Nicotine Component of Cigarette Smoke Extract (CSE) Decreases the Cytotoxicity of CSE in BEAS-2B Cells Stably Expressing Human Cytochrome P450 2A13

 , ,
, ,  and
and
Abstract
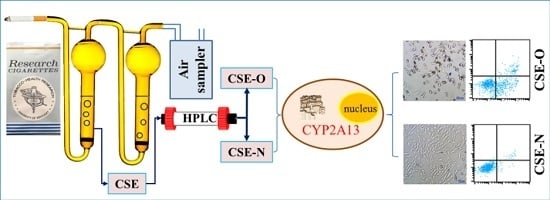
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cigarettes, Chemical and Reagents
2.2. Preparation of CSE and Separation of Nicotine from CSE
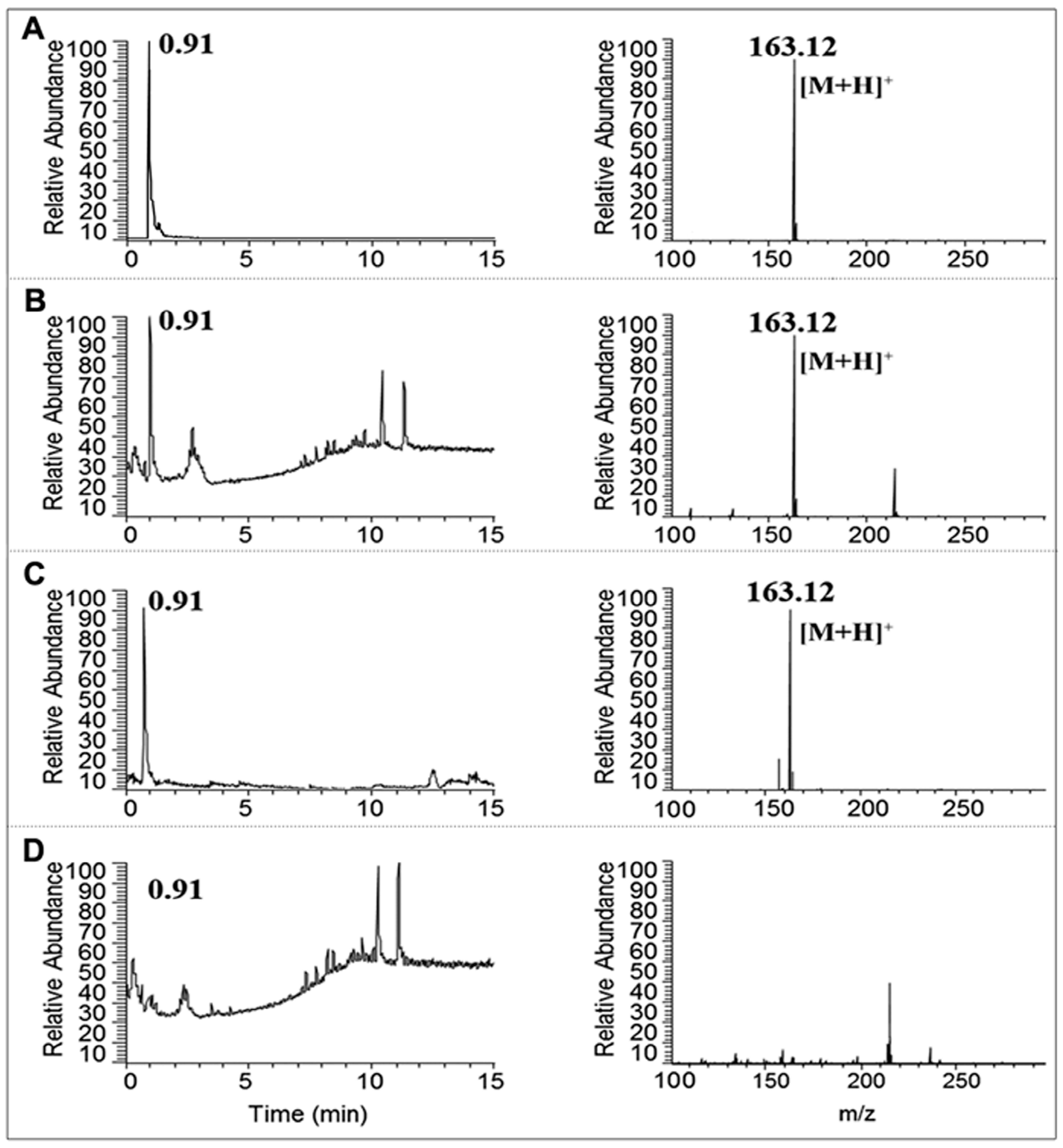
2.3. Identification of Nicotine by UPLC-MS/MS
2.4. Cell Viability Assay
2.5. Cell Apoptosis Analysis
2.6. Determination of the Expression of Apoptosis-Related Proteins in Cells
2.7. Statistical Analysis
3. Results
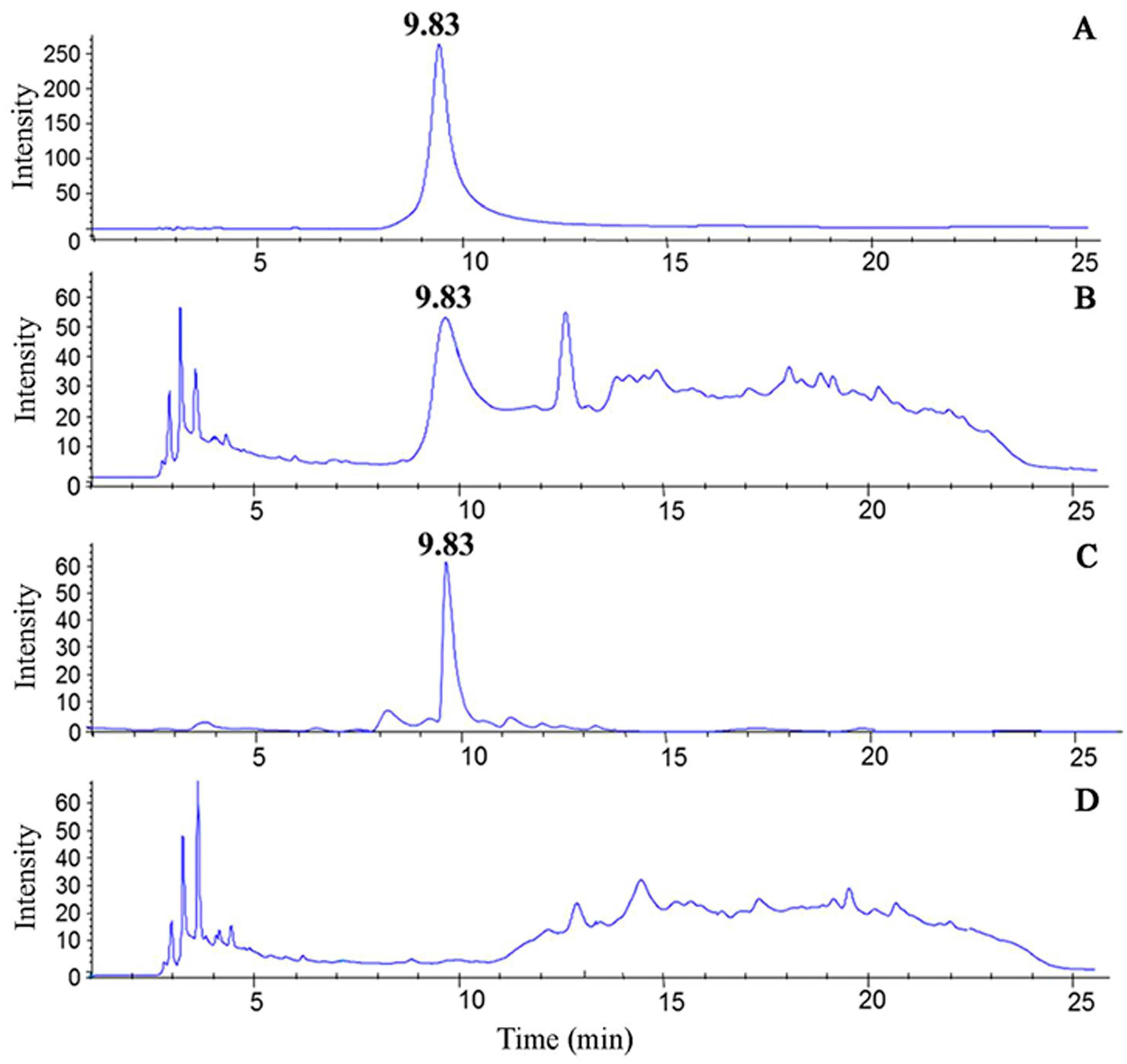
3.1. Separation and Identification of Nicotine in CSE
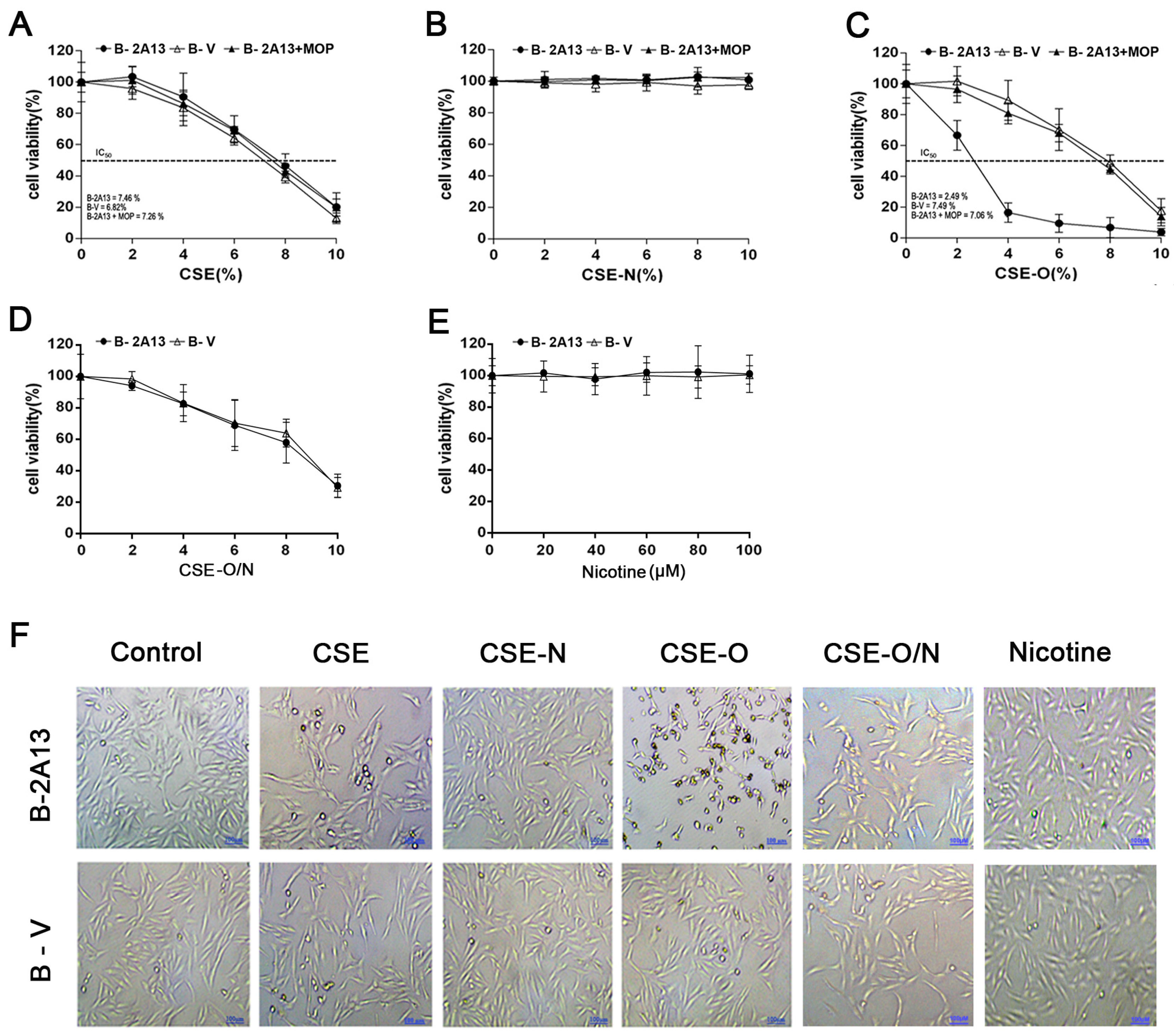
3.2. Effects of Nicotine on CSE-Induced Cytotoxicity in B-2A13 Cells
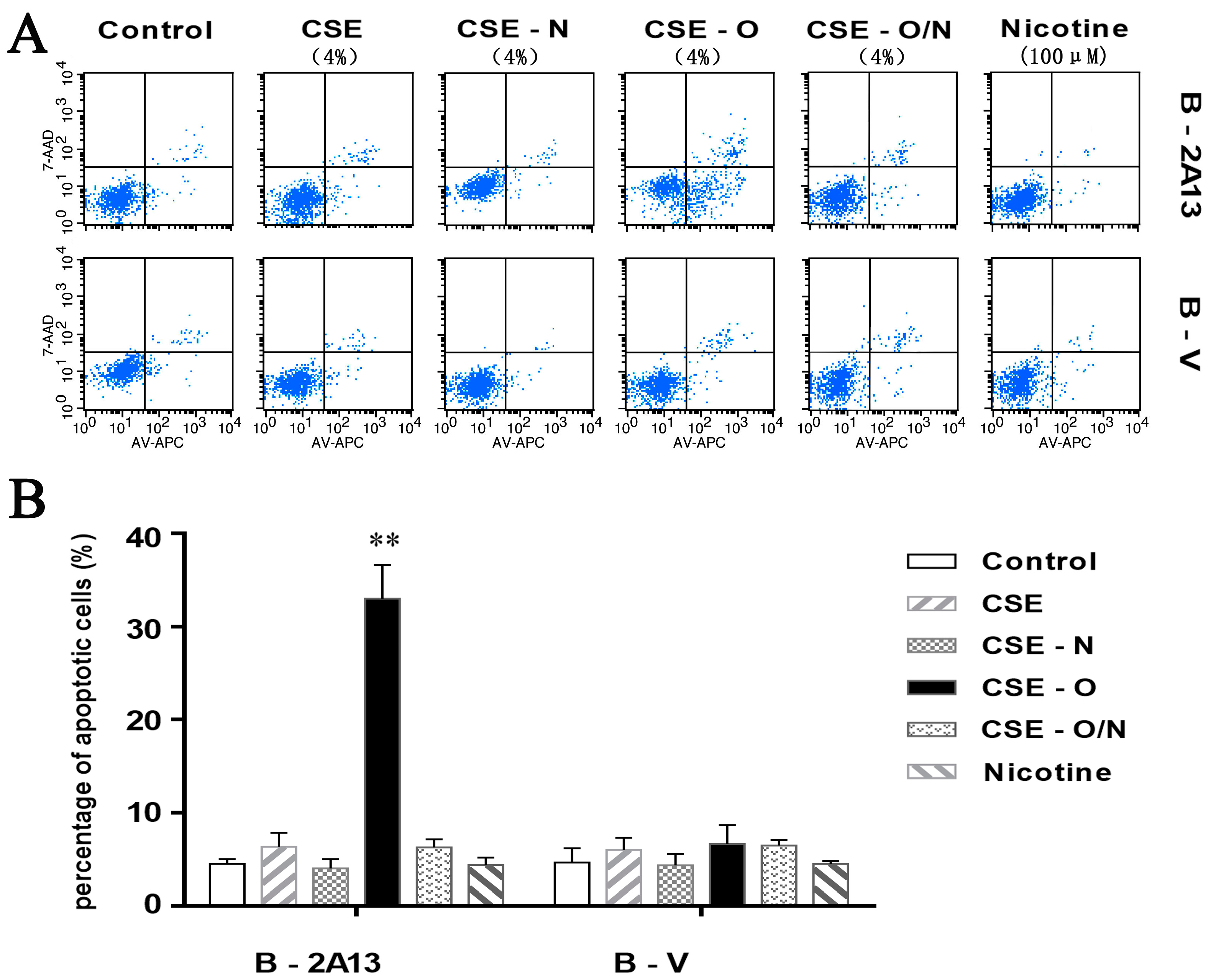
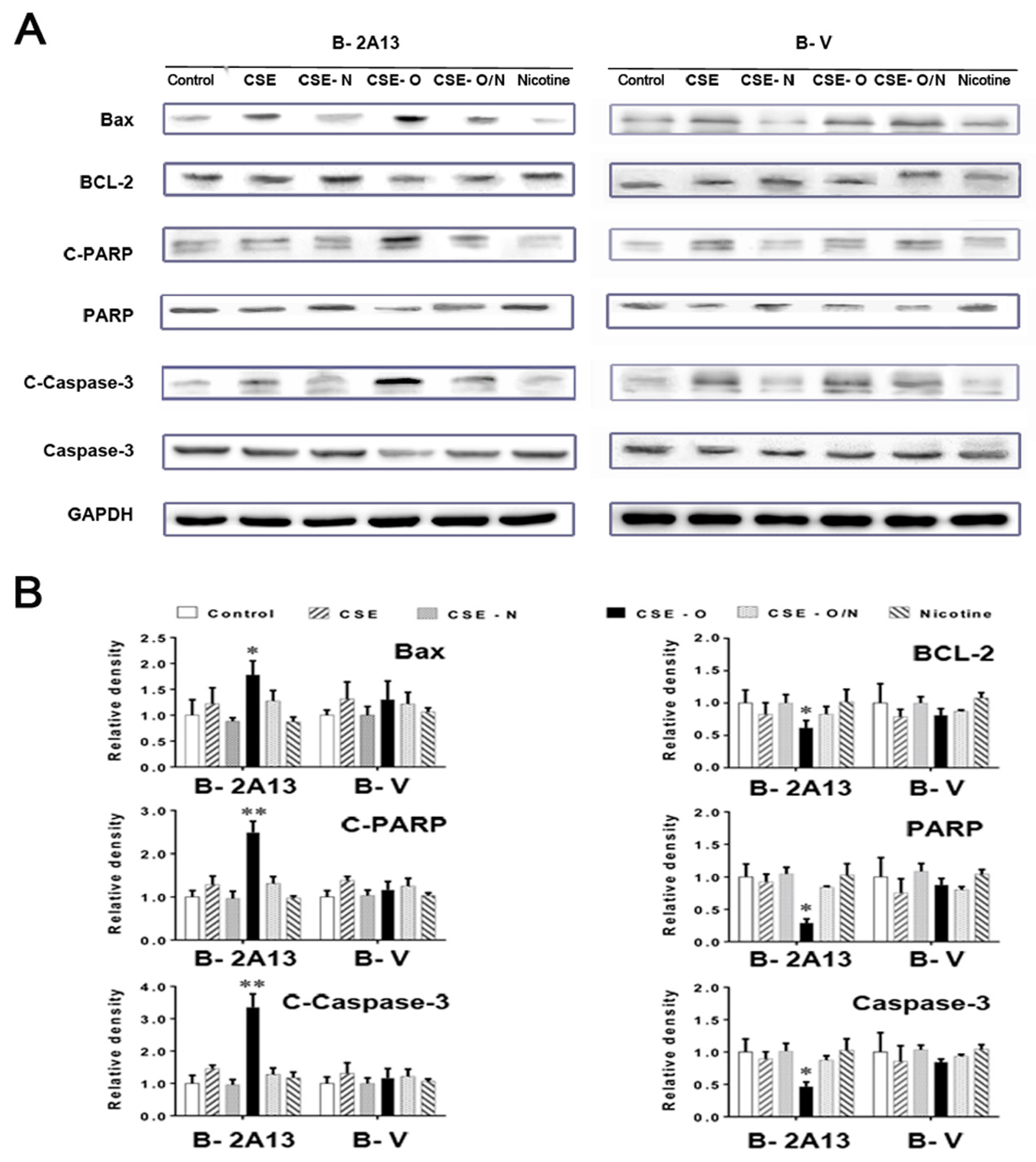
3.3. Effects of Nicotine on CSE-Induced Apoptosis and Expression of Related Proteins in B-2A13 Cells
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CSE | Cigarette smoke extract |
| CYP | Cytochrome P450 |
| CYP2A13 | Cytochrome P450 2A13 |
| NNK | 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone |
| 3-MI | 3-methylindole |
| AFB1 | Aflatoxin B1 |
| BEAS-2B | Immortalized human bronchial epithelial (cells) |
| B-2A13 | BEAS-2B cells stably expressing CYP2A13 |
| HPLC | high-performance liquid chromatography |
| UPLC-MS/MS | Ultra-performance liquid chromatography tandem mass-spectrometry |
| C-PARP | Cleaved Poly (Adenosine Diphosphate-Ribose) Polymerase |
| PARP | Poly (Adenosine Diphosphate-Ribose) Polymerase |
| Bcl-2 | B cell lymphoma-2 |
| Bax | Bcl-2 Associated X Protein |
References
