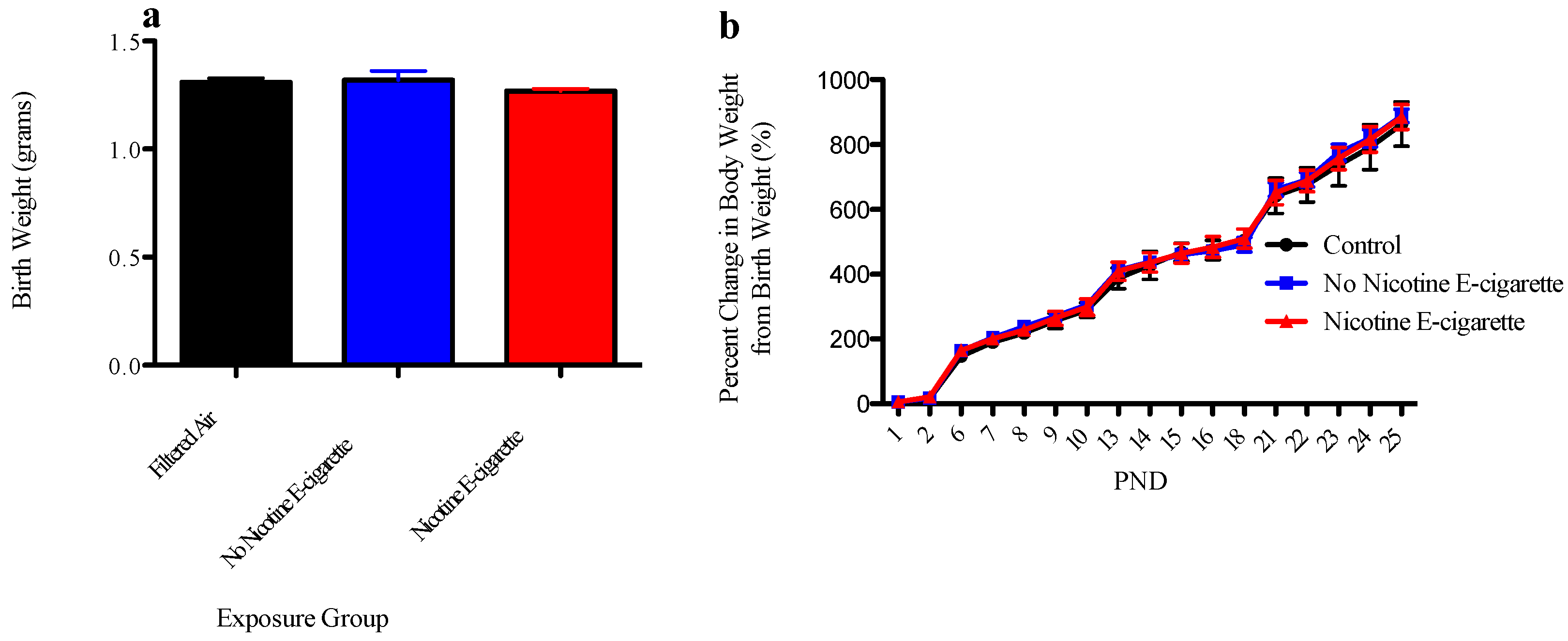
3.1. Effect of E-Cigarette Exposure on Neonatal Growth
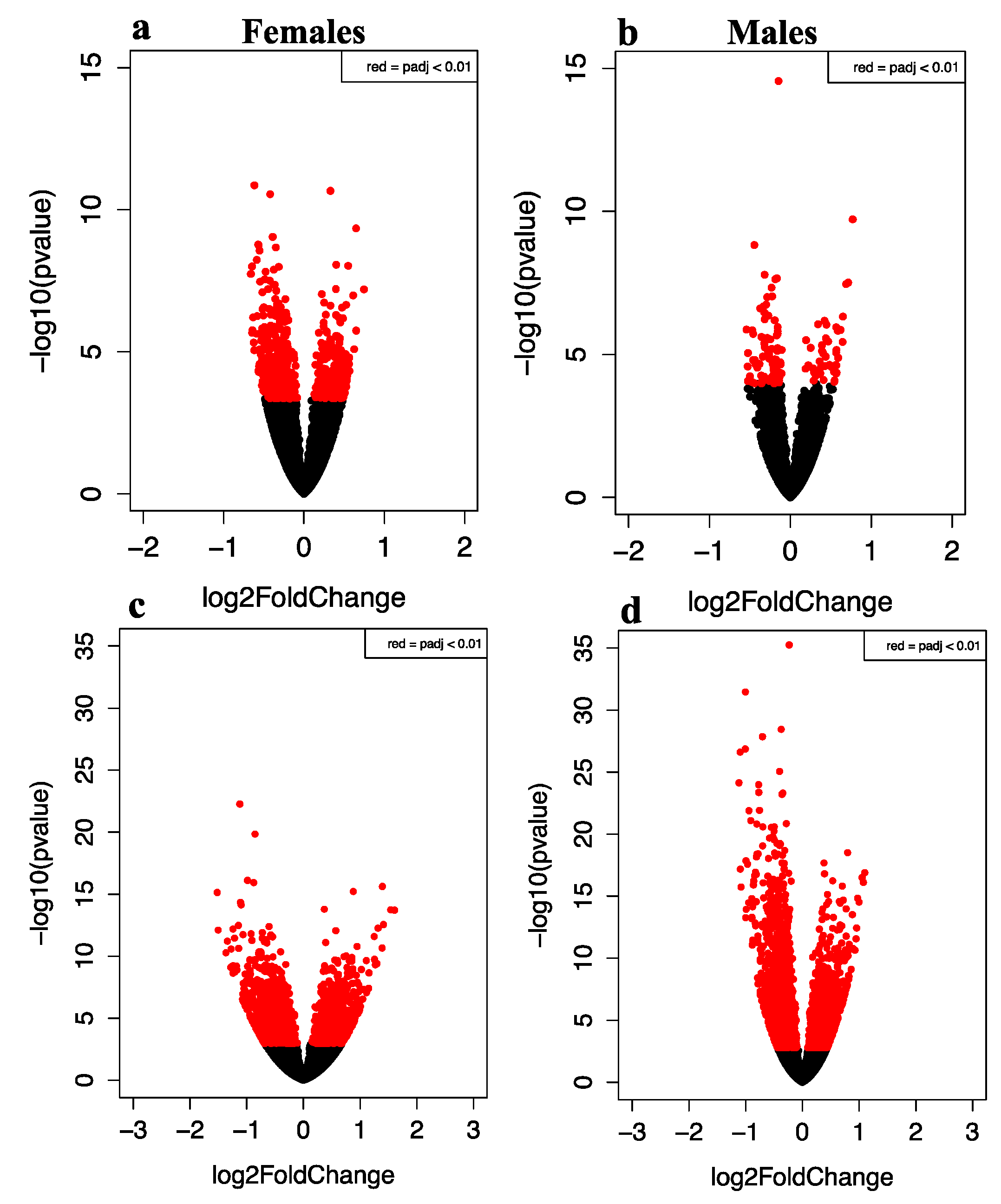
3.2. Global Gene Expression Changes in Murine Frontal Cortex Samples from E-Cigarette Exposed Male and Female Offspring
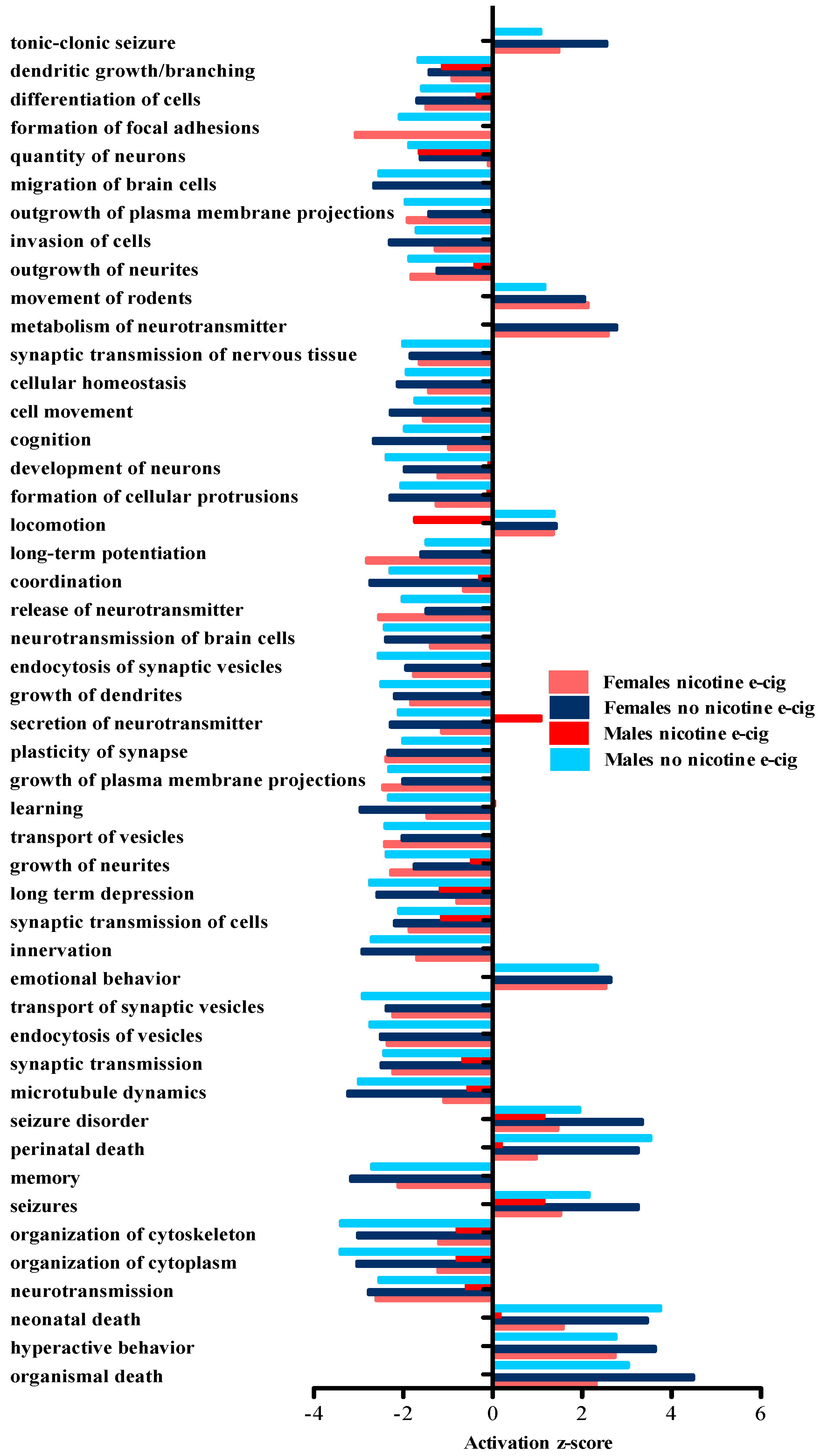
3.3. IPA Predicted Downstream Disease and Biological Function Effects Associated with the Observed Gene Expression Changes
3.4. Follow-Up of Gene Expression Analysis of Selected Genes by qPCR in Male Frontal Cortex Samples
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| E-cigarettes | Electronic cigarettes |
| CNS | central nervous system |
| PND | postnatal day |
| RNA-Seq | RNA Sequencing |
| IPA | Ingenuity® Pathway Analysis |
| GD | gestational day |
| GRAS | generally regarded as safe |
| Ngfr | nerve growth factor receptor |
| Chat | choline O-acetyltransferase |
| Bdnf | brain derived neurotrophic factor |
| Gdnf | glial cell line derived neurotrophic factor |
| Gal | galanin |
| Tbr1 | T-box brain gene 1 |
| Adra1d | alpha-1D adrenoreceptor |
| Gapdh | glyceraldehyde-3-phosphate dehydrogenase |
References
- Arrazola, R.A.; Singh, T.; Corey, C.G.; Husten, C.G.; Neff, L.J.; Apelberg, B.J.; Bunnell, R.E.; Choiniere, C.J.; King, B.A.; Cox, S.; et al. Tobacco use among middle and high school students—United States, 2011–2014. MMWR Morb. Mortal Wkly. Rep. 2015, 14, 381–385. [Google Scholar]
