The Occurrence of the Colistin Resistance Gene mcr-1 in the Haihe River (China)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specific Primers and Probe Design
2.2. Preparation of Standard Curve
2.3. qPCR Procedures
2.4. Detection of mcr-1 Genes in Haihe River
2.5. Statistical Analysis
3. Results
3.1. Development of qPCR
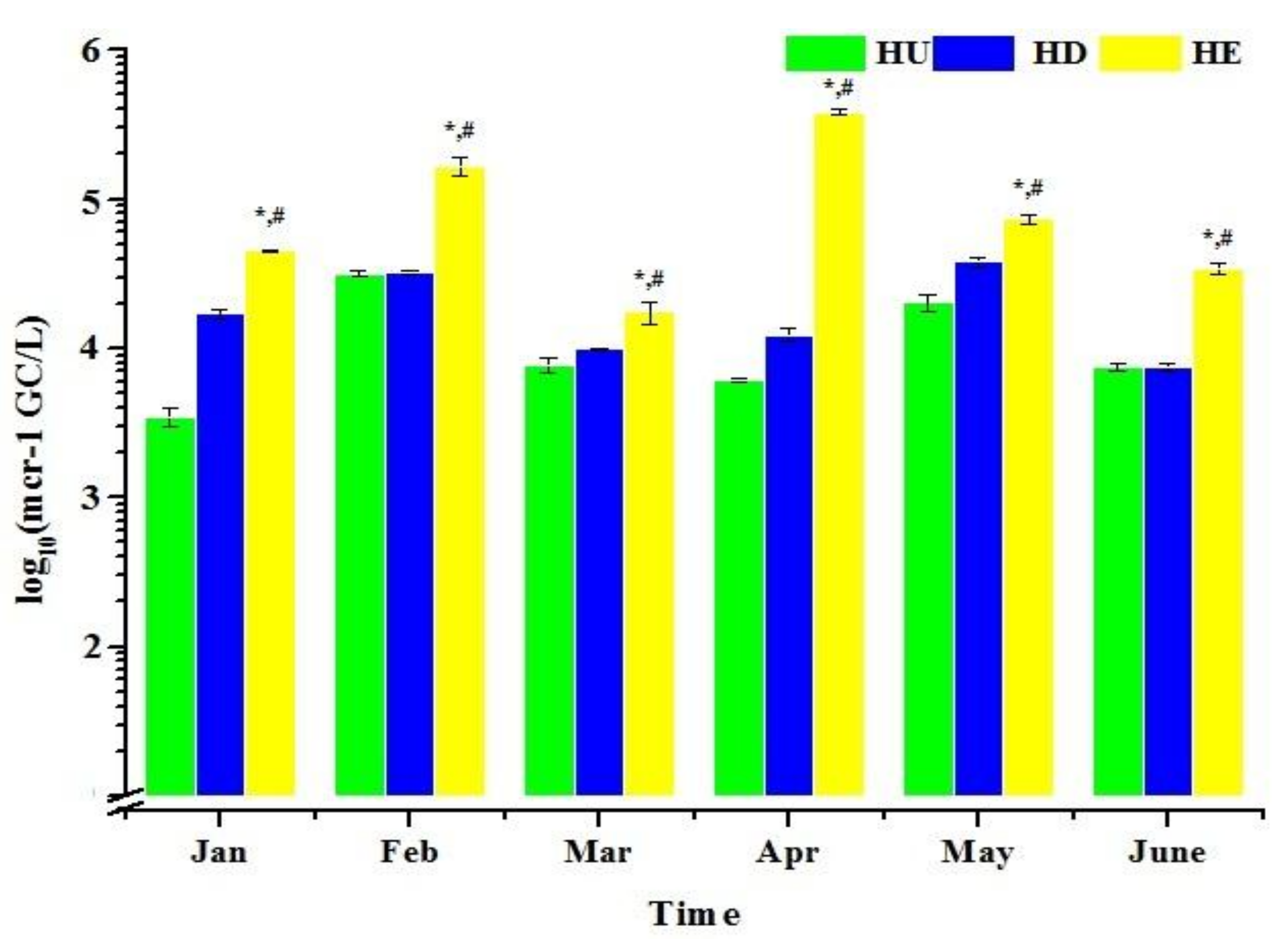
3.2. The mcr-1 Genes in Haihe River
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflict of Interest
References
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. P&T. 2015, 40, 277–283. [Google Scholar] [PubMed]
- Woolhouse, M.E.; Ward, M.J. Microbiology. Sources of antimicrobial resistance. Science 2013, 341, 1460–1461. [Google Scholar] [CrossRef] [PubMed]
- Neu, H.C. The crisis in antibiotic resistance. Science 1992, 257, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, J.G.; Gilbert, D.N.; Spellberg, B. Seven ways to preserve the miracle of antibiotics. Clin. Infect. Dis. 2013, 56, 1445–1450. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Antimicrobial Resistance Threat Report; CDC: Atlanta, GA, USA, 2013.