- Bilano, V.; Gilmour, S.; Moffiet, T. Global trends and projections for tobacco use, 1990–2025: An analysis of smoking indicators from the WHO Comprehensive Information Systems for Tobacco Control. Lancet 2015, 385, 966–976. [Google Scholar] [CrossRef]
- Chen, Z.; Peto, R.; Zhou, M.; Iona, A.; Smith, M.; Yang, L.; Guo, Y.; Chen, Y.; Bian, Z.; Lancaster, G.; et al. Contrasting male and female trends in tobacco-attributed mortality in China: Evidence from successive nationwide prospective cohort studies. Lancet 2015, 386, 1447–1456. [Google Scholar] [CrossRef]
- Koplan, J.; Eriksen, M. Smoking cessation for Chinese men and prevention for women. Lancet 2015, 386, 1422–1423. [Google Scholar] [CrossRef]
- Schwartlander, B.; Pratt, A. Tobacco in China: Taming the smoking dragon. Lancet 2015, 385, 2123–2124. [Google Scholar] [CrossRef]
- Sweeney, S.; Theodorou, I.G.; Zambianchi, M.; Chen, S.; Gow, A.; Schwander, S.; Zhang, J.J.; Chung, K.F.; Shaffer, M.S.; Ryan, M.P.; et al. Silver nanowire interactions with primary human alveolar type-II epithelial cell secretions: Contrasting bioreactivity with human alveolar type-I and type-II epithelial cells. Nanoscale 2015, 7, 10398–10409. [Google Scholar] [CrossRef] [PubMed]
- Talhout, R.; Schulz, T.; Florek, E.; van Benthem, J.; Wester, P.; Opperhuizen, A. Hazardous compounds in tobacco smoke. Int. J. Environ. Res. Public Health 2011, 8, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Li, J.; Zheng, M.; Zhao, Y.; Zhou, J.; Li, W. Nnk, a tobacco-specific carcinogen, inhibits the expression of lysyl oxidase, a tumor suppressor. Int. J. Environ. Res. Public Health 2014, 12, 64–82. [Google Scholar] [CrossRef] [PubMed]
- Leon, M.E.; Peruga, A.; McNeill, A.; Kralikova, E.; Guha, N.; Minozzi, S.; Espina, C.; Schuz, J. European Code against Cancer, 4th edition: Tobacco and cancer. Cancer Epidemiol. 2015, S20–S33. [Google Scholar] [CrossRef] [PubMed]
