Effects of Short-Term Set-Aside Management Practices on Soil Microorganism and Enzyme Activity in China


Abstract
:1. Introduction
2. Methods
2.1. Site Description and Experimental Design
2.2. Soil Sampling and Preparation
2.3. Measurements of Soil Physicochemical Property
2.4. Biological Analysis
2.4.1. DNA Extraction and MiSeq Illumina Sequencing
2.4.2. Sequencing Data Processing
2.4.3. Soil Microbial Biomass
2.5. Soil Enzymes
2.6. Statistical Analyses
3. Results
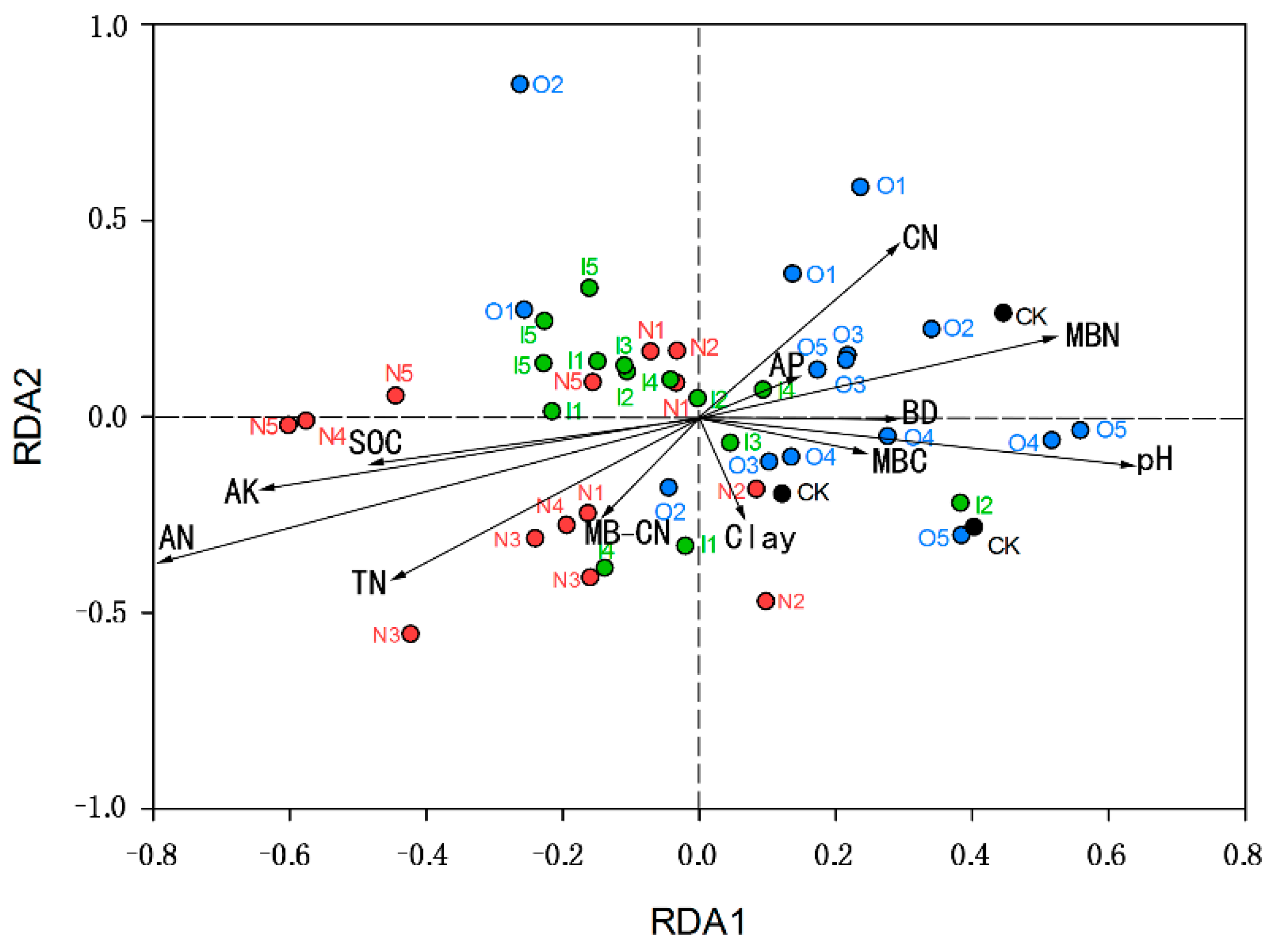
3.1. Analysis of Microbial Community and Influencing Factors on Cultivated Land
3.1.1. Alpha Diversity of Soil Microbial Community
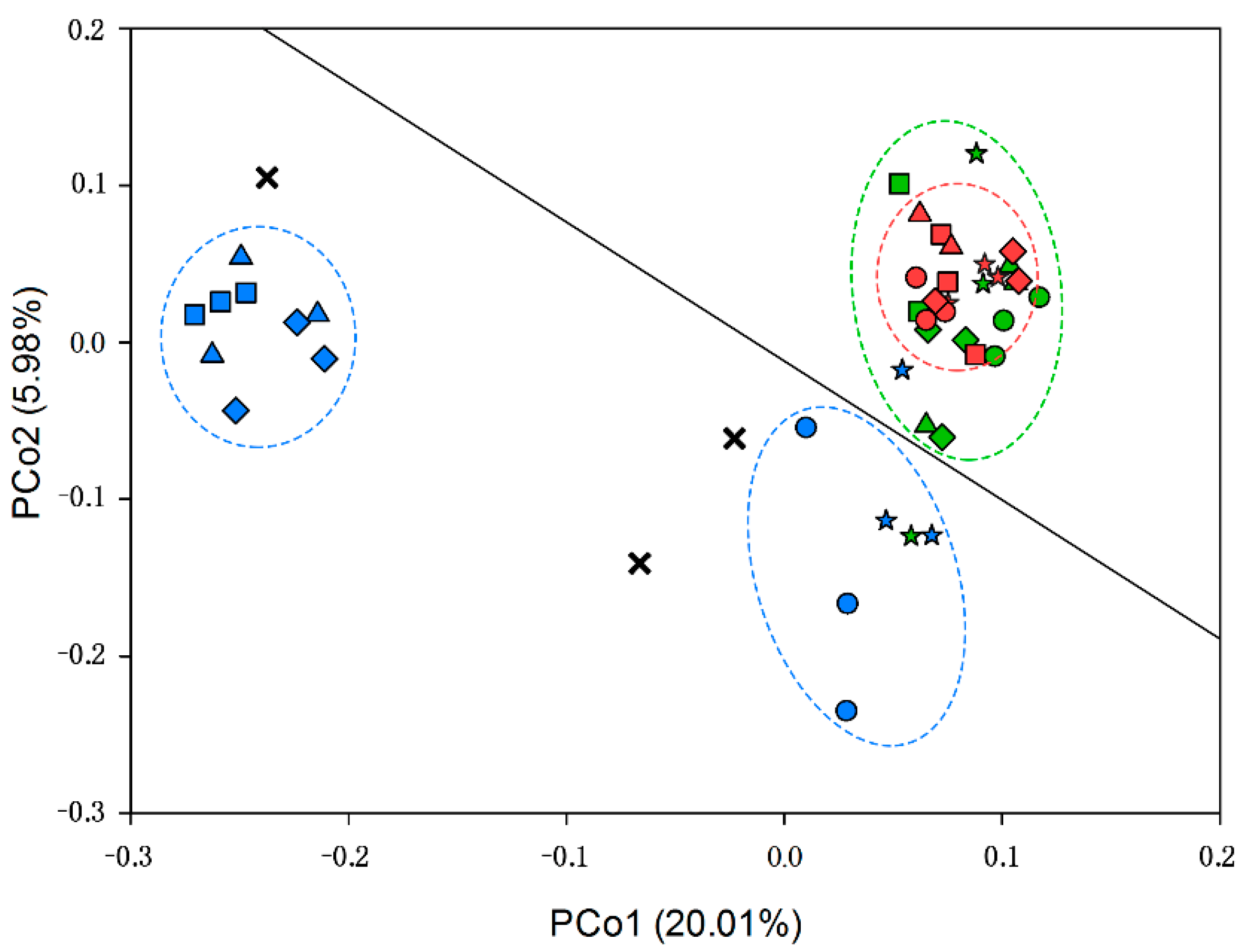
3.1.2. Beta Diversity of Soil Microbial Community
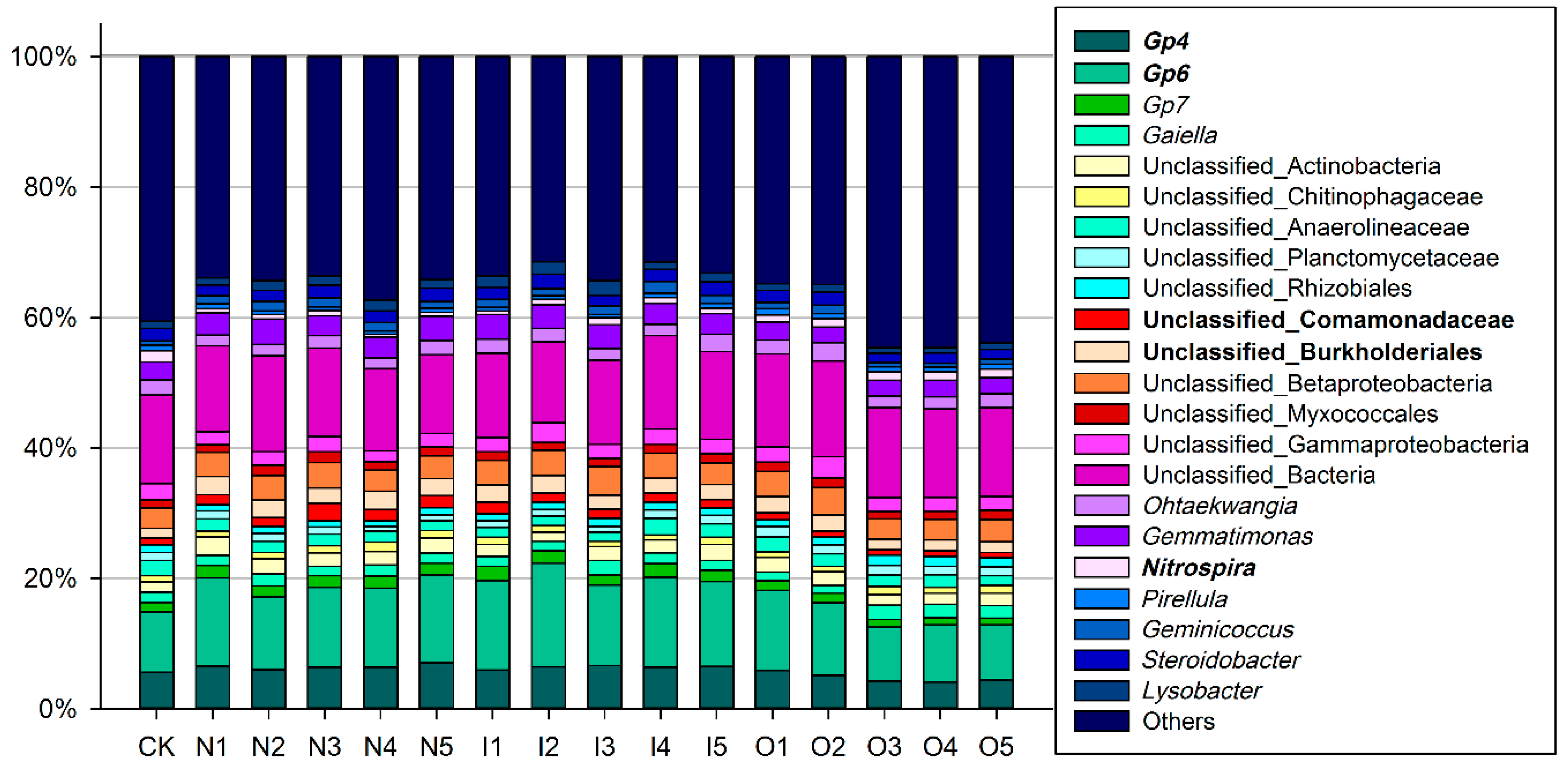
3.1.3. Microbial Community Structure
3.2. Soil Microbial Biomass Carbon and Nitrogen
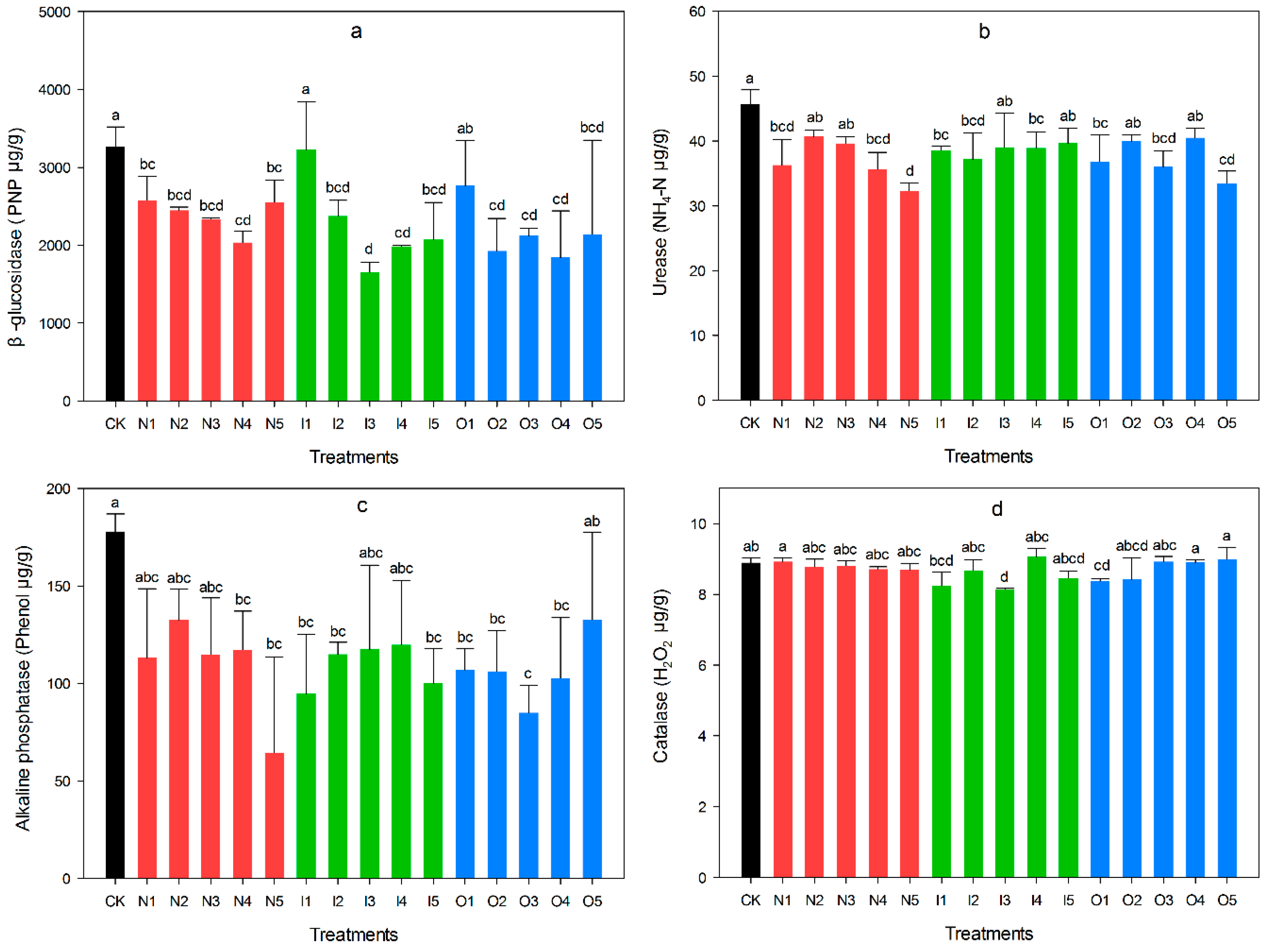
3.3. Soil Enzyme
4. Discussion
- (i).
- Fungal dominant with more organic matter, which occurred in N2, I1 and I4.
- (ii).
- Fungal dominant with less organic matter, which occurred in N1, N3, N4, O2 and O5.
- (iii).
- Bacterial dominant with more organic matter, which occurred in N5, I2, I3 and O1.
- (iv).
- Bacterial dominant with less organic matter, which occurred in I5, O3 and O4.
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cheng, F.; Wang, H.; Yun, W. Study on investigation and assessment of cultivated land quality grade in China. China Land Sci. 2014, 2, 75–82. (In Chinese) [Google Scholar]
- Zentner, R.P.; Campbell, C.A. First 18 years of a long-term crop rotation study in southwestern Saskatchewan-yields, grain protein, and economic performance. Can. J. Plant Sci. 1988, 68, 1–21. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics. Soil Till. Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Altieri, M.A. The ecological role of biodiversity in agroecosystems. Agric. Ecosyst. Environ. 1999, 74, 19–31. [Google Scholar] [CrossRef]