- England, L.J.; Bunnell, R.E.; Pechacek, T.F.; Tong, V.T.; McAfee, T.A. Nicotine and the Developing Human: A Neglected Element in the Electronic Cigarette Debate. Am. J. Prev. Med. 2015, 2, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Agaku, I.T.; King, B.A.; Husten, C.G.; Bunnell, R.; Ambrose, B.K.; Hu, S.S.; Holder-Hayes, E.; Day, H.R. Tobacco Product Use Among Adults—United States 2012–2013. MMWR Morb. Mortal Wkly. Rep. 2014, 25, 542–547. [Google Scholar]
- Farquhar, B.; Mark, K.; Terplan, M.; Chisolm, M.S. Demystifying electronic cigarette use in pregnancy. J. Addict. Med. 2015, 2, 157–158. [Google Scholar] [CrossRef] [PubMed]
- Tong, V.T.; Dietz, P.M.; Farr, S.L.; D’Angelo, D.V.; England, L.J. Estimates of smoking before and during pregnancy: Comparing two population-based data sources. Public Health Rep. 2013, 3, 179–188. [Google Scholar]
- Walton, K.M.; Abrams, D.B.; Bailey, W.C.; Clark, D.; Connolly, G.N.; Djordjevic, M.V.; Eissenberg, T.E.; Fiore, M.C.; Goniewicz, M.L.; Haverkos, L.; et al. NIH electronic cigarette workshop: developing a research agenda. Nicotine Tob. Res. 2015, 2, 259–269. [Google Scholar] [CrossRef] [PubMed]
- National Center for Chronic Disease Prevention and Health Promotion (US) Office on Smoking and Health. The Health Consequences of Smoking—50 Years of Progress: A Report of the Surgeon General. Available online: http://www.ncbi.nlm.nih.gov/books/NBK179276/ (accessed on 1 November 2015).