- Chan-Yeung, M.; Dimich-Ward, H. Respiratory health effects of exposure to environmental tobacco smoke. Respirology 2003, 8, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Megaraj, V.; Wei, Y.; Ding, X. Identification of cytochrome P450 enzymes critical for lung tumorigenesis by the tobacco-specific carcinogen 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK): Insights from a novel Cyp2abfgs-null mouse. Carcinogenesis 2014, 35, 2584–2591. [Google Scholar] [CrossRef] [PubMed]
- Oyama, T.; Uramoto, H.; Kagawa, N.; Yoshimatsu, T.; Osaki, T.; Nakanishi, R.; Nagaya, H.; Kaneko, K.; Muto, M.; Kawamoto, T.; et al. Cytochrome P450 in non-small cell lung cancer related to exogenous chemical metabolism. Front. Biosci. 2012, 4, 1539–1546. [Google Scholar] [CrossRef]
- Zhu, L.R.; Thomas, P.E.; Lu, G.; Reuhl, K.R.; Yang, G.Y.; Wang, L.D.; Wang, S.L.; Yang, C.S.; He, X.Y.; Hong, J.Y. Cyp2a13 in human respiratory tissues and lung cancers: An immunohistochemical study with a new peptide-specific antibody. Drug Metab. Dispos. 2006, 34, 1672–1676. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Tan, W.; Hao, B.; Miao, X.; Zhou, G.; He, F.; Lin, D. Substantial reduction in risk of lung adenocarcinoma associated with genetic polymorphism in Cyp2A13, the most active cytochrome P450 for the metabolic activation of tobacco-specific carcinogen NNK. Cancer Res. 2003, 63, 8057–8061. [Google Scholar] [PubMed]
- D’Agostino, J.; Zhang, X.L.; Wu, H.; Ling, G.Y.; Wang, S.P.; Zhang, Q.Y.; Liu, F.C.; Ding, X.X. Characterization of Cyp2A13*2, a variant cytochrome P450 allele previously found to be associated with decreased incidences of lung adenocarcinoma in smokers. Drug Metab. Dispos. 2008, 36, 2316–2323. [Google Scholar] [CrossRef] [PubMed]