- Henle, K.; Alard, D.; Clitherow, J.; Cobb, P.; Firbank, L.; Kull, T.; Mccracken, D.; Moritz, R.F.A.; Niemelä, J.; Rebane, M.; et al. Identifying and managing the conflicts between agriculture and biodiversity conservation in Europe—A review. Agric. Ecosyst. Environ. 2008, 124, 60–71. [Google Scholar] [CrossRef]
- Xu, Z.; Xu, J.; Deng, X.; Huang, J.; Uchida, E.; Rozelle, S. Grain for green versus grain: Conflict between food security and conservation set-aside in China. World Dev. 2006, 34, 130–148. [Google Scholar] [CrossRef]
- Lam, H.-M.; Remais, J.; Fung, M.-C.; Xu, L.; Sun, S.S.-M. Food supply and food safety issues in China. Lancet 2013, 381, 2044–2053. [Google Scholar] [CrossRef]
- Cassman, K.G. Ecological intensification of cereal production systems: Yield potential, soil quality, and precision agriculture. Proc. Natl. Acad. Sci. USA 1999, 96, 5952–5959. [Google Scholar] [CrossRef] [PubMed]
- Doré, T.; Makowski, D.; Malézieux, E.; Munier-Jolain, N.; Tchamitchian, M.; Tittonell, P. Facing up to the paradigm of ecological intensification in agronomy: Revisiting methods, concepts and knowledge. Eur. J. Agron. 2011, 34, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trend Ecol. Evolut. 2013, 28, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Rusinamhodzi, L.; Corbeels, M.; Nyamangara, J.; Giller, K.E. Maize-grain legume intercropping is an attractive option for ecological intensification that reduces climatic risk for smallholder farmers in central Mozambique. Field Crop Res. 2012, 136, 12–22. [Google Scholar] [CrossRef]
- Zhou, L.X.; Ding, M. Soil microbial characteristics as bioindicators of soil health. Biodivers. Sci. 2007, 15, 162–171. [Google Scholar]
- Yusuf, A.; Abaidoo, R.; Iwuafor, E.; Olufajo, O.; Sanginga, N. Rotation effects of grain legumes and fallow on maize yield, microbial biomass and chemical properties of an Alfisol in the Nigerian savanna. Agric. Ecosyst. Environ. 2009, 129, 325–331. [Google Scholar] [CrossRef]
- Lin, X.G.; Feng, Y.Z.; Zhang, H.Y.; Chen, R.R.; Wang, J.H.; Zhang, J.B.; Chu, H.Y. Long-term balanced fertilization decreases arbuscular mycorrhizal fungal diversity in an arable soil in North China revealed by 454 pyrosequencing. Environ. Sci. Technol. 2012, 46, 5764–5771. [Google Scholar] [CrossRef] [PubMed]