- Hellström-Lindahl, E.; Gorbounova, O.; Seiger, A.; Mousavi, M.; Nord-berg, A. Regional distribution of nicotinic receptors during prenatal development of human brain and spinal cord. Brain Res. Dev. Brain Res. 1998, 108, 147–160. [Google Scholar] [CrossRef]
- Dwyer, J.B.; McQuown, S.C.; Leslie, F.M. The dynamic effects of nicotine on the developing brain. Pharmacol. Ther. 2009, 2, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Jaspers, M.; de Winter, A.F.; Huisman, M.; Verhulst, F.C.; Ormel, J.; Stewart, R.E.; Reijneveld, S.A. Trajectories of psychosocial problems in adolescents predicted by findings from early well-child assessments. J. Adolesc. Health 2012, 5, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Heijmans, B.T.; Tobi, E.W.; Lumey, L.H.; Slagboom, P.E. The epigenome: Archive of the prenatal environment. Epigenetics. 2009, 8, 526–531. [Google Scholar] [CrossRef]
- Goodwin, R.D.; Perkonigg, A.; Höfler, M.; Wittchen, H.U. Mental disorders and smoking trajectories: A 10-year prospective study among adolescents and young adults in the community. Drug Alcohol Depend. 2013, 130, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Dome, P.; Lazary, J.; Kalapos, M.P.; Rihmer, Z. Smoking, nicotine and neuropsychiatric disorders. Neurosci. Biobehav. Rev. 2010, 3, 295–342. [Google Scholar] [CrossRef] [PubMed]