- World Health Organization (WHO). Antibiotic Resistance; WHO: Geneva, Switzerland, 2016. [Google Scholar]
- Perry, J.A.; Westman, E.L.; Wright, G.D. The antibiotic resistome: What’s new? Curr. Opin. Microbiol. 2014, 21, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Mah, M.W.; Memish, Z.A. Antibiotic resistance. An impending crisis. Saudi Med. J. 2000, 21, 1125–1129. [Google Scholar] [PubMed]
- Mazel, D. Integrons: Agents of bacterial evolution. Nat. Rev. Microbiol. 2006, 4, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Aminov, R.I.; Mackie, R.I. Evolution and ecology of antibiotic resistance genes. FEMS Microbiol. Lett. 2007, 271, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Marti, E.; Variatza, E.; Balcazar, J.L. The role of aquatic ecosystems as reservoirs of antibiotic resistance. Trends Microbiol. 2014, 22, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Xi, C.; Zhang, Y.; Marrs, C.F.; Ye, W.; Simon, C.; Foxman, B.; Nriagu, J. Prevalence of antibiotic resistance in drinking water treatment and distribution systems. Appl. Environ. Microbiol. 2009, 75, 5714–5718. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.X.; Zhang, T.; Fang, H.H. Antibiotic resistance genes in water environment. Appl. Microbiol. Biotechnol. 2009, 82, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.R.; Weeks, J.; Livermore, D.M.; Toleman, M.A. Dissemination of ndm-1 positive bacteria in the new delhi environment and its implications for human health: An environmental point prevalence study. Lancet Infect. Dis. 2011, 11, 355–362. [Google Scholar] [CrossRef]
- Paterson, D.L.; Harris, P.N. Colistin resistance: A major breach in our last line of defence. Lancet Infect. Dis. 2016, 16, 132–133. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism mcr-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Tse, H.; Yuen, K.Y. Dissemination of the mcr-1 colistin resistance gene. Lancet Infect. Dis. 2016. [CrossRef]
- Olaitan, A.O.; Chabou, S.; Okdah, L.; Morand, S.; Rolain, J.M. Dissemination of the mcr-1 colistin resistance gene. Lancet Infect. Dis. 2016, 16, 147. [Google Scholar] [CrossRef]
- Malhotra-Kumar, S.; Xavier, B.B.; Das, A.J.; Lammens, C.; Butaye, P.; Goossens, H. Colistin resistance gene mcr-1 harboured on a multidrug resistant plasmid. Lancet Infect. Dis. 2016, 16, 283–284. [Google Scholar] [CrossRef]
- Yao, X.; Doi, Y.; Zeng, L.; Lv, L.; Liu, J.H. Carbapenem-resistant and colistin-resistant escherichia coli co-producing ndm-9 and mcr-1. Lancet Infect. Dis. 2016, 16, 288–289. [Google Scholar] [CrossRef]
- Martinez, J.L. Antibiotics and antibiotic resistance genes in natural environments. Science 2008, 321, 365–367. [Google Scholar] [CrossRef] [PubMed]
- Calero-Caceres, W.; Melgarejo, A.; Colomer-Lluch, M.; Stoll, C.; Lucena, F.; Jofre, J.; Muniesa, M. Sludge as a potential important source of antibiotic resistance genes in both the bacterial and bacteriophage fractions. Environ.Sci. Technol. 2014, 48, 7602–7611. [Google Scholar] [CrossRef] [PubMed]