- Nivelle, E.; Verzeaux, J.; Habbib, H.; Kuzyakov, Y.; Decocq, G.; Roger, D.; Lacoux, J.; Duclercq, J.; Spicher, F.; Nava-Saucedo, J.-E. Functional response of soil microbial communities to tillage, cover crops and nitrogen fertilization. Appl. Soil Ecol. 2016, 108, 147–155. [Google Scholar] [CrossRef]
- Jusselme, M.D.; Miambi, E.; Mora, P.; Diouf, M.; Rouland-Lefèvre, C. Increased lead availability and enzyme activities in root-adhering soil of Lantana camara during phytoextraction in the presence of earthworms. Sci. Total Environ. 2013, 445, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Brussaard, L.; de Ruiter, P.C.; Brown, G.G. Soil biodiversity for agricultural sustainability. Agric. Ecosyst. Environ. 2007, 121, 233–244. [Google Scholar] [CrossRef]
- Myers, R. Nitrogen Management of Upland Crops: From Cereals to Food Legumes to Sugarcane; CAB International: Wallingford, UK, 1988. [Google Scholar]
- Angenent, L.T.; Kelley, S.T.; Amand, A.S.; Pace, N.R.; Hernandez, M.T. Molecular identification of potential pathogens in water and air of a hospital therapy pool. Proc. Natl. Acad. Sci. USA 2005, 102, 4860–4865. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.; Brookes, P.; Jenkinson, D. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Joergensen, R.G. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEC value. Soil Biol. Biochem. 1996, 28, 25–31. [Google Scholar] [CrossRef]
- Dick, R.P.; Breakwell, D.P.; Turco, R.F.; Doran, J.W.; Jones, A.J. Soil enzyme activities and biodiversity measurements as integrative microbiological indicators. Soil Sci. Soc. Am. J. 1996, 49, 247–271. [Google Scholar]
- Hoffmann, G.; Teicher, K. Ein kolorimetrisches verfahren zur bestimmung der ureaseaktivität in Böden. J. Plant Nutr. Soil Sci. 1961, 95, 55–63. [Google Scholar] [CrossRef]
- Hoffmann, G. Eine photometrische methode zur bestimmung der phosphatase-aktivität in Böden. J. Plant Nutr. Soil Sci. 1968, 118, 161–172. [Google Scholar] [CrossRef]