- Ikegame, T.; Bundo, M.; Murata, Y.; Kasai, K.; Kato, T.; Iwamoto, K. DNA methylation of the BDNF gene and its relevance to psychiatric disorders. J. Hum. Genet. 2013, 7, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Fuchikami, M.; Morinobu, S.; Segawa, M.; Okamoto, Y.; Yamawaki, S.; Ozaki, N.; Inoue, T.; Kusumi, I.; Koyama, T.; Tsuchiyama, K.; et al. DNA methylation profiles of the brain-derived neurotrophic factor (BDNF) gene as a potent diagnostic biomarker in major depression. PLoS ONE 2011, 8. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Borges, G.; Orozco, R.; Mukamal, K.; Rimm, E.B.; Benjet, C.; Medina-Mora, M.E. Exposure to alcohol, drugs and tobacco and the risk of subsequent suicidality: Findings from the Mexican Adolescent Mental Health Survey. Drug Alcohol Depend. 2011, 113, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Yauk, C.L.; Williams, A.; Buick, J.K.; Chen, G.; Maertens, R.M.; Halappanavar, S.; White, P.A. Genetic toxicology and toxicogenomic analysis of three cigarette smoke condensates in vitro reveals few differences among full-flavor, blonde, and light products. Environ. Mol. Mutagen. 2012, 4, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 4, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, J.C. (2007–2015) Venny. An Interactive Tool for Comparing Lists with Venn’s Diagrams. Available online: http:// bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 2 September 2015).