- Xiang, C.; Wang, J.C.; Kou, X.C.; Chen, X.B.; Qin, Z.Y.; Jiang, Y.; Sun, C.; Xu, J.B.; Tan, W.; Jin, L.; et al. Pulmonary expression of Cyp2A13 and ABCB1 is regulated by FOXA2, and their genetic interaction is associated with lung cancer. FASEB J. 2015, 29, 1986–1998. [Google Scholar] [CrossRef] [PubMed]
- Megaraj, V.; Zhou, X.; Xie, F.; Liu, Z.; Yang, W.; Ding, X. Role of Cyp2A13 in the bioactivation and lung tumorigenicity of the tobacco-specific lung procarcinogen 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone: In vivo studies using a Cyp2A13-humanized mouse model. Carcinogenesis 2014, 35, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Jalas, J.R.; Hecht, S.S.; Murphy, S.E. Cytochrome P450 enzymes as catalysts of metabolism of 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone, a tobacco specific carcinogen. Chem. Res. Toxicol. 2005, 18, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Takenaka, S.; Murayama, N.; Kramlinger, V.M.; Kim, J.H.; Kim, D.; Liu, J.; Foroozesh, M.K.; Yamazaki, H.; Guengerich, F.P.; et al. Oxidation of pyrene, 1-hydroxypyrene, 1-nitropyrene and 1-acetylpyrene by human cytochrome P450 2A13. Xenobiotica 2016, 46, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Ohgaki, H.; Kusama, K.; Matsukura, N.; Morino, K.; Hasegawa, H.; Sato, S.; Takayama, S.; Sugimura, T. Carcinogenicity in mice of a mutagenic compound, 2-amino-3-methylimidazo[4,5-f]quinoline, from broiled sardine, cooked beef and beef extract. Carcinogenesis 1984, 5, 921–924. [Google Scholar] [CrossRef] [PubMed]
- Benowitz, N.L.; Hukkanen, J.; Jacob, P., III. Nicotine Chemistry, Metabolism, Kinetics and Biomarkers. Handb. Exp. Pharmacol. 2009, 29–60. [Google Scholar] [CrossRef]