- Torsvik, V.; Øvreås, L. Microbial diversity and function in soil: From genes to ecosystems. Curr. Opin. Microbiol. 2002, 5, 240–245. [Google Scholar] [CrossRef]
- Scheu, S. Changes in microbial nutrient status during secondary succession and its modification by earthworms. Oecologia 1990, 84, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Attard, E.; Poly, F.; Commeaux, C.; Laurent, F.; Terada, A.; Smets, B.F.; Recous, S.; Roux, X.L. Shifts between Nitrospira- and Nitrobacter-like nitrite oxidizers underlie the response of soil potential nitrite oxidation to changes in tillage practices. Environ. Microbiol. 2010, 12, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Sul, W.J.; Asuming-Brempong, S.; Wang, Q.; Tourlousse, D.M.; Penton, C.R.; Deng, Y.; Rodrigues, J.L.; Adiku, S.G.; Jones, J.W.; Zhou, J. Tropical agricultural land management influences on soil microbial communities through its effect on soil organic carbon. Soil Biol. Biochem. 2013, 65, 33–38. [Google Scholar] [CrossRef]
- Van, V.T.; Berge, O.; Ke, S.N.; Balandreau, J.; Heulin, T. Repeated beneficial effects of rice inoculation with a strain of Burkholderia vietnamiensison early and late yield components in low fertility sulphate acid soils of Vietnam. Plant Soil 2000, 218, 273–284. [Google Scholar] [CrossRef]
- Yoshida, M.; Ishii, S.; Otsuka, S.; Senoo, K. Temporal shifts in diversity and quantity of nirS and nirK in a rice paddy field soil. Soil Biol. Biochem. 2009, 41, 2044–2051. [Google Scholar] [CrossRef]
- Zaller, J.G.; Köpke, U. Effects of traditional and biodynamic farmyard manure amendment on yields, soil chemical, biochemical and biological properties in a long-term field experiment. Biol. Fert. Soils 2004, 40, 222–229. [Google Scholar] [CrossRef]
- Hu, S.; Chapin, F.S., III; Firestone, M.K.; Field, C.B.; Chiariello, N.R. Nitrogen limitation of microbial decomposition in a grassland under elevated CO2. Nature 2001, 409, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.A.; Biederbeck, V.O.; Zentner, R.P.; Lafond, G.P. Effect of crop rotations and cultural practices on soil organic matter, microbial biomass and respiration in a thin Black Chernozem. Can. J. Soil Sci. 1991, 71, 363–376. [Google Scholar] [CrossRef]