- Calero-Caceres, W.; Muniesa, M. Persistence of naturally occurring antibiotic resistance genes in the bacteria and bacteriophage fractions of wastewater. Water Res. 2016, 95, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jin, M.; Qiu, Z.G.; Guo, C.; Chen, Z.L.; Shen, Z.Q.; Wang, X.W.; Li, J.W. A survey of drug resistance bla genes originating from synthetic plasmid vectors in six Chinese rivers. Environ. Sci. Technol. 2012, 46, 13448–13454. [Google Scholar] [CrossRef] [PubMed]
- Chabou, S.; Leangapichart, T.; Okdah, L.; Le Page, S.; Hadjadj, L.; Rolain, J.M. Real-time quantitative PCR assay with Taqman® probe for rapid detection of mcr-1 plasmid-mediated colistin resistance. New Microbes New Infect. 2016, 13, 71–74. [Google Scholar] [CrossRef] [PubMed]
- El-Tahawy, A.T. The crisis of antibiotic-resistance in bacteria. Saudi Med. J. 2004, 25, 837–842. [Google Scholar] [PubMed]
- Lushniak, B.D. Antibiotic resistance: A public health crisis. Public Health Rep. 2014, 129, 314–316. [Google Scholar] [PubMed]
- Kempf, I.; Fleury, M.A.; Drider, D.; Bruneau, M.; Sanders, P.; Chauvin, C.; Madec, J.Y.; Jouy, E. What do we know about resistance to colistin in Enterobacteriaceae in avian and pig production in Europe? Int. J. Antimicrob. Agents 2013, 42, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, J.; Corcoran, C.; Prentice, E.; Moodley, M.; Mendelson, M.; Poirel, L.; Nordmann, P.; Brink, A.J. Emergence of plasmid-mediated colistin resistance (mcr-1) among escherichia coli isolated from South African patients. S. Afr. Med. J. 2016, 106, 35–36. [Google Scholar] [CrossRef] [PubMed]
- Falgenhauer, L.; Waezsada, S.E.; Yao, Y.; Imirzalioglu, C.; Kasbohrer, A.; Roesler, U.; Michael, G.B.; Schwarz, S.; Werner, G.; Kreienbrock, L.; et al. Colistin resistance gene mcr-1 in extended-spectrum beta-lactamase-producing and carbapenemase-producing gram-negative bacteria in Germany. Lancet Infect. Dis. 2016, 16, 282–283. [Google Scholar] [CrossRef]
- Se´verine Bontron, L.P. Patrice Nordmann Real-time PCR for detection of plasmid-mediated polymyxin resistance (mcr-1) from cultured bacteria and stools. J. Antimicrob. Chemother. 2016, 71, 3. [Google Scholar]
- Li, H.-X. Preparation and comparison on function detecting of the two types of Taqman probes. Chem. Res. Appl. 2011, 23, 5. [Google Scholar]
- Ahmed, T.; Scholz, M.; Al-Faraj, F.; Niaz, W. Water-related impacts of climate change on agriculture and subsequently on public health: A review for generalists with particular reference to Pakistan. Int. J. Environ. Res. Public Health 2016, 13, 1051. [Google Scholar] [CrossRef] [PubMed]
- Beach, M. Water, pollution, and public health in China. Lancet 2001, 358, 735. [Google Scholar] [CrossRef]