- Murphy, S.E.; Raulinaitis, V.; Brown, K.M. Nicotine 5’-oxidation and methyl oxidation by P450 2A enzymes. Drug Metab. Dispos. 2005, 33, 1166–1173. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.J.; Lu, H.Y.; Li, Z.Y.; Bian, Q.; Qiu, L.L.; Li, Z.; Liu, Q.; Li, J.; Wang, X.; Wang, S.L. Cytochrome P450 2A13 mediates aflatoxin B1-induced cytotoxicity and apoptosis in human bronchial epithelial cells. Toxicology 2012, 300, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Von Weymarn, L.B.; Brown, K.M.; Murphy, S.E. Inactivation of CYP2A6 and CYP2A13 during Nicotine Metabolism. J. Pharmacol. Exp. Ther. 2006, 316, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Comer, D.M.; Elborn, J.S.; Ennis, M. Inflammatory and cytotoxic effects of acrolein, nicotine, acetylaldehyde and cigarette smoke extract on human nasal epithelial cells. BMC Pulm. Med. 2014, 14, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, M.; Earla, R.; Shah, A.; Earla, R.L.; Gupte, R.; Mitra, A.K.; Kumar, A.; Kumar, S. A LC-MS/MS Method for Concurrent Determination of Nicotine Metabolites and Role of CYP2A6 in Nicotine Metabolism in U937 Macrophages: Implications in Oxidative Stress in HIV + Smokers. J. Neuroimmune Pharmacol. 2012, 7, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, X.; Wang, Y.; Wang, X.; Lu, H.; Zhang, X.; Xiao, X.; Li, S.; Wang, X.; Wang, S.L. Cytochrome P450 2A13 is an efficient enzyme in metabolic activation of aflatoxin G1 in human bronchial epithelial cells. Arch. Toxicol. 2013, 87, 1697–1707. [Google Scholar] [CrossRef] [PubMed]
- Von Weymarn, L.B.; Zhang, Q.Y.; Ding, X.; Hollenberg, P.F. Effects of 8-methoxypsoralen on cytochrome P450 2A13. Carcinogenesis 2005, 26, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Noya, Y.; Seki, K.; Asano, H.; Mai, Y.; Horinouchi, T.; Higashi, T.; Terada, K.; Hatate, C.; Hoshi, A.; Nepal, P.; et al. Identification of stable cytotoxic factors in the gas phase extract of cigarette smoke and pharmacological characterization of their cytotoxicity. Toxicology 2013, 314, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Asano, H.; Horinouchi, T.; Mai, Y.; Sawada, O.; Fujii, S.; Nishiya, T.; Minami, M.; Katayama, T.; Iwanaga, T.; Terada, K.; et al. Nicotine- and tar-free cigarette smoke induces cell damage through reactive oxygen species newly generated by PKC-dependent activation of NADPH oxidase. J. Pharmacol. Sci. 2012, 118, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Mei, Q.; Wan, X.; Que, H.; Li, L.; Wan, D. Determination of rutin and isoquercetin contents in Hibisci mutabilis folium in different collection periods by HPLC. J. Chromatogr. Sci. 2015, 53, 1680–1684. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Murayama, N.; Yamazaki, H.; Tanaka, K.; Takenaka, S.; Komori, M.; Kim, D.; Guengerich, F.P. Metabolic activation of polycyclic aromatic hydrocarbons and aryl and heterocyclic amines by human cytochromes P450 2A13 and 2A6. Chem. Res. Toxicol. 2013, 26, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Pouli, A.E.; Hatzinikolaou, D.G.; Piperi, C.; Stavridou, A.; Psallidopoulos, M.C.; Stavrides, J.C. The cytotoxic effect of volatile organic compounds of the gas phase of cigarette smoke on lung epithelial cells. Free Radic. Biol. Med. 2003, 34, 345–355. [Google Scholar] [CrossRef]
- Naik, P.; Fofaria, N.; Prasad, S.; Sajja, R.K.; Weksler, B.; Couraud, P.O.; Romero, I.A.; Cucullo, L. Oxidative and pro-inflammatory impact of regular and denicotinized cigarettes on blood brain barrier endothelial cells: Is smoking reduced or nicotine-free products really safe? BMC Neurosci. 2014, 15, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeVore, N.M.; Scott, E.E. Nicotine and 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone binding and access channel in human cytochrome P450 2A6 and 2A13 enzymes. J. Biol. Chem. 2012, 287, 26576–26585. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Megaraj, V.; Li, L.; Sell, S.; Hu, J.; Ding, X. Suppression of pulmonary cyp2a13 expression by carcinogen-induced lung tumorigenesis in a Cyp2A13-humanized mouse model. Drug Metab. Dispos. Biol. Fate Chem. 2015, 43, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Jia, K.; Li, L.; Liu, Z.; Hartog, M.; Kluetzman, K.; Zhang, Q.Y.; Ding, X. Generation and characterization of a novel Cyp2A13—Transgenic mouse model. Drug Metab. Dispos. 2014, 42, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Van Vleet, T.R.; Bombick, D.W.; Coulombe, R.A., Jr. Inhibition of human cytochrome P450 2E1 by nicotine, cotinine, and aqueous cigarette tar extract in vitro. Toxicol. Sci. 2001, 64, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Yano, J.K.; Denton, T.T.; Cerny, M.A.; Zhang, X.; Johnson, E.F.; Cashman, J.R. Synthetic inhibitors of cytochrome P-450 2A6: Inhibitory activity, difference spectra, mechanism of inhibition, and protein cocrystallization. J. Med. Chem. 2006, 49, 6987–7001. [Google Scholar] [CrossRef] [PubMed]
- Naserzadeh, P.; Hosseini, M.J.; Mohamadzadeh Asl, B.; Pourahmad, J. Toxicity mechanisms of cigarette smoke on mouse fetus mitochondria. Iran. J. Pharm. Res. 2015, 14, 131–138. [Google Scholar] [PubMed]
- He, X.; Song, W.; Liu, C.; Chen, S.; Hua, J. Rapamycin inhibits acrolein-induced apoptosis by alleviating ROS-driven mitochondrial dysfunction in male germ cells. Cell Prolif. 2014, 47, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.A.; Yang, F.; Cole, G.M.; Chan, S.O. Inhibition of caspase-3-like activity reduces glutamate induced cell death in adult rat retina. Brain Res. 2001, 904, 177–188. [Google Scholar] [CrossRef]