- Sparling, G.P. Ratio of microbial biomass carbon to soil organic carbon as a sensitive indicator of changes in soil organic matter. Soil Res. 1992, 30, 195–207. [Google Scholar] [CrossRef]
- Thiet, R.K.; Frey, S.D.; Six, J. Do growth yield efficiencies differ between soil microbial communities differing in fungal:bacterial ratios? Reality check and methodological issues. Soil Biol. Biochem. 2006, 38, 837–844. [Google Scholar] [CrossRef]
- De Graaff, M.-A.; Classen, A.T.; Castro, H.F.; Schadt, C.W. Labile soil carbon inputs mediate the soil microbial community composition and plant residue decomposition rates. New Phytol. 2010, 188, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Rousk, J.; Frey, S.D. Revisiting the hypothesis that fungal-to-bacterial dominance characterizes turnover of soil organic matter and nutrients. Ecol. Monogr. 2015, 85, 457–472. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Lea, T.; Beaudoin, J.L.; Clayton, G.W. Soil microbial biomass, functional diversity and crop yields following application of cattle manure, hog manure and inorganic fertilizers. Can. J. Soil Sci. 2005, 85, 193–201. [Google Scholar] [CrossRef]
- Zhen, Z.; Liu, H.; Wang, N.; Guo, L.; Meng, J.; Ding, N.; Wu, G.; Jiang, G. Effects of manure compost application on soil microbial community diversity and soil microenvironments in a temperate cropland in China. PLoS ONE 2014, 9, e108555. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, B.S.; Johnson, D.E. Influence of tillage systems on weed seedling emergence pattern in rainfed rice. Soil Till. Res. 2009, 106, 15–21. [Google Scholar] [CrossRef]
- Inselsbacher, E.; Hinko-Najera Umana, N.; Stange, F.C.; Gorfer, M.; Schüller, E.; Ripka, K.; Zechmeister-Boltenstern, S.; Hood-Novotny, R.; Strauss, J.; Wanek, W. Short-term competition between crop plants and soil microbes for inorganic N fertilizer. Soil Biol. Biochem. 2010, 42, 360–372. [Google Scholar] [CrossRef]