- Jacobsen, L.K.; Slotkin, T.A.; Mencl, W.E.; Frost, S.J.; Pugh, K.R. Gender-specific effects of prenatal and adolescent exposure to tobacco smoke on auditory and visual attention. Neuropsychopharmacology 2007, 12, 2453–2464. [Google Scholar] [CrossRef] [PubMed]
- Blood-Siegfried, J.; Rende, E.K. The long-term effects of prenatal nicotine exposure on neurologic development. J. Midwifery Womens Health 2010, 2, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, J.B.; Broide, R.S.; Leslie, F.M. Nicotine and brain development. Birth Defects Res. C Embryo Today 2008, 1, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Slotkin, T.A. Cholinergic systems in brain development and disruption by neurotoxicants: Nicotine, environmental tobacco smoke, organophosphates. Toxicol. Appl. Pharmacol. 2004, 2, 132–151. [Google Scholar] [CrossRef] [PubMed]
- Slotkin, T.A.; MacKillop, E.A.; Rudder, C.L.; Ryde, I.T.; Tate, C.A.; Seidler, F.J. Permanent, sex-selective effects of prenatal or adolescent nicotine exposure, separately or sequentially, in rat brain regions: Indices of cholinergic and serotonergic synaptic function, cell signaling, and neural cell number and size at 6 months of age. Neuropsychopharmacology 2007, 5, 1082–1097. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Aherrera, A.; Lopez, A.; Neptune, E.; Winickoff, J.P.; Klein, J.D.; Chen, G.; Lazarus, P.; Collaco, J.M.; McGrath-Morrow, S.A. Adult Behavior in Male Mice Exposed to E-Cigarette Nicotine Vapors during Late Prenatal and Early Postnatal Life. PLoS ONE 2015, 9, e0137953. [Google Scholar] [CrossRef] [PubMed]
- Biegon, A.; Alia-Klein, N.; Fowler, J.S. Potential contribution of aromatase inhibition to the effects of nicotine and related compounds on the brain. Front. Pharmacol. 2012, 3, 185. [Google Scholar] [CrossRef] [PubMed]
- Bakker, J.; Honda, S.; Harada, N.; Balthazart, J. Restoration of male sexual behavior by adult exogenous estrogens in male aromatase knockout mice. Horm. Behav. 2004, 1, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, U.; Mullis, P.E. The essential role of the aromatase/p450arom. Semin. Reprod. Med. 2002, 3, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Wang, J.; Dwyer, J.B.; Gautier, N.M.; Wang, S.; Leslie, F.M.; Li, M.D. Gestational nicotine exposure modifies myelin gene expression in the brains of adolescent rats with sex differences. Transl. Psychiatry. 2013, 3, e247. [Google Scholar] [CrossRef] [PubMed]
- Dalla, C.; Antoniou, K.; Papadopoulou-Daifoti, Z.; Balthazart, J.; Bakker, J. Oestrogen-deficient female aromatase knockout (ArKO) mice exhibit depressive-like symptomatology. Eur. J. Neurosci. 2004, 1, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Dalla, C.; Antoniou, K.; Papadopoulou-Daifoti, Z.; Balthazart, J.; Bakker, J. Male aromatase-knockout mice exhibit normal levels of activity, anxiety and “depressive-like” symptomatology. Behav. Brain Res. 2005, 2, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.D.; Chen, W.J. Gender-related response in open-field activity following developmental nicotine exposure in rats. Pharmacol. Biochem. Behav. 2004, 4, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Harrod, S.B.; Mactutus, C.F.; Bennett, K.; Hasselrot, U.; Wu, G.; Welch, M.; Booze, R.M. Sex differences and repeated intravenous nicotine: Behavioral sensitization and dopamine receptors. Pharmacol. Biochem. Behav. 2004, 3, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Vaglenova, J.; Birru, S.; Pandiella, N.M.; Breese, C.R. An assessment of the long-term developmental and behavioral teratogenicity of prenatal nicotine exposure. Behav. Brain Res. 2004, 150, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Farsalinos, K.E.; Romagna, G.; Tsiapras, D.; Kyrzopoulos, S.; Voudris, V. Characteristics, perceived side effects and benefits of electronic cigarette use: A worldwide survey of more than 19,000 consumers. Int. J. Environ. Res. Public Health 2014, 4, 4356–4373. [Google Scholar] [CrossRef] [PubMed]
- Farsalinos, K.E.; Kistler, K.A.; Gillman, G.; Voudris, V. Evaluation of electronic cigarette liquids and aerosol for the presence of selected inhalation toxins. Nicotine Tob. Res. 2015, 2, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Tayyarah, R.; Long, G.A. Comparison of select analytes in aerosol from e-cigarettes with smoke from conventional cigarettes and with ambient air. Regul. Toxicol. Pharmacol. 2014, 3, 704–710. [Google Scholar] [CrossRef] [PubMed]
- Flavors and Extracts Manufacturers’ Association (FEMA). Safety Assessment and Regulatory Authority to Use Flavors: Focus on E-Cigarettes. Available online: https://www.femaflavor.org/safety-assessment-and-regulatory-authority-use-flavors-focus-e-cigarettes (accessed on 3 March 2015).