- Williams, M.D.; Sandler, A.B. The epidemiology of lung cancer. Cancer Treat. Res. 2001, 105, 31–52. [Google Scholar] [PubMed]
- Weems, J.M.; Lamb, J.G.; D’Agostino, J.; Ding, X.; Yost, G.S. Potent mutagenicity of 3-methylindole requires pulmonary cytochrome P450-mediated bioactivation: A comparison to the prototype cigarette smoke mutagens B(a)P and NNK. Chem. Res. Toxicol. 2010, 23, 1682–1690. [Google Scholar] [CrossRef] [PubMed]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, M.; Zhang, Y.; Li, N.; Wang, C.; Xia, R.; Zhang, Z.; Wang, S.-L. Nicotine Component of Cigarette Smoke Extract (CSE) Decreases the Cytotoxicity of CSE in BEAS-2B Cells Stably Expressing Human Cytochrome P450 2A13. Int. J. Environ. Res. Public Health 2017, 14, 1221. https://doi.org/10.3390/ijerph14101221
Ji M, Zhang Y, Li N, Wang C, Xia R, Zhang Z, Wang S-L. Nicotine Component of Cigarette Smoke Extract (CSE) Decreases the Cytotoxicity of CSE in BEAS-2B Cells Stably Expressing Human Cytochrome P450 2A13. International Journal of Environmental Research and Public Health. 2017; 14(10):1221. https://doi.org/10.3390/ijerph14101221
Chicago/Turabian StyleJi, Minghui, Yudong Zhang, Na Li, Chao Wang, Rong Xia, Zhan Zhang, and Shou-Lin Wang. 2017. "Nicotine Component of Cigarette Smoke Extract (CSE) Decreases the Cytotoxicity of CSE in BEAS-2B Cells Stably Expressing Human Cytochrome P450 2A13" International Journal of Environmental Research and Public Health 14, no. 10: 1221. https://doi.org/10.3390/ijerph14101221
APA StyleJi, M., Zhang, Y., Li, N., Wang, C., Xia, R., Zhang, Z., & Wang, S. -L. (2017). Nicotine Component of Cigarette Smoke Extract (CSE) Decreases the Cytotoxicity of CSE in BEAS-2B Cells Stably Expressing Human Cytochrome P450 2A13. International Journal of Environmental Research and Public Health, 14(10), 1221. https://doi.org/10.3390/ijerph14101221