- Kotroczó, Z.; Veres, Z.; Fekete, I.; Krakomperger, Z.; Tóth, J.A.; Lajtha, K.; Tóthmérész, B. Soil enzyme activity in response to long-term organic matter manipulation. Soil Biol. Biochem. 2014, 70, 237–243. [Google Scholar] [CrossRef]
- Guenzi, W.; McCalla, T. Phenolic acids in oats, wheat, sorghum, and corn residues and their phytotoxicity. Agron. J. 1966, 58, 303–304. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Renella, G.; Puglisi, E.; Ceccanti, B.; Masciandaro, G.; Fornasier, F.; Moscatelli, M.C.; Marinari, S. Soil enzymology: Classical and molecular approaches. Biol. Fert. Soils 2012, 48, 743–762. [Google Scholar] [CrossRef]
- Kiss, S.; Simihaian, M. Improving Efficiency of Urea Fertilizers by Inhibition of Soil Urease Activity; Springer: Berlin, Germany, 2013. [Google Scholar]
- Hu, J.; Yang, A.; Wang, J.; Zhu, A.; Dai, J.; Wong, M.H.; Lin, X. Arbuscular mycorrhizal fungal species composition, propagule density, and soil alkaline phosphatase activity in response to continuous and alternate no-tillage in Northern China. Catena 2015, 133, 215–220. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation | Management Groups | |||
|---|---|---|---|---|
| No Management (N) | Irrigation (I) | Organic Manure and Irrigation (O) | Control Group (CK) | |
| Native plants (1) | N1 | I1 | O1 | n |
| Soybean-maize (2) | N2 | I2 | O2 | n |
| Soybean-dahurian wild rye (3) | N3 | I3 | O3 | n |
| Alfalfa-maize (4) | N4 | I4 | O4 | n |
| Alfalfa-dahurian wild rye (5) | N5 | I5 | O5 | n |
| Control group (CK) | n | n | n | CK |
| Sample ID | Shannon Index | Observed_Otus | Chao1 Index | PD_Whole_Tree |
|---|---|---|---|---|
| CK | 9.88 ± 0.07abc | 1986.00 ± 25.96abc | 3309.87 ± 40.61abc | 111.25 ± 2.11abc |
| N1 | 9.73 ± 0.03bcd | 1887.33 ± 9.03cdef | 3201.78 ± 132.00abcd | 105.46 ± 0.80bcd |
| N2 | 9.85 ± 0.08abc | 1941.33 ± 53.32bcd | 3205.43 ± 158.09abcd | 106.96 ± 2.84bcd |
| N3 | 9.73 ± 0.09bcd | 1929.67 ± 41.49bcd | 3200.35 ± 113.72abcd | 106.74 ± 3.00bcd |
| N4 | 9.76 ± 0.04abcd | 1858.67 ± 38.42def | 3051.08 ± 78.06cde | 101.33 ± 2.12df |