- Grana, R.; Benowitz, N.; Glantz, S.A. E-cigarettes: A scientific review. Circulation 2014, 19, 1972–1986. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.G.; Flanigan, S.S.; LeBlanc, M.; Vallarino, J.; MacNaughton, P.; Stewart, J.H.; Christiani, D.C. Flavoring Chemicals in E-Cigarettes: Diacetyl, 2,3-Pentanedione, and Acetoin in a Sample of 51 Products, Including Fruit-, Candy-, and Cocktail-Flavored E-Cigarettes. Available online: http://ehp.niehs.nih.gov/wp-content/uploads/advpub/2015/12/ehp.1510185.acco.pdf (accessed on 8 December 2015).
- Kanwal, R.; Kullman, G.; Piacitelli, C.; Boylstein, R.; Sahakian, N.; Martin, S.; Fedan, K.; Kreiss, K. Evaluation of flavorings-related lung disease risk at six microwave popcorn plants. J. Occup. Environ. Med. 2006, 2, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Kreiss, K.; Gomaa, A.; Kullman, G.; Fedan, K.; Simoes, E.J.; Enright, P.L. Clinical bronchiolitis obliterans in workers at a microwave-popcorn plant. N. Engl. J. Med. 2002, 5, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Saffari, A.; Daher, N.; Ruprecht, A.; De Marco, C.; Pozzi, P.; Boffi, R.; Hamad, S.H.; Shafer, M.M.; Schauer, J.J.; Westerdahl, D.; et al. Particulate metals and organic compounds from electronic and tobacco-containing cigarettes: Comparison of emission rates and secondhand exposure. Environ. Sci. Process Impacts 2014, 10, 2259–2267. [Google Scholar] [CrossRef] [PubMed]
- Farsalinos, K.E.; Le Houezec, J. Regulation in the face of uncertainty: The evidence on electronic nicotine delivery systems (e-cigarettes). Risk Manag. Healthc. Policy 2015, 8, 157–167. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Female Offspring Exposed to E-Cigarette Containing Nicotine | ||
| Ingenuity Disease and Disorders | p-Value | # of Molecules |
| Cancer | 5.43 E−05–2.09 E−28 | 1157 |
| Organismal Injury and Abnormalities | 7.31 E−05–2.09 E−28 | 1162 |
| Neurological Disease | 7.75 E−05–1.78 E−27 | 484 |
| Psychiatric Disorders | 7.00 E−05–1.33 E−22 | 320 |
| Gastrointestinal Disease | 5.85 E−05–1.26 E−21 | 838 |
| Female Offspring Exposed to E-Cigarette without Nicotine | ||
| Ingenuity Disease and Disorders | p-Value | # of Molecules |
| Cancer | 6.03 E−06–3.98 E−46 | 2121 |
| Organismal Injury and Abnormalities | 6.82 E−06–3.98 E−46 | 2130 |
| Neurological Disease | 8.19 E−06–1.55 E−35 | 811 |
| Gastrointestinal Disease | 6.03 E−06–3.33 E−35 | 1553 |
| Psychiatric Disorders | 8.19 E−06–5.77 E−31 | 537 |
| Male Offspring Exposed to E-Cigarette Containing Nicotine | ||
| Ingenuity Disease and Disorders | p-Value | # of Molecules |
| Cancer | 1.38 E−02–2.18 E−12 | 137 |
| Organismal Injury and Abnormalities | 1.38 E−02–2.18 E−12 | 137 |
| Dermatological Diseases and Conditions | 6.92 E−03–9.47 E−11 | 80 |