- Allen, H.K.; Donato, J.; Wang, H.H.; Cloud-Hansen, K.A.; Davies, J.; Handelsman, J. Call of the wild: Antibiotic resistance genes in natural environments. Nat. Rev. Microbiol. 2010, 8, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Yu, Y.; Chen, Z.; Jin, M.; Yang, D.; Zhao, Z.; Wang, J.; Shen, Z.; Wang, X.; Qian, D. Nanoalumina promotes the horizontal transfer of multiresistance genes mediated by plasmids across genera. Proc. Natl. Acad. Sci. USA 2012, 109, 4944–4949. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Shen, Z.; Qian, D.; Jin, M.; Yang, D.; Wang, J.; Zhang, B.; Yang, Z.; Chen, Z.; Wang, X.; et al. Effects of nano-tio2 on antibiotic resistance transfer mediated by rp4 plasmid. Nanotoxicology 2015, 9, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Mei-Ting Guo, Q.-B.Y.; Yang, J. Distinguishing effects of ultraviolet exposure and chlorination on the horizontal transfer of antibiotic resistance genes in municipal wastewater. Environ. Sci. Technol. 2015, 49, 9. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer/Probe Name | Target Sequences | Sequence (5’–3’) | Tm (°C) | Product Length (bp) | Reference |
|---|---|---|---|---|---|
| qPCR | locus region 22541-22635 in the KP347227 | 116 | This study | ||
| M-F | CATCGCGGACAATCTCGG | 57.2 | |||
| M-R | AAATCAACACAGGCTTTAGCAC | 55.9 | |||
| M-Probe | FAM-AACAGCGTGGTGATCAGTAGCAT-BHQ | 61.4 | |||
| Standard-PCR | locus region 22447-22755 in the KP347227 | 305 | [16] | ||
| CLR5-F | CGGTCAGTCCGTTTGTTC | 56.1 | |||
| CLR5-R | CTTGGTCGGTCTGTAGGG | 56.3 |
| Species | Presence of mcr-1 Gene | COL MIC(mg/L) | CT Value | Origins |
|---|---|---|---|---|
| E.coli (n = 11) | + | 4–16 | 15–28 | China (water environment) |
| E.coli (n = 7) | − | <1 | <LOQ | |
| K.pneumoniae (n = 2) | + | 8 | 18–28 | |
| K.pneumoniae (n = 10) | − | <1 | <LOQ | |
| Morganellamorganii (n = 3) | − | >256 | <LOQ | |
| Providenciaalcalifaciens (n = 5) | − | <1 | <LOQ | |
| Serratiamarcescens (n = 4) | − | >128 | <LOQ | |
| Yokenellaregensburgei (n = 5) | − | >128 | <LOQ | |
| Aeromonashydrophila (n = 4) | − | >128 | <LOQ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, D.; Qiu, Z.; Shen, Z.; Zhao, H.; Jin, M.; Li, H.; Liu, W.; Li, J.-W. The Occurrence of the Colistin Resistance Gene mcr-1 in the Haihe River (China). Int. J. Environ. Res. Public Health 2017, 14, 576. https://doi.org/10.3390/ijerph14060576
Yang D, Qiu Z, Shen Z, Zhao H, Jin M, Li H, Liu W, Li J-W. The Occurrence of the Colistin Resistance Gene mcr-1 in the Haihe River (China). International Journal of Environmental Research and Public Health. 2017; 14(6):576. https://doi.org/10.3390/ijerph14060576
Chicago/Turabian StyleYang, Dong, Zhigang Qiu, Zhiqiang Shen, Hong Zhao, Min Jin, Huaying Li, Weili Liu, and Jun-Wen Li. 2017. "The Occurrence of the Colistin Resistance Gene mcr-1 in the Haihe River (China)" International Journal of Environmental Research and Public Health 14, no. 6: 576. https://doi.org/10.3390/ijerph14060576
APA StyleYang, D., Qiu, Z., Shen, Z., Zhao, H., Jin, M., Li, H., Liu, W., & Li, J. -W. (2017). The Occurrence of the Colistin Resistance Gene mcr-1 in the Haihe River (China). International Journal of Environmental Research and Public Health, 14(6), 576. https://doi.org/10.3390/ijerph14060576