| N5 | 9.53 ± 0.15d | 1773.50 ± 97.50fg | 2871.64 ± 141.33e | 97.50 ± 5.01f |
| I1 | 9.61 ± 0.07cd | 1801.00 ± 42.62efg | 2986.73 ± 118.40de | 101.16 ± 3.10df |
| I2 | 9.55 ± 0.05d | 1803.67 ± 46.95efg | 3056.10 ± 160.58cde | 101.38 ± 3.14df |
| I3 | 9.23 ± 0.43e | 1714.50 ± 139.50g | 2866.69 ± 315.44e | 96.85 ± 6.06f |
| I4 | 9.73 ± 0.10bcd | 1896.67 ± 60.81bcde | 3110.23 ± 131.15bcde | 104.83 ± 3.54cd |
| I5 | 9.73 ± 0.07bcd | 1899.00 ± 22.55bcde | 3149.99 ± 101.43abcde | 104.73 ± 2.43cd |
| O1 | 9.87 ± 0.04abc | 2016.33 ± 23.70ab | 3271.84 ± 59.80abcd | 112.05 ± 1.11ab |
| O2 | 9.89 ± 0.04ab | 2001.33 ± 43.76abc | 3330.37 ± 129.47abc | 111.52 ± 2.65ab |
| O3 | 10.02 ± 0.03a | 2088.67 ± 20.76a | 3463.42 ± 13.08a | 116.00 ± 0.83a |
| O4 | 9.98 ± 0.03ab | 2062.67 ± 17.46a | 3412.02 ± 53.47ab | 114.04 ± 1.03a |
| O5 | 9.10 ± 0.02ab | 2073.67 ± 27.40a | 3440.88 ± 110.25a | 115.26 ± 0.60a |
| Sample ID | SOC | Soil TN | C-to-N Ratio of Soil | MBC | MBN | MBC-to-MBN Ratio | % of Total C or N as | |
|---|---|---|---|---|---|---|---|---|
| (g/kg) | (mg/kg) | MBC | MBN | |||||
| CK | 5.74 | 0.80b | 7.17abc | 256.48abc | 20.51ab | 12.56 | 4.47 | 3.61 |
| N1 | 5.50 | 0.90b | 6.14abc | 293.86ab | 20.88ab | 14.34 | 5.34 | 3.67 |
| N2 | 5.89 | 1.36a | 4.64c | 249.82abc | 18.60ab | 13.67 | 4.24 | 2.80 |
| N3 | 5.28 | 0.95b | 5.53bc | 256.30abc | 17.60b | 14.55 | 4.85 | 2.50 |
| N4 | 5.32 | 1.00b | 5.43bc | 241.32abc | 16.43b | 14.86 | 4.54 | 2.09 |
| N5 | 5.23 | 0.86b | 6.08abc | 226.64bc | 21.19ab | 11.21 | 4.33 | 3.30 |
| I1 | 6.01 | 0.78b | 7.74ab | 219.12c | 21.02ab | 12.66 | 3.65 | 3.10 |
| I2 | 6.36 | 0.96b | 6.63abc | 268.66abc | 22.37ab | 12.27 | 4.22 | 2.98 |
| I3 | 5.92 | 0.92b | 6.43abc | 225.88bc | 20.22ab | 11.37 | 3.82 | 3.05 |
| I4 | 6.02 | 0.86b | 7.01abc | 268.93abc | 19.10ab | 14.33 | 4.47 | 3.09 |
| I5 | 5.29 | 0.71b | 7.61ab | 255.33abc | 20.84ab | 12.30 | 4.83 | 3.67 |
| O1 | 6.22 | 0.73b | 8.47a | 235.93abc | 21.26ab | 11.13 | 3.79 | 3.73 |
| O2 | 5.19 | 0.71b | 7.37ab | 255.92abc | 20.18ab | 12.84 | 4.93 | 3.87 |
| O3 | 5.37 | 0.81b | 6.79abc | 253.21abc | 22.23ab | 12.21 | 4.72 | 3.33 |