| Connective Tissue Disorders | 6.92 E−03–1.73 E−10 | 83 |
| Gastrointestinal Disease | 1.38 E−02–7.37 E−06 | 99 |
| Male Offspring Exposed to E-Cigarette without Nicotine | ||
| Ingenuity Disease and Disorders | p-Value | # of Molecules |
| Cancer | 2.97 E−06–2.20 E−35 | 2096 |
| Organismal Injury and Abnormalities | 2.97 E−06–2.20 E−35 | 2108 |
| Neurological Disease | 3.27 E−06–7.20 E−34 | 814 |
| Gastrointestinal Disease | 1.10 E−06–1.27 E−30 | 1516 |
| Psychiatric Disorders | 3.27 E−06–1.34 E−27 | 510 |
| Gene | Frontal Cortex Samples from Male Offspring Exposed to E-Cigarettes Containing Nicotine | Frontal Cortex Samples from Male Offspring Exposed to E-Cigarettes Without Nicotine | ||||||
|---|---|---|---|---|---|---|---|---|
| RNA-Seq | qPCR | RNA-Seq | qPCR | |||||
| Fold Change | Adjusted p-Value | Fold Change | p-Value | Fold Change | Adjusted p-Value | Fold Change | p-Value | |
| Ngfr | 2.37 | 3.94E−3 | 2.82 | NS | 3.16 | 1.57E−6 | 4.36 | 0.05 |
| Chat | 1.88 | 6.38E−4 | 2.17 | NS | 2.66 | 9.71E−11 | 3.31 | NS |
| Bdnf | −1.22 | NS | −0.91 | NS | −1.67 | 1.81E−5 | −1.55 | NS |
| Gdnf | ND | NS | 1.47 | NS | 1.93 | 3.6E−5 | 2.13 | NS |
| Gal | ND | NS | 5.6 | 0.001 | 3.66 | 2.01E−6 | 9.56 | 0.001 |
| Tbr1 | −1.31 | NS | −1.25 | NS | −2.02 | 1.68E−11 | −1.91 | NS |
| Adra1d | −1.47 | NS | −2.21 | NS | −2.29 | 9.86E−5 | −3.05 | 0.05 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lauterstein, D.E.; Tijerina, P.B.; Corbett, K.; Akgol Oksuz, B.; Shen, S.S.; Gordon, T.; Klein, C.B.; Zelikoff, J.T. Frontal Cortex Transcriptome Analysis of Mice Exposed to Electronic Cigarettes During Early Life Stages. Int. J. Environ. Res. Public Health 2016, 13, 417. https://doi.org/10.3390/ijerph13040417
Lauterstein DE, Tijerina PB, Corbett K, Akgol Oksuz B, Shen SS, Gordon T, Klein CB, Zelikoff JT. Frontal Cortex Transcriptome Analysis of Mice Exposed to Electronic Cigarettes During Early Life Stages. International Journal of Environmental Research and Public Health. 2016; 13(4):417. https://doi.org/10.3390/ijerph13040417
Chicago/Turabian StyleLauterstein, Dana E., Pamella B. Tijerina, Kevin Corbett, Betul Akgol Oksuz, Steven S. Shen, Terry Gordon, Catherine B. Klein, and Judith T. Zelikoff. 2016. "Frontal Cortex Transcriptome Analysis of Mice Exposed to Electronic Cigarettes During Early Life Stages" International Journal of Environmental Research and Public Health 13, no. 4: 417. https://doi.org/10.3390/ijerph13040417
APA StyleLauterstein, D. E., Tijerina, P. B., Corbett, K., Akgol Oksuz, B., Shen, S. S., Gordon, T., Klein, C. B., & Zelikoff, J. T. (2016). Frontal Cortex Transcriptome Analysis of Mice Exposed to Electronic Cigarettes During Early Life Stages. International Journal of Environmental Research and Public Health, 13(4), 417. https://doi.org/10.3390/ijerph13040417