| O4 | 5.66 | 0.80b | 7.00abc | 278.92abc | 24.75a | 11.38 | 4.93 | 4.28 |
| O5 | 5.63 | 0.90b | 6.32abc | 303.56a | 21.53ab | 14.43 | 5.39 | 3.36 |
| Soil Properties | pH | AN | AK | MBC | SOC | TN | AP | APL | β | Ur | CAT |
|---|---|---|---|---|---|---|---|---|---|---|---|
| pH | 1.000 | ||||||||||
| AN | −0.302 * | 1.000 | |||||||||
| AK | −0.178 | 0.243 | 1.000 | ||||||||
| MBC | 0.034 | 0.057 | 0.000 | 1.000 | |||||||
| SOC | −0.215 | 0.386 ** | 0.396 ** | −0.091 | 1.000 | ||||||
| TN | −0.155 | 0.620 ** | 0.321 * | 0.049 | 0.358 * | 1.000 | |||||
| AP | −0.333 * | 0.285 * | −0.051 | −0.044 | 0.136 | −0.128 | 1.000 | ||||
| APL § | −0.256 | 0.535 ** | 0.237 | 0.242 | 0.343* | 0.303 * | 0.263 | 1.000 | |||
| β§ | −0.131 | 0.179 | 0.013 | −0.071 | 0.023 | −0.047 | 0.451 ** | 0.183 | 1.000 | ||
| Ur § | −0.330 * | 0.372 ** | 0.200 | 0.055 | 0.221 | 0.185 | 0.548 ** | 0.422 ** | 0.116 | 1.000 | |
| CAT § | −0.071 | 0.172 | −0.042 | 0.775 ** | −0.111 | 0.172 | −0.069 | 0.226 | 0.029 | −0.032 | 1.000 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.; Wu, C. Effects of Short-Term Set-Aside Management Practices on Soil Microorganism and Enzyme Activity in China. Int. J. Environ. Res. Public Health 2017, 14, 913. https://doi.org/10.3390/ijerph14080913
Li G, Wu C. Effects of Short-Term Set-Aside Management Practices on Soil Microorganism and Enzyme Activity in China. International Journal of Environmental Research and Public Health. 2017; 14(8):913. https://doi.org/10.3390/ijerph14080913
Chicago/Turabian StyleLi, Guangyu, and Cifang Wu. 2017. "Effects of Short-Term Set-Aside Management Practices on Soil Microorganism and Enzyme Activity in China" International Journal of Environmental Research and Public Health 14, no. 8: 913. https://doi.org/10.3390/ijerph14080913
APA StyleLi, G., & Wu, C. (2017). Effects of Short-Term Set-Aside Management Practices on Soil Microorganism and Enzyme Activity in China. International Journal of Environmental Research and Public Health, 14(8), 913. https://doi.org/10.3390/ijerph